
Abstract
Traumatic brain injury constitutes a significant proportion of cases requiring forensic examination, and it encompasses (1) blunt, nonmissile head injury, especially involving motor vehicle accidents, and (2) penetrating, missile injury produced by a range of high- and lower-velocity projectiles. This review examines the complex pathophysiology and biomechanics of both types of neurotrauma and assesses the macroscopic and histologic features of component lesions, which may be used to determine the cause and manner of death resulting from an intentional assault or accident. Estimation of the survival time postinjury by pathologic examination is also important where malicious head injury is suspected, in an attempt to ascertain a time at which the traumatic event might have been committed, thereby evaluating the authenticity of statements made by the alleged perpetrator.
Keywords
In human medicine, forensic pathology has evolved into a distinct subspecialty. While veterinary pathologists have traditionally been infrequently called on to adjudicate in forensic cases where intentional head trauma is suspected, such cases are becoming more common and require more in-depth forensic knowledge. When an animal presents with signs of external head trauma, such as scalp bruises and skin abrasions, abetted by a reliable history of a traumatic event, a diagnosis of traumatic brain injury (TBI) may be relatively straightforward. However, in the absence of visible evidence of trauma and a reliable history, the onset of neurologic signs may provide the only clue that a traumatic event has occurred, especially since grazing livestock and free-roaming companion animals are not closely scrutinized.
In forensic pathology, the timing of tissue responses is often the primary medicolegal task, particularly as it may support or refute statements made by a suspected perpetrator of abuse. These pathologic determinations usually depend on an estimate of the time between a traumatic insult and death. This interval between a traumatic event and circulatory arrest is defined as the survival time and, as such, equates to duration of the brain circulation after injury, although the latter can vary among the brain regions. However, while an estimate of the survival time following head trauma is sometimes possible by the application of relevant histologic and immunohistochemical techniques, the ability to date/age an injury solely on this basis should not be overestimated. 66,75,103
It is not always possible to accurately date TBI lesions because (1) determinations are variously based on studies in humans, experimental animal models (usually laboratory rodents), or both and (2) it is not always stated from which species the data were derived and how these timings were calculated. 46 These assessments are important since pathologic processes often differ in their time of development among species. Secondary axotomy, for example, following traumatic axonal injury (AI) requires a survival period of >12 hours in humans but only 2 to 4 hours in rats and cats. 24 These interspecies differences are highlighted by the frequent failure of potential neuroprotective agents for TBI to provide a consistent and significant improvement of outcome in human clinical trials, despite showing promise in experimental animals. If more cases of TBI in domestic animals that result in fatality or euthanasia are subjected to rigorous neuropathologic examination, the timeline of development of the resulting brain lesions in different species could be more accurately evaluated. 39 Moreover, correlation of histology with magnetic resonance imaging may facilitate an assessment of the temporal progression of TBI lesions. 60
While the necropsy for forensic purposes aims to determine the cause and manner of death, it may not be possible to determine the precise mode of death in some circumstances, and the unsatisfying label of “undetermined” may have to be assigned to a case. In view of the frequent difficulty in interpreting the time frame of histopathologic findings after TBI, there may be insufficient data to determine the survival time in a given case, despite a thorough pathologic examination. It may be possible to only conclude that a given finding is consistent with (or not consistent with) a particular insult, or a certain scenario may be favored on the basis that there is insufficient reason to consider an alternative possibility.
In the present forensic context of TBI, it is often pertinent to assess whether there has been a head injury, the approximate timing of its occurrence, the likely mechanisms of injury, the nature and distribution of lesions, and whether there were any preexisting (premorbid) lesions that might be confounding variables. In humans, the presence of natural disease processes in the nervous system of adults past middle age is very high, but this is less true of domestic animal species.
A number of factors can influence the interpretation of neuropathologic findings after TBI—including (1) the death-to-necropsy interval, with autolytic and sometimes putrefactive changes contributing to the final histologic expression; (2) the histologic technique used to age a given lesion—with traumatic AI, for example, being detected much earlier with amyloid precursor protein (APP) immunohistochemistry than with routine hematoxylin and eosin (HE) or silver staining; (3) the volume of brain parenchyma involved, with focal and diffuse lesions differing in the spectrum and rapidity of cellular reactions; (4) the age of the patient, with mature and immature brains having different reaction times for different pathologic processes; (5) the many artifacts encountered in postmortem brains, which must be distinguished from antemortem lesions; and (6) the complex interplay of immediate and evolving events constituting TBI, often rendering it very difficult to reconstruct the numerous primary and secondary lesions of varying severity and regional distribution that form the final expression of TBI.
Neuropathologic Techniques
As a prelude to histopathologic examinations, good fixation of the brain is essential. The formalin used should be neutral and buffered, at least 10% (3.7% formaldehyde) concentration, and 10 times the volume of brain suspended in it. For larger brains, a fixation time of at least 2 weeks is recommended with immersion fixation but not exceeding 4 weeks (when formalin becomes acidic), and the formalin should be changed after 24 hours and toward the end of the first week. For large bovine and human brains, fixation in concentrated (40%) formalin can be effective. 66,103
With immersion fixation of the brain, there is unavoidable mechanical damage during removal and the development of artifacts, but any intravascular pathology is not disturbed. By contrast, perfusion of fixative into the brain via carotid artery stumps fixes neural tissue in situ in about 2 to 3 hours and minimizes mechanical handling artifacts. After perfusion, the brain should remain in situ for at least 2 hours, up to 24 hours with large brains, to permit adequate penetration of the fixative. However, perfusion fixation is often impractical, and the lengthy death-to-necropsy interval frequently encountered in forensic cases means that perfusion will be incomplete and inconsistent. Artifactual brain shrinkage may be caused by unduly high osmolality (>0.1 mol/l) of the perfusate; conversely, immersion fixation with formalin may increase the brain weight by up to 10%, although this will usually almost return to its fresh state weight after 3 weeks of fixation. The entire brain should be sectioned in a transverse plane at about 0.5-cm intervals, sometimes aided by a frame composed of 2 L-shaped metal bars, and during processing, the high lipid and water content of brain requires longer dehydration times. 66,70,103
Immunohistochemistry has now replaced many traditional special neuropathologic stains, especially in human laboratories but also, increasingly, in veterinary settings. However, to be reliable, cognizance must be taken of appropriate fixation times and antigen retrieval methods, correct antibody dilutions, and positive and negative controls to ensure that the antibody used is detecting the antigen of interest in a given species. Antibodies for use in formalin-fixed, paraffin-embedded material are now widely available. 26
Postmortem decomposition of the brain is the sum of 3 processes. Autolysis is cellular disintegration resulting from enzymatic autodigestion. In humans, there is a tendency for normal brains to autolyze more rapidly than those with chronic disease. Putrefaction refers to anaerobic degradation of tissue by the action of invading intestinal bacteria, while decay is aerobic destruction of tissue by other microorganisms, sometimes abetted by arthropods, rodents, and other necrophages. 84 In humans, brains have been macroscopically classified as normal, discolored, softened, and pulpified with postmortem intervals of 1–2, 2–9, 4–7, and 3–30 days, respectively, 96 but unless these timings can be reliably corroborated, the descriptions are approximate, as rates of decomposition vary widely under different circumstances.
Postmortem Artifacts
Fresh brain is soft—even more so when diseased and thus easily damaged. It must therefore be handled with care. Since most brains examined in diagnostic laboratories are fixed by immersion in 10% neutral buffered formalin, artifacts will commonly be present, especially if the postmortem is delayed. Artifactual vacuolation of neurons, glia, and neuropil, for example, develops so commonly between death and necropsy that the diagnosis of vacuolation attending early ischemic-hypoxic changes and edema can be problematic. If there is a difference in the number and size of clear vacuoles at the lesion margin and if this vacuolation diminishes with increasing distance from the lesion, edema can sometimes be more confidently diagnosed. Artifactual perivascular and perineuronal spaces, due to swelling of astrocytic processes, may also mimic edema in immersion-fixed material. 79,102 Axonal spheroids are commonly found in medullary cuneate and gracile nuclei in aged animals of all species, and these must be distinguished from neuroaxonal dystrophy. 89
Shrunken, hyperchromatic “dark” neurons are commonly found at necropsy, with their number decreasing in humans as the postmortem interval increases and minimized by a death-to-necropsy duration of at least 10 hours before formalin fixation. They are infrequently found in perfusion-fixed brains in which the adverse effects of mechanical handling during brain removal are limited. Dark neurons have a “corkscrew” dendrite, an important discriminator of this change from similar-appearing neurons in some diseases states. Dark neurons represent early, potentially reversible alterations with preservation of cellular substructure, and serial studies have revealed temporal reversal of cytoplasmic condensation. 20 The most common site of selective postmortem autolysis is the cerebellar granular layer (sometimes termed cerebellar conglutination), especially in cattle 129 and humans. 6,65 Individual or small groups of granule cells shrink, and chromatin condenses (pyknosis); then, karyorrhexis ensues, with the granular layer eventually becoming severely depleted. Purkinje cells and Bergmann glia are initially preserved, but when the granular layer is represented by only a few degenerate neurons and remnant nuclear dust, Purkinje cells are “ghosts” with cytoplasmic homogenization and indistinct nuclei. This artifact increases in frequency as the interval between death and necropsy rises, and it must also be distinguished from several degenerative, toxic, and metabolic disorders that selectively damage this neuronal population. In normal sheep and cattle, incidental neuronal vacuolation is sometimes found in certain brainstem nuclei, which must be distinguished from spongiform encephalopathy. 89
Classification of TBI
TBI can be broadly classified in 2 forms: blunt, nonmissile (closed) or penetrating, missile (open; Table 1). 13,15,66,103
Pathologic Features of Traumatic Brain Injury.

TBI has received relatively little attention in the veterinary literature, 39,83,118 and much of our understanding of TBI in animals is derived from neuropathologic studies of human neurotrauma cases. While most domestic animal species have a relatively large gyrencephalic brain resembling that of humans and although the general pattern of reaction of their brains to a traumatic insult appears to be similar, extrapolating human TBI findings to the veterinary sphere can sometimes be problematic. There are significant anatomic differences in brain and encasing skull between humans and domestic animals, and the orientation of the neuraxis is very different. Nevertheless, animal models have been invaluable in elucidating the pathology and pathogenesis of human head injury, as many replicate the important neuropathologic lesions produced by TBI in humans. 41
Accidental, nonmissile head injury is commonly encountered in veterinary practice as a result of animals being struck by motor vehicles or involved in falls, bites from predation, kicks, or crushing injuries. Dogs are most frequently struck by automobiles; cats may sustain head injuries after falling from buildings and being hit by motor vehicles; and horses and ruminants can suffer cranial injuries during periods of excitement, by falling when rearing, and with attempted restraint. 123 In 1 study, 124 most dogs sustained head injuries from motor vehicle accidents, while the majority of head-injured cats had crush injuries. Although there is a paucity of data concerning blunt trauma in dogs, 2 large epidemiologic studies were conducted in the United States. In the first, 122 vehicular trauma accounted for 90% of cases. The thorax was the most commonly traumatized region (70%), followed by the abdomen and extremities; 25% of dogs sustained head trauma. In the second study, 73 the skeletal system was the favored site for traumatic injury, but central nervous system injuries occurred in 50% of cases. Penetrating, missile head injury is commonly used for the humane euthanasia of livestock by firearms to produce immediate unconsciousness, either by gunshot wound to the head or by a penetrating captive bolt pistol (humane stunner). 36,40 However, firearms and assaults with a variety of weapons may cause inadvertent or intentional head injuries to animals.
This review discusses the principal diagnostic brain lesions constituting nonmissile and missile head injury and their pathogenesis. Additionally, where possible, an indication of the dating/aging of the tissue reactions constituting a given lesion is provided, which can have important medicolegal implications.
Biomechanical Factors Determining Traumatic Cerebral Lesions
The necropsy for TBI should incorporate an assessment of injury biomechanics because these mechanisms are often the principal focus of a forensic examination. The actual or candidate impact surface or object suspected of causing an injury should be examined for linkage to the pattern of scalp injuries. It is important to elicit the location and dimensions of the impact site, as well as whether the head was moving at the time of impact or was motionless relative to the impacting force and whether the scalp or skull was intact. These observations may permit an estimation of the mass, velocity, and direction of the impacting object. However, because many external variables can affect the type and extent of posttraumatic brain lesions, there is often difficulty interpreting them. 9,76
Head injuries are sometimes conceived of as occurring only at impact and being essentially completed within moments, and the deleterious effects of a localized impact are assumed to remain localized. Indeed, acceleration forces may cause localized injury to the skull, brain, or meninges. However, as a general principle, when the head is subjected to acceleration forces, there is compelling evidence that the entire brain usually sustains injurious forces, at least to some degree, irrespective of the apparently trivial nature of this injury. 9,76
Biomechanics is the study of tissue failure (manifest as injury) when exposed to a given loading event, after which it ceases, either structurally or functionally, to perform normally. In this case, loading refers to forces acting on the skull/brain in the acute phase of an injury. Neural tissue is sensitive not only to the load applied (force, pressure, stress [force applied per unit area] or strain [deformation from the original configuration]) but also to the rate and duration of the loading. Determination of injury tolerance is also important in biomechanics and is defined as the level of a particular physical parameter (eg, force, acceleration/deceleration, stress, strain, impact velocity) above which tissue failure (injury) will occur. 9,76
External loading to the head may affect the intracranial contents by directly displacing or deforming the skull or intracranial partitions (contact phenomena) or by differential motion between skull/dura and the intracranial contents by virtue of these contents lagging behind the motion of the skull during acceleration of the head (inertial or impulsive phenomena). With the exception of crushing injuries, mechanical forces that lead to TBI are applied dynamically. 9,76
The mechanisms of brain damage differ in many respects between closed, nonmissile and missile head injury. In the former, acceleration/deceleration forces impart large-momentum, rotational, and shear forces to the head and brain but relatively low kinetic energy (KE). By contrast, penetrating head wounds produced by firearms deliver high focal KE and relatively low cranial momentum. 112
Most head injuries are produced by contact or acceleration/deceleration forces. Contact injuries such as scalp lacerations, skull fractures (with or without an associated epidural hematoma), contusions, and lacerations occur when the head strikes or is struck by an object. However, the absence of evidence of a contact injury, such as scalp bruising or skull fracture, does not necessarily imply that there was no head impact. 9,27,52,111
Impact to a head that is free to move (and change in velocity) produces loss of consciousness more often than one that is constrained, with the latter being more likely to cause localized defects of the skull and brain. When the head is freely mobile, direct impact produces significant acceleration/deceleration forces, with rotational movement in the transverse plane more productive of diffuse AI and coma than translational (linear) movements. If the line of action of an impact force passes through the center of gravity of the head, it will be accelerated without rotation; if not, the head will sustain both linear and angular acceleration. Most motion in real-world TBI is a combination of linear and angular (curvilinear) motion. Primary axonal and vascular damage following a head impact is more well correlated with the amount of brain deformation (strain) rather than stress caused by rotational acceleration/deceleration of the head, with the cerebral hemispheres seemingly more vulnerable than the brainstem. 9,27,52,111
Shearing forces and inertial loading are related to brain mass; therefore, animals that have small brains can tolerate much greater acceleration/deceleration forces than nonhuman primates and humans can, and the margin between fatal and nonfatal impacts to the head is slender. Moreover, in quadrupeds, the long axes of the brain and spinal cord are parallel, while in humans they are almost at right angles. This almost linear neuraxis in quadrupeds may impede rotational shearing after TBI and render the subject much less vulnerable to concussion. 35 After some head impacts, the brain of many animals is better protected than that of humans by well-developed temporal muscles and large frontal sinuses. 123
Blunt, Nonmissile Head Injury
TBI of this type is composed of many different neural lesions that may occur singly or, more commonly, in various combinations, and each patient with TBI has a unique and complex pattern of brain damage. Brain damage after TBI is heterogeneous, and the injury severity increases with the multiplicity of lesions. 1 –3
The pathologic substrate for ongoing neurologic deficits is the sum of the different types of direct and immediate (primary) and evolving (secondary) brain damage. This interplay between primary and secondary injury explains why an animal presenting initially with a mild brain injury may subsequently develop severe brain damage and, conversely, why one with a seemingly life-threatening condition substantially recovers. 2,13,15
The traumatic brain lesion develops through several phases, each with its own pathophysiology and outcome, which may or may not lead to the next phase, depending on the severity of the injury and the efficiency of repair. Pathologic processes in TBI occur not as separate events but as a cascading phenomenon, which is dynamic, unstereotypic, and inherently unpredictable. 15,75
Thus, while the classification of TBI appears well defined in concept, in reality it is extremely variable among individual cases. The types of injuries are limited in number but are expressed in a complex manner in each case. If primary injuries do not cause immediate death, longer survival permits the development of secondary changes, which may render detection of primary lesions difficult. 15,75
Blunt, nonmissile TBI may be categorized as primary or secondary, with parenchymal damage occurring in a focal, multifocal, or diffuse pattern (Table 1). From a clinical perspective, recognition of these types of brain damage is important. Primary brain damage occurs at the time of head impact; it has an almost immediate clinical effect; and it is refractory to most treatment. By contrast, secondary brain damage occurs in a dynamic and evolving manner at varying times postimpact and may be amenable to treatment, although it can be manifest very rapidly after a head impact. If a head-injured animal survives the initial traumatic insult, any increase in a neurologic deficit or deeper level of unconsciousness will be determined by the severity of secondary injury events. Accordingly, an important part of the clinical management of TBI is the differentiation between primary and secondary damage. Focal and diffuse brain damage also have individual pathophysiology, and while it is usually possible to ascertain the nature of a focal lesion, this is often more difficult when diffuse brain damage is present, particularly in the absence of an intracranial mass lesion.*
Primary Brain Damage
Primary injury is produced by mechanical forces causing brain deformation at the moment of head impact, with neural elements (blood vessels, axons, neurons, and glia) directly damaged in a focal, multifocal, or diffuse pattern. Moreover, injury thresholds for the various neural elements differ. 13,15
The scalp is the first barrier after head impact and provides protection to underlying structures by absorbing and dampening some of the effects of trauma. The scalp is composed of 5 layers: skin, dense connective tissue, galea aponeurotica, loose connective tissue, and the periosteum of the skull. The scalp is also highly vascular and will bleed profusely after laceration. The curved structure of the cranium receives an external impact at a point of contact and distributes that load across the bone to its margins, where the load is then shared with and transmitted to other calvarial bones via the sutures. The potential for cranial fracture depends on the location of the applied load, its magnitude, and the nature of the surface applying the load to the head. Cranial fractures can be linear or depressed or—in the case of impacts from small objects such as a hammer—punched out, small, and circular, with the underlying brain damage depending on the size of the blow. A single linear fracture confirms that the cranium has been subjected to trauma. A bilateral cranial fracture is suggestive, in humans at least, of inflicted trauma. While a calvarial fracture may not necessarily produce important neurologic dysfunction, sequelae of hemorrhage and contusions may; conversely, lethal TBI can occur in the absence of a cranial fracture. Inbending of the inner table of the calvarium may also lacerate the dura and cortex. 66,103,123
In horses, the most common and potentially harmful trauma to the head occurs when an animal lurches backward and strikes the poll, generally when being halter trained or forcibly restrained or led when excitable. There is rapid acceleration when the head arcs upward, then downward, and sudden deceleration when the head strikes a surface with considerable force. The resulting brain damage is greater when the head contacts a flat hard surface as compared with a wall or an overhead object. While there is seldom a full-thickness fracture of the occiput, the forces generated upon impact may be transmitted through the skull and fracture less robust bones on the sides and base of the calvarium. Fracture of basilar bones may cause meningeal hemorrhage around the brainstem and stretching/tearing of cranial nerves and, if depressed, laceration of these structures. The second-most frequent traumatic event is impact to the frontal or parietal region when a horse is kicked or collides with a fixed object. Blows to these skull bones are most likely to cause brain damage when contact between object and skull is narrow or uneven (eg, after a kick or collision with a post); however, when contact is more broadly distributed (eg, when a horse pitches forward into the ground at high speed), the force tends to be transmitted through the occipital condyles to the cervical vertebrae. 83
Contusions are common features of TBI and confirm that a head injury has occurred. However, contusions may be minimal, even absent, with fatal, diffuse TBI. Contusions are focal surface bruises (Figs. 1–3) resulting from damage to small blood vessels, and they present as petechial hemorrhages at the crests of gyri; they often increase in size over subsequent hours to days. Contusions may be subdivided into (1) contusion hemorrhages, when vascular injury predominates, and (2) contusion necrosis, with or without hemorrhage, when parenchymal injury is the principal feature; most contusions occur along a spectrum between these 2 extremes. Contusions are wedge shaped on cross section, with a broad base at the brain surface; at the deep aspect, they are directed toward and enter the subcortical white matter to varying depths. By contrast, infarcts are more randomly distributed and are not confined to gyral apices; furthermore, there is relative preservation of superficial cortical layers. Contusions generally open to the subarachnoid space and are associated with subarachnoid hemorrhage (SAH). Damage to blood vessels in contusions can lead to a cascade of events, including hemorrhage, thrombosis, and infarction. If there is coexistence of a contusion, hematoma formation at its margin, and adjacent meningeal hemorrhage, the term burst lobe is used (Fig. 1). When there is physical disruption of the neural parenchyma, the term laceration (Fig. 3) is applied. 4,15,80 –82,89
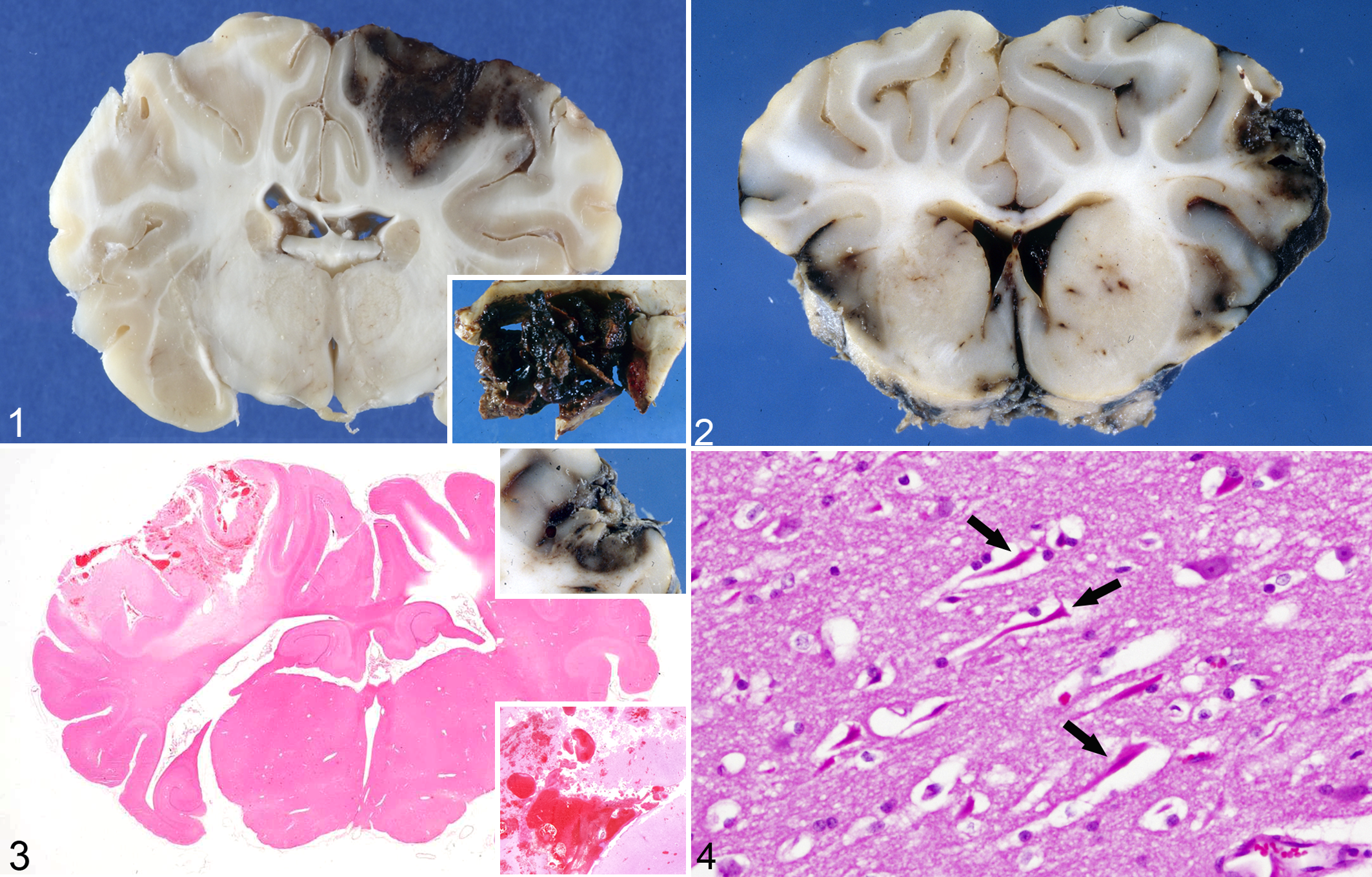
Traumatic impact contusions, brain, sheep.
Contusions often occur beneath the impact site (coup contusions), and they are produced by compressive forces beneath a site of skull inbending or tensile forces generated when the displaced skull suddenly resumes its original position. Coup contusions are generally small under the impact site but can be larger beneath depressed skull fractures. Contrecoup (contralateral) contusions may be found more or less opposite the impact site but do not necessarily indicate the direction of the applied force. 97 Intermediate coup contusions are sometimes found midway between coup and contrecoup contusions and may reflect tissue tears produced by severe diffuse AI. 80
Macroscopically, contusions are initially red, change to purple-black after several days, then dark brown after 3 to 4 weeks, and golden orange-brown after 6 weeks. With more chronic contusions, the subjacent white matter may be grayish due to myelin loss and gliosis, and the final outcome of contusion injury is a golden-brown, shrunken scar at the gyral crest. 80
The microscopic appearance of contusions depends on the survival time and factors such as acidosis, hypoxia, hypotension, and electrolyte disturbances. Small hemorrhages may be resorbed in a few days, while larger extravasations may require weeks to months. The larger the hemorrhagic component, the greater the cellular response. 110 Dating/aging of contusions has produced variable results, and the following guidelines are approximate. With respect to early reactions, edema is detectable within minutes of injury, increases over the following several hours, remains stable for a few days, then declines and disappears by about 6 days postinjury. Hemorrhage begins almost immediately in perivascular sites and extends into the adjacent parenchyma over the next few hours, being maximal by about 24 hours. While intact erythrocytes may remain for as long as 5 to 6 months in some cases, they generally deteriorate and disappear after about 5 days. Within a few hours of injury, neutrophils may be seen emigrating from vessels and infiltrating damaged tissue; intravascular neutrophil margination and extravasation can also be found in the early postmortem period, and these leucocytes maintain their motility for about 6 to 8 hours after death. 7,119 In contusions, neutrophils may be visible for up to a month after injury, even in the absence of infection. Neurons in superficial cortical areas are frequently elongated (Fig. 4), appearing to have been stretched (“plastic creep”). They may show cloudy swelling very early, progressing to shrinkage, cytoplasmic eosinophilia, and nuclear pyknosis (red neurons). Red neurons may appear within about 2 hours, sometimes in asynchronous waves. Before dissolution, red neurons may remain for many days and possibly longer, even becoming mineralized (ferrugination) and remaining for years. Phagocytosis (neuronophagia) occurs between 12 and 24 hours and up to 5 days postinjury but is not prominent. At the contusion periphery, red neurons may persist for as long as 5 to 6 months.
Axonal swellings (spheroids) may be observed within several hours to between 24 and 48 hours after injury and may persist for many years. Blood-derived monocytes and reactive resident microglia are seen at 12 to 24 hours, increasing in number up to 7 to 14 days, then declining. They phagocytose erythrocytes and cellular debris and may be found in old contusions for up to 20 years. Lymphocytes are seen at about 3 to 4 days and may be long lasting. Hemosiderin-containing macrophages can appear in small numbers at about 5 days but are generally not obvious until 7 days or later. Intermediate to late reactions include capillary proliferation, which commences at about 5 to 7 days postinjury, entering the contusion as phagocytosis of debris begins, plateaus at about 3 weeks, then gradually declines. Astrocytic reaction with visible cytoplasm is evident at 4 to 6 days, with numbers increasing over ensuing weeks to months and eventually resulting in a glial scar. Some astrocytes contain hemosiderin and lipid debris, reflecting some phagocytic capacity. When the contusion involves arachnoid or dura, fibroblastic proliferation and collagen deposition may occur but are usually minimal. 10,82,103
Intracranial hemorrhages include epidural, subdural, subarachnoid, and brain parenchymal bleeds, and they occur in up to 10% of dogs and cats with mild head injury and >80% with severe injury. 28 They may occur singly or in combination, and while traumatic brain hemorrhage generally results from tearing of blood vessels at the moment of head impact, gradually expanding, delayed hematomas may not be manifest until hours or days postinjury. Small hematomas may completely organize with time, but large hematomas may only partially resolve, persisting with a cystic, fluid-filled center (Fig. 5). During the first 48 hours, the hematoma is composed of clot, which gradually liquefies over the ensuing 2 weeks. 15,66
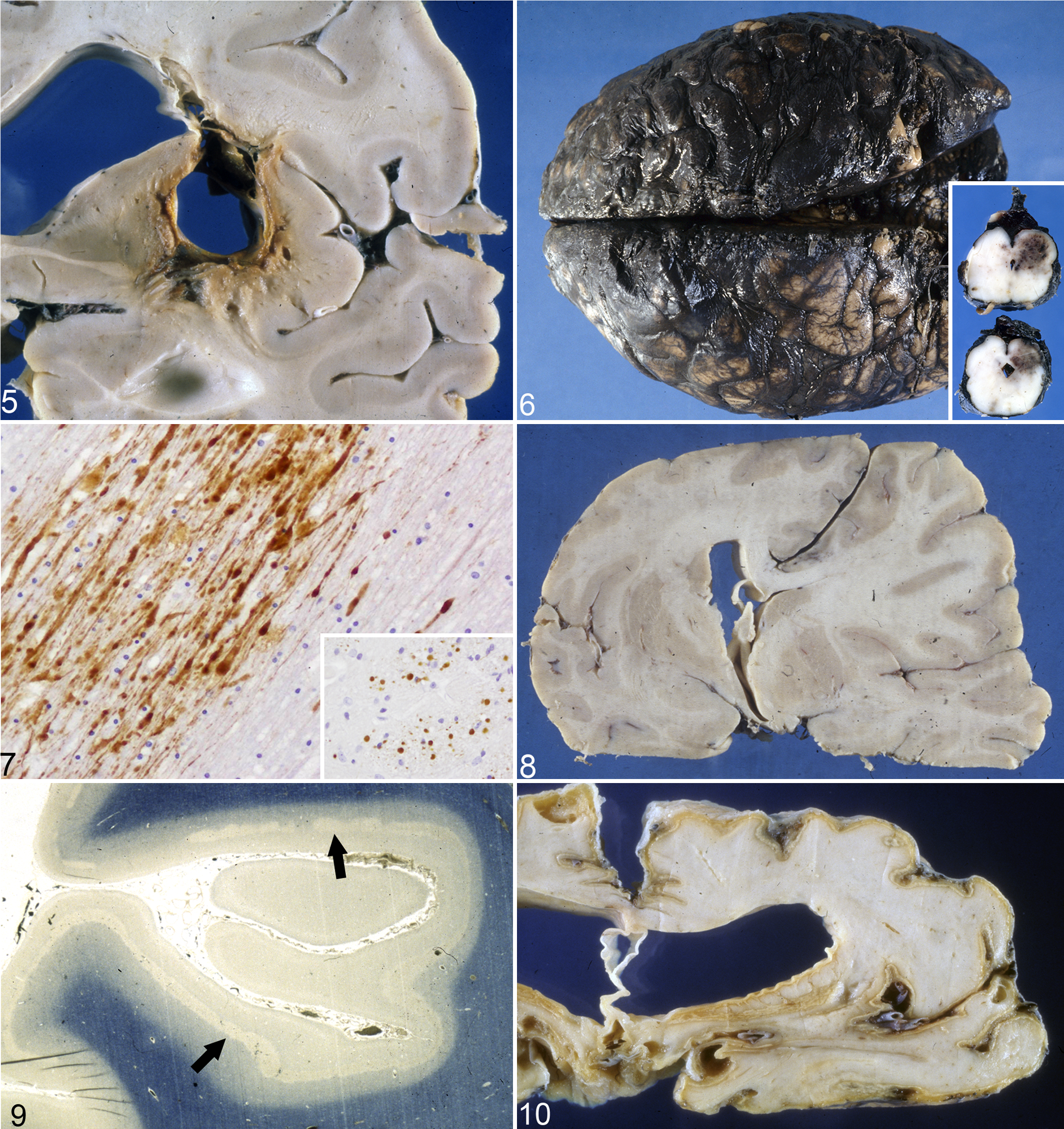
Epidural (extradural) hemorrhages are usually caused by traumatic cranial fractures, and as the hematoma enlarges, it gradually strips the firmly adherent dura from the cranium to form a rapidly developing, circumscribed, ovoid mass that progressively indents and flattens the adjacent brain. They most commonly result from laceration of a meningeal artery of the dura, and since these blood vessels lie within the periosteum, they are prone to injury from skull fractures. Since epidural hemorrhages form under arterial pressure, they most often present clinically within hours or a few days postinjury and are lethal in a high proportion of cases. By contrast, subdural hematomas are of venous origin, with a larger interval between head impact and the onset of clinical signs; unlike extradural hemorrhages, they do not tend to flatten gyri and efface sulci. 15,66 Subdural (intradural) hemorrhages are usually caused by traumatic damage to bridging veins, which drain toward the dural sinuses. There is evidence that hemorrhage initially dissects between the 2 loosely fused layers forming the dura, thus creating an intradural hematoma-filled space, and may then penetrate into the potential space between dura and arachnoid. 15 Where these veins lie free in the subdural space, they are thin walled and vulnerable to acceleration/deceleration forces as the brain shifts within the cranium after a head impact. While epidural hemorrhages tend to be confined by dural adhesion to the cranium, subdural hemorrhages spread widely over cerebral hemispheres. Solid clotted blood that is dark red-black denotes acute subdural hemorrhages; mixed clotted and fluid blood, a subacute bleed; and dark chocolate brown, fluid blood, a more chronic subdural hemorrhages. 51,68
SAH is the most common indicator of TBI, but minor SAH can sometimes be difficult to distinguish from artifact due to tearing of subarachnoid vessels at necropsy. SAH is generally more pronounced at the site of leakage, even though blood spreads rapidly in the subarachnoid space (Fig. 6). While generally minor and frequently efficiently resolved, with minimal effects on the underlying cortex (mild subpial gliosis), large SAHs may evolve into significant space-occupying lesions and be fatal. Localized SAHs typically attend cortical contusions and, when large, may obscure the contusion unless the brain is sectioned. Vasospasm of arteries bathed in SAH can cause ischemic parenchymal complications. 15,103
Studies of the timing of microscopic changes in SAH have produced disparate results; accordingly, the following should be interpreted as a guide only. With a survival time of 1 to 4 hours, a few infiltrating neutrophils appear, and erythrocytes begin to lyse. 61,98 From 4 to 16 hours, the neutrophilic reaction intensifies, peaking at about 3 days, 126 then beginning to be replaced by lymphocytes and macrophages. By 2 to 4 days, phagocytosis of erythrocytes and degenerate leucocytes is well developed, but the first appearance of hemosiderin-laden macrophages seems not to occur until a 6- to 7-day survival period. 5 By 7 days, approximately equal numbers of neutrophils and lymphocytes/macrophages are present, and while a few intact erythrocytes are found, they are completely lysed by 9 to 20 days. 74,98 At 3 days, fibroblastic proliferation occurs in the meninges 62 ; fibrosis is observed at 10 days; and collagen deposition is prominent at 35 days. 8
Intraventricular hemorrhage is a frequent complication of TBI and, when produced by severe external head trauma, carries a poor prognosis. 77 Hematomas may also arise in the brain parenchyma, devoid of contact with the brain surface, and are surrounded by an ischemic penumbra. If a human head injury victim dies within minutes of a severe head impact, numerous petechial hemorrhages may be scattered throughout the brain (diffuse vascular injury). 15
AI is believed to be an almost universal consequence of closed head injury, being demonstrated 24 hours after head impact in 90% of human cases, with a peak between 10 and 15 hours postimpact. Acceleration/deceleration of the head is the major cause of AI and tends to be diffusely distributed, whereas AI from direct blows is much milder and more focal. AI is a vital reaction and, as such, can be useful to distinguish intravital from postmortem brain damage and assist a conclusion that, although the victim sustained a blunt force head injury, it was not immediately fatal. The use of more sensitive immunohistochemical techniques has also recently enabled the time of traumatic AI occurrence to be more accurately determined. 1,49,53,90,113,114
Since there is a constant antero- and retrograde movement of various cargoes in axons, they react to any transport disruption by swelling and forming spheroids. The term axonal spheroid is now widely accepted to describe the ovoid enlargements found in injured axons, but they have been termed axonal retraction balls, retraction bulbs, dystrophic axons, axonal torpedoes (if located in Purkinje cells), axonal balloons, axonal swellings, and axonal varicosities. The number and size of axonal spheroids increases over the ensuing 24 hours. 66,108
The timeline of AI can be determined by application of different histologic techniques and the total burden of AI in a given case depends on the sensitivity of the histologic or immunohistochemical technique used. Accumulation of proteins transported by fast axoplasmic transport—including APP, neurofilament proteins, synaptophysin, and ubiquitin—can be used as sensitive, early immunohistochemical markers of AI, but this expression, due to disruption of axoplasmic flow, does not usually imply a particular pathogenesis (eg, traumatic or ischemic hypoxia). APP is normally transported in undetectable quantities but accumulates rapidly in damaged fibers proximal to the site of injury, being visible by light microscopy as early as 1.75 to 3 hours postinjury; only injured axons are labeled (Fig. 7), and some APP-immunoreactive axons may not be irreversibly damaged. 16,17 APP immunohistochemistry has been used to demonstrate AI in domestic animals such as sheep 42,44,78 and pigs. 43 APP-immunoreactive axons increase in number up to 24 hours postinjury, then decline, but can still be found for up to 3 months. Granular APP deposits may be found at longer periods. Ubiquitin-immunopositive AI is apparent at 6 hours postinjury, 103 but AI can be detected only by silver staining after 12 to 18 hours and HE stain after 12 to 24 hours. 29 Antibodies targeted to the 68-kDA subunit of neurofilament protein show immunopositivity within 60 minutes postinjury. 103 At 48 hours, microglial hyperplasia occurs, 48 and by 5 days, microglial clusters are found around axonal spheroids and begin phagocytosis. 47 However, microglial nodules are not specific for TAI, being also found, for example, in viral encephalitides. At 8 days, there is reactive astrogliosis, 47 but this is not concentrated around spheroids. AI of myelinated axons is usually attended by demyelination (myelin loss). Some contend that it is possible to time an injury from the size of APP-labeled axonal swellings, but axons of different sizes have different cytoskeletal responses to the same magnitude of injury. Thus, the size of an axonal swelling depends partly on the diameter of the injured axon from which it arose and partly on systemic and intracranial pathophysiology. 48
While complete mechanical transection of axons (primary axotomy) can occur after severe head injury, AI more commonly evolves over time (secondary axotomy), leading to eventual disconnection. In most cases, secondary axotomy is delayed until several hours after the initial insult, eventually resulting in disconnection of the proximal from the distal segment, the latter undergoing Wallerian degeneration. 24,91 Moreover, not all damaged axons are irreversibly injured, and some may recover, thus providing a potential window for therapeutic intervention. Calcium influx is important in the pathogenesis of traumatic AI, with the increased intra-axonal calcium activating calpain proteolytic activity and leading to microtubular disorganization and loss, with impaired axonal transport. 130
AI may be focal or diffuse, and the total burden of AI in a given brain is a combination of mechanical deformation (traumatic) and ischemic-hypoxic (vascular) injury. The latter is due to a failure of cerebral perfusion and constitutes one of the major secondary insults after a head injury, particularly when the intracranial pressure (ICP) is high. Since AI demonstrable by APP immunohistochemistry requires a posttraumatic interval of several hours, the possibility that ischemia-hypoxia was also operative during this period cannot be excluded; longer survival times may assist the differentiation of traumatic from vascular AI. 48 Scattered or groups of APP-immunoreactive axons, especially involving a single white matter bundle, may suggest a traumatic etiology. Moreover, a wavelike pattern of APP immunoreactivity seems to be produced by acceleration/deceleration responses to impulsive head rotation, whereas irregularly shaped aggregates of injured axons favor an ischemic-hypoxic insult. 109 While current histologic techniques are often unable to distinguish between these 2 types of AI, that due to ischemia probably constitutes a large proportion of the total AI in fatal human TBI cases. 15
Diffuse AI (DAI) is widely distributed but, despite this designation, not diffusely so in the brain, as it tends to occur multifocally at certain selective neuroanatomic sites. The clinical correlate of severe DAI is unconsciousness at the moment of head impact, and while these patients may remain comatose, there is a spectrum of DAI, with some victims experiencing a lucid interval before coma develops and with others showing neurologic recovery to varying degrees. DAI impedes consciousness by damaging white matter tracts connecting the cerebral hemispheres to the brainstem activating centers and, in more severe neurotrauma, may additionally injure these neuroanatomic sites. 14
Diffuse brain injuries that disrupt the awake state by injuring pathways from the brainstem reticular activating system to the cerebral cortex may result in concussion, defined as a clinical state characterized by immediate impairment of neural functions, such as changes in consciousness and disturbance of vision, motion, and sensation due to mechanical forces. Classically, concussion was conceived as a reversible syndrome without detectable pathology, but this has evolved into a concept of graded concussion. There is temporary confusion with retention of consciousness at the mildest end of the concussion spectrum, with more severe concussion including loss of consciousness and posttraumatic amnesia. Some grades of concussion are also attended by a degree of permanent axonal damage, and with repeated head injuries, the functional impairment may be more severe and prolonged (cumulative concussion). 15,121
Since AI is a critical component of TBI and a major determinant of clinical outcome, it is pertinent to briefly examine the mechanism and consequences of disrupted axonal transport. Axonal transport is composed of 3 basic components: cargo, including organelles, mitochondria, and cytosolic and cytoskeletal proteins; motor proteins that move the cargo—kinesin and dynein; and tracks upon which the cargo travels, mainly microtubules. Traumatic damage to any of these elements, as well as impairment of energy supply to motors from mitochondrial injury, can lead to neurologic disturbance. However, while disruption of axonal transport may contribute to the development and progression of neurologic signs after TBI, the precise causal relationship remains unclear. Some cargoes that are normally transported along axons can accumulate, but whether these aggregates are the cause or a consequence of axonal transport disruption is debated. 12,55,127
The axon, being largely devoid of any biosynthetic capability, is dependent on the transport of essential nutrients produced by the neuronal cell body, and the long length of many axons renders them vulnerable to transport deficits. The motor proteins kinesin and dynein move cargoes in an antero- and retrograde direction, respectively. Anterograde transport conveys myriad proteins, lipids, and polysaccharides bound to organelles and vesicles toward the synapse. Retrograde transport facilitates the removal of misfolded and recycled proteins and effete organelles to prevent a buildup of toxic aggregates, and it enables signaling from the periphery of the axon to permit neurons to respond to trophic influences and stressful stimuli. 55,127
After attachment of kinesin motors to a cargo, kinesin converts the chemical energy derived from ATP hydrolysis into mechanical work, allowing it to “walk” along microtubules, taking consecutive steps along these tracks without dissociating (termed processivity). Dynein also “walks” in a similar manner, but its “stepping” is more irregular. Axonal transport has conventionally been classified as either “fast” or “slow,” but in fact, all motor proteins are “fast,” and the overall velocity of a particular cargo is determined by the length of pauses between movements when it is dissociated from its motor protein. 19,54,63 Regulation and cooperation between motor proteins bound to a cargo are required to increase the force and distance traveled along the axon (same motor type) or to produce bidirectionality of movement (different motor types). Two mechanisms have been proposed to explain directional transport and intermittent pausing during transit, and both are probably operative. First, there may be regulated switching of motor activity such that only 1 type of motor is active at a given time; second, there may be a “tug of war” between opposing motors, the final direction of movement being determined by the dominant motor. 19,54,63
When axonal swellings or spheroids form after traumatic disruption to axonal transport, most are long lasting and remain stable in size and location, but some become larger by fusing with their neighbors, suggesting retention of some axoplasmic flow. Moreover, spheroid formation may be reversible under some circumstances, leading to functional axonal recovery. 19,54,63
In addition to AI, dendritic injury in the form of loss of microtubule-associated protein (MAP-2) has been demonstrated following TBI. MAP-2 is an immunohistochemical marker of dendritic damage, as it is principally expressed in these processes. 87 A diffuse microglial reaction, which can be diffuse or localized to the lesion and which follows a relatively constant clinical course, may be demonstrated after TBI through ionized calcium-binding adaptor molecule 1 (lba1) as an immunocytochemical marker, although, like other microglial markers, it stains only subsets of these glia. There is almost invariably astrogliosis—that is, reactive astrocytes showing increased glial fibrillary acidic protein (GFAP) immunopositivity and an often expanded, eosinophilic cytoplasm due to uptake of extravasated plasma protein. 25,101
Secondary Brain Damage
Secondary or delayed TBI occurs as a complication of the different types of primary brain damage. It develops over hours, days, or weeks after the initial insult and exacerbates the already altered homeostasis of the injured brain. There is evidence of secondary brain injury at autopsy in 70% to 90% of all fatally head-injured human patients. The complex interplay of vascular, cellular, and biochemical cascades leads to ischemic-hypoxic damage, cerebral swelling (Fig. 8), and the results of elevated ICP, such as distortion, shift, and herniation of the brain, hydrocephalus, and infection. Posttraumatic events may involve both neuroprotective and autodestructive/neurotoxic cascades. 15,23
While primary brain damage is produced by mechanical forces operating at the moment of head impact, most brain injury evolves as a progressive cascade of delayed, secondary events that are complications of the initial injury. 23,56
Significant neurologic dysfunction occurs only when hypoxia is combined with perfusion failure (termed ischemic-hypoxic encephalopathy). When an area of brain is perfused, even though the blood is hypoxic, damage is generally less than that if there is blood flow stasis. Ischemia is particularly damaging to the brain because, in addition to energy and nutrient deprivation, there is a concomitant failure to remove potentially toxic metabolic wastes. 11
Ischemic-hypoxic injury is the most common cause of cerebral necrosis and is usually related to cerebral perfusion failure. It can be focal or global and incremental, transient, or permanent. Focal (regional) ischemia occurs when impaired cerebral blood flow leads to reduced perfusion within the supply area of the affected vessel and the collateral circulation is inadequate. It impedes the delivery of substrates, especially oxygen and glucose, and the final expression is reduced availability of ATP. This leads to membrane pump failure, activation of calcium channels (eg, N-methyl-D-aspartate) with an influx of calcium ions (which are neurotoxic), cellular swelling, and death by necrosis or apoptosis, the latter occurring particularly when ischemia is brief. Calcium ions are an essential messenger in many cellular processes but are also often the final common pathway to cell death. Excessive increases in calcium levels severely impede virtually all anabolic and metabolic cell functions in the brain. 130 Further ischemia may result in free radical release, damaging phospholipid cell membranes and resulting in an additional influx of calcium and water. Calcium influx is also abetted by the release of excitatory amino acids, such as glutamate and aspartate. Global cerebral ischemia, for example following cardiac arrest, is induced when cerebral perfusion pressure (mean arterial blood pressure minus ICP) falls below a certain threshold because of either increased ICP or reduced arterial blood pressure. Global ischemia will eventually compromise vital brainstem centers, leading to cardiorespiratory arrest. One of the most important sequelae of ischemia-hypoxia is cerebral edema, which elevates ICP. In addition, if there is reestablishment of the circulation after a protracted period of ischemia-hypoxia, even more severe neural damage can ensue (ischemia reperfusion injury), which tends to be heterogeneous and random in distribution. 11,15,56
The morphologic expression of ischemia-hypoxia is highly variable and unpredictable, in part because the brain is not uniformly affected. There may be persistence of a local blood supply when the circulation in most parts of the brain has ceased. Moreover, certain regions are selectively vulnerable, and there is variability in neuronal sensitivity to ischemia-hypoxia within a given vulnerable neuroanatomical region, since an ischemic-hypoxic insult is a time-dependent process and does not involve all neurons simultaneously. There is a spectrum of resulting tissue damage, varying from selective neuronal necrosis to pannecrosis (infarction). Selective neuronal necrosis is believed to be a type of excitotoxic injury, secondary to the uncontrolled release of the excitatory neurotransmitter glutamate, which preferentially damages dendrites and initially spares neuronal perikarya and axons. 56,57
Ischemic-hypoxic events produce tinctorial changes (hypereosinophilia) in the neuronal cytoplasm, termed neuronal red cell change, acidophilic (or eosinophilic) change, or acute ischemic cell change, the latter term better avoided since this neuronal change is not restricted to ischemic conditions. The middle laminae of the cerebral cortex are particularly vulnerable and usually manifested as a pseudolaminar pattern of necrosis (Figs. 9, 10), with >1 lamina affected; this is especially common in the depths of the sulci. Cerebellar Purkinje cells and the hippocampal pyramidal layer are also highly vulnerable, and regions of central gray matter (eg, caudate-putamen, thalamus) are also commonly damaged. In the cerebral cortex, the boundary zones between territories supplied by cerebral arteries are sites of predilection for ischemic injury because they are the last to be perfused. The reason for these selective vulnerabilities is conjectural, but varying vascular patterns and innate biochemical differences (pathoclisis) among neurons have been implicated. At the cellular level, gray matter is more vulnerable to ischemia-hypoxia than white matter and, in descending order, neurons (which show a differential susceptibility), oligodendrocytes, astrocytes, microglia, and vascular endothelium. 11,15,57,66
Estimations of the first appearance of red (acidophilic) neurons in, for example, infarcts and contusions vary widely, together with the time that a red neuron can be interpreted as being irretrievably destined for death. In contusions, many red neurons express APP, probably as an acute-phase stress reaction, indicating that they are still metabolically active. Red cell change in neurons is generally accepted not to be manifest until several hours postinjury, sometimes up to 7 hours, but in very acute ischemic episodes, it can evolve in 1 or 2 hours or even less. Red cell change is more rapid in smaller than larger neurons, and the time course of development of red neurons also varies among species and among brain regions within the same species. Red neurons, for example, develop more rapidly in cerebellar Purkinje cells than any other brain region. Variability in HE staining among laboratories can sometimes impede a definitive diagnosis of neuronal red cell change, together with subtle variations in color discrimination among pathologists. 11,15,57,66
In areas of ischemic-hypoxic damage, the onset of the first microscopic changes is very variable. The infarcted area shows pallor with HE staining, which is accentuated in the surrounding parenchyma. By light microscopy, the earliest change leading to classical red neurons is vacuolation, corresponding ultrastructurally to swollen mitochondria and some dilated cisternae of endoplasmic reticulum. 69 The neuronal cell body then shrinks, and the cytoplasm appears vivid pink to reddish (Fig. 4) with HE staining. 72 These cytologic changes may persist for at least 6 hours in humans. Some red neurons have an irregular cell contour due to swollen astrocytic processes indenting the cell body, but these can sometimes be difficult to distinguish from autolytic change in immersion-fixed brains. After 24 hours of survival, there is usually increased homogenization and loss of cytoplasmic staining intensity (homogenizing cell change), with often nuclear fragmentation. The final stage is a “ghost cell” appearance, with the cytoplasm being markedly reduced; eventually, there is neuronal loss, which is often attended by gliosis. Red neurons may persist for weeks or disappear rapidly, even in a few days, and take longer to disappear in the neocortex (5–7 days or, less commonly, up to 2 weeks) than cerebellar Purkinje cells (often <4 days). Red neurons may remain for years if they attract minerals, becoming encrusted with calcium and iron (ferruginated neurons). There is also usually early pallor and neuropil vacuolation in areas of neuronal red cell change when compared with adjacent unaffected areas. 18
Within 1 to 2 hours after ischemic-hypoxic injury, there is neutrophilic infiltration into necrotic tissue, and within 18 hours, marked microglial proliferation occurs. Macrophages increase in number along the infarct margin, with hypertrophic astrocytes evident after 4 to 6 days. The infarct then liquefies at its center, and necrotic cellular debris is phagocytosed. Mineralization can occur within days. Both necrosis and apoptosis may occur with ischemia-hypoxia and, without molecular biologic techniques, can be difficult to distinguish by routine light microscopy. Apoptotic cells do, however, tend to die as individuals, whereas necrotic cells generally face their demise in groups. Apoptosis commences in these lesions at approximately 4 hours and is demonstrable for approximately 3 days. Around infarcts and contusions, there is an ischemic penumbra, in which the tissue is perfused at a level sufficient to sustain functional and morphologic integrity; parenchyma in the penumbra has the capacity to recover if perfusion improves. 11,15,57,66
Elevated ICP is the most important secondary complication of TBI and is the commonest cause of death after such an injury. It results from space-occupying contusions, hematomas, and the spreading edema (largely vasogenic) surrounding them. At necropsy, the dura should be left as intact as possible, as this allows an assessment of the tenseness of intracranial contents by palpation and the likelihood of increased ICP being present. 70 Raised ICP is usually an indicator of the severity of the underlying brain injury and represents exhaustion of compensatory mechanisms. Initially, displacement of an equal volume of cerebrospinal fluid and venous blood limits the rise in ICP (the Monro-Kellie doctrine), but when no further compensation is possible, ICP can rise rapidly with a concomitant fall in cerebral perfusion and ischemic-hypoxic damage as ICP approaches blood pressure. Although elevated ICP is found in most severe head injuries, marked distortion and herniation of the brain can result from slowly expanding, space-occupying lesions, without significantly increasing ICP. By contrast, very rapid elevations of ICP may cause minimal distortion and herniation. Moreover, when distortion and herniation of the brain are found at necropsy, they do not necessarily indicate the existence of elevated ICP antemortem, as these change might have occurred as part of the compensatory mechanism before there was a significant change in ICP. 15,58,94
Brain swelling after TBI may be due to (1) vascular engorgement (congestion) from arterial dilatation and/or venous obstruction, which can occur rapidly or be delayed by several days, or (2) increased brain water content (edema). With cerebral edema, white matter is softer in consistency and paler than normal, and the usual sharp distinction between gray and white matter is obscured, often with thinning of the overlying cortex. Brain swelling is sometimes suggested by hemispheric asymmetry, as well as the degree of gyral flattening and sulcal narrowing. In cases where a pressure gradient has developed across the foramen magnum, the cerebellar vermis is displaced caudally through the foramen (“cerebellar coning”), and, if severe and protracted, the tonsils may become necrotic. Furthermore, a portion of the occipital lobes can herniate under the tentorium cerebelli, resulting in brainstem compression. Apart from hematomas, edema is the major cause of raised ICP and an important cause of death after TBI. Head injury results in 2 main types of cerebral edema, both of which are usually present in varying combinations and can alter over time but may occur sequentially. Vasogenic (open barrier) edema occurs after blood-brain barrier breakdown, resulting in extravasation of a protein-rich fluid, and it can be demonstrated through albumin immunohistochemistry as a surrogate marker of increased blood-brain barrier permeability. Macroscopically, there is a light green discoloration of the white matter; microscopically, there is swelling of pericapillary astrocytic foot processes and oligodendroglial cytoplasm, together with spread of extravasated fluid in the extracellular space of the white matter. Vasogenic edema can occur within a few minutes of injury or develop after several days around contusions and intraparenchymal hemorrhage. Cytotoxic (closed barrier) edema occurs with ischemia-hypoxia from loss of high-energy phosphates and derangement of the sodium/potassium pumps. Water then enters cells, especially astrocytes; calcium channels open; and there is an influx of these ions. Cytotoxic edema, which predominates in gray matter, is characterized by astrocytic swelling and enlargement of perivascular and perineuronal spaces due to swelling of astrocytic processes. 93,99
Inflammation (and attendant edema) is one of the major delayed, evolving responses to brain trauma, with recruitment of inflammatory cells during secondary damage cascades. Neuroparenchymal damage elicits nonspecific but complex and programmed cellular and molecular processes constituting inflammation, which may have substantial pathologic consequences and can be an important determinant of the eventual neurologic outcome. The initiation and orchestration of inflammation following TBI is multifactorial, encompassing pro- and anti-inflammatory cytokines, chemokines, adhesion molecules, complement factors, and reactive oxygen and nitrogen species. Neuroinflammatory events are a “double-edged sword,” having dual and opposing roles after TBI. While inflammation is responsible for some of the secondary brain damage and adverse clinical outcomes, it may also be neuroprotective and promote neuroreparative mechanisms. These beneficial and deleterious effects also often differ between the acute and more delayed phases after a head injury. 37,95
Infection of the brain is an ever-present threat when the integrity of the protective skull is traumatically breached. Meningitis is a well-recognized complication after a head injury, and subdural or intracerebral abscesses may form, being more common after penetrating than nonpenetrating head injuries. Another complication of nonmissile head injury is posttraumatic hydrocephalus, which is usually due to impairment of cerebrospinal fluid absorption via the arachnoid granulations owing to the presence of SAH or fibrosis and hemosiderosis of the meninges. However, ventricular dilatation can also be secondary to loss of brain tissue after a severe injury (ex vacuo ventriculomegaly). 66,103
Other sequelae of TBI include a catecholamine surge, which may lead to myocardial necrosis and neurogenic pulmonary edema; coagulopathy, including disseminated intravascular coagulation; large release of inflammatory mediators into the circulation (systemic inflammatory response syndrome); excitotoxic neuronal injury; and convulsive disorders, impaired immune responses, and fluid and electrolyte imbalance. 71,85,117,128
Penetrating, Missile Head Injury
The clinical sequelae of gunshot and other missile wounds to the head are largely determined by the course of the projectile through the skull and brain and the energy released. 30 When a bullet enters the skull, it may travel along its inner curvature and either come to rest without entering the brain or ricochet off the inner table (or other firm structure, such as the falx cerebri or tentorium cerebelli) and penetrate the brain ≥1 times. 45 SAH will be present, and surface contusions may occur around entrance and/or exit wounds. If bullet fragmentation occurs, secondary wound tracks will be produced in the brain, often abetted by indriven skull bone fragments, with the paths of bone fragments providing strong evidence of the direction of the missile. By contrast, the size and configuration of the permanent wound cavity in the brain are poor predictors of the caliber and direction of the bullet. 30,104
High-velocity bullets usually produce a perforating injury, with both entry and exit wounds. By contrast, lower-velocity bullets often produce a penetrating injury, with the projectile being retained within the skull or soft tissues on the contralateral side of the head (Table 1). In human gunshot injuries due to low-velocity firearms, the majority (approximately 70%) are penetrating, while about 30% are perforating. 45 In general, exit wounds are larger than entrance wounds, but there are exceptions. They are both round to ovoid in shape, with exit wounds sometimes being more irregular. Since the cranial bone is composed of outer and inner tables, a penetrating entry wound produces 2 fractures, with that of the outer table almost invariably smaller than the inner table fracture (termed internal beveling). This occurs because the missile penetrates the cranium in a broadening cone. Conversely, if the missile has sufficient energy to penetrate the contralateral skull, the exit fracture will be smaller in the inner than outer table (external beveling). At necropsy, fracture lines extend from the bullet holes into more remote regions of the skull in most cases—sometimes radiating from and, less commonly, unrelated to the entrance hole. 115
The brain damage inflicted by gunshot wounds is largely determined by the amount of KE deposited in the brain by a slowing projectile. The KE transferred to the brain by any missile equals 0.5 × missile mass × (missile velocity) 2 . 4,30 Accordingly, the resulting brain damage is a function of the mass of the missile, and more particularly, its velocity. All moving objects have KE, and when a projectile enters the brain and eventually comes to rest, all of this energy must have been dissipated. This occurs by imparting motion to tissues and fluids, fracturing bones, vaporizing liquids, and generating heat as the missile loses velocity. If the missile is retained within the brain, then all the missile energy will have been deposited in neural tissue. For effective euthanasia of livestock, there are considerable advantages in missile fragmentation and dissipation of KE of the projectile within the cranial cavity, without penetration of the contralateral skull bones, to enhance the severity of the resulting brain damage. Other retardant factors augment brain injury, such as tissue-specific gravity and aerodynamic factors, principally yaw. Yaw is deviation of the long axis of the projectile from its line of flight, with the energy imparted to the parenchyma being greatest when the yaw angle is maximal. 21,22,64,103
There are several mechanisms by which a bullet or other missile damages the brain. Initially, a shock wave moves through neural tissue ahead of the missile at approximately the speed of sound, but it is of short duration and may not be particularly injurious. A path (permanent wound cavity) is then hollowed out by the projectile as it passes through the brain (Fig. 11), causing extensive laceration and crushing of parenchyma. The size of the permanent track, which is always hemorrhagic, is dictated by the diameter of the missile, abetted by any instability or deformation of the projectile. This injury is complicated by the intrusion of bone fragments and portions of lacerated scalp and subjacent soft tissues, and penetrating head injuries have a high risk of infection. Secondary tracks delineating the path of bone fragments are often found, and bullet fragments are frequently entangled with damaged hair follicles acquired during skin penetration. Shotgun injuries produce multiple tracks in the brain, which vary in length according to the KE deposited, and a pellet can be found at the terminus of each track. A bullet imparts momentum to all particles that are struck, and hard particles (eg, bone from the perforated skull, especially the inner table) are driven in varying directions from the main wound path. In permanent wound tracks of the cerebrocortical gray matter, neurons are usually totally destroyed, while those in adjacent tissue are markedly elongated and hyperchromatic (“stretched” or “plastic creep”). Copious intraventricular hemorrhage, with frequent detachment of the choroid plexus, and midline shift and distortion/displacement of the brain are also commonly found. 22,30 Most important, there is a temporary cavity formed in the wake of the missile. The brain is also displaced by gases generated by burning powder when the gun muzzle contacts or is in close proximity to the head. 21,22,31 –34,103
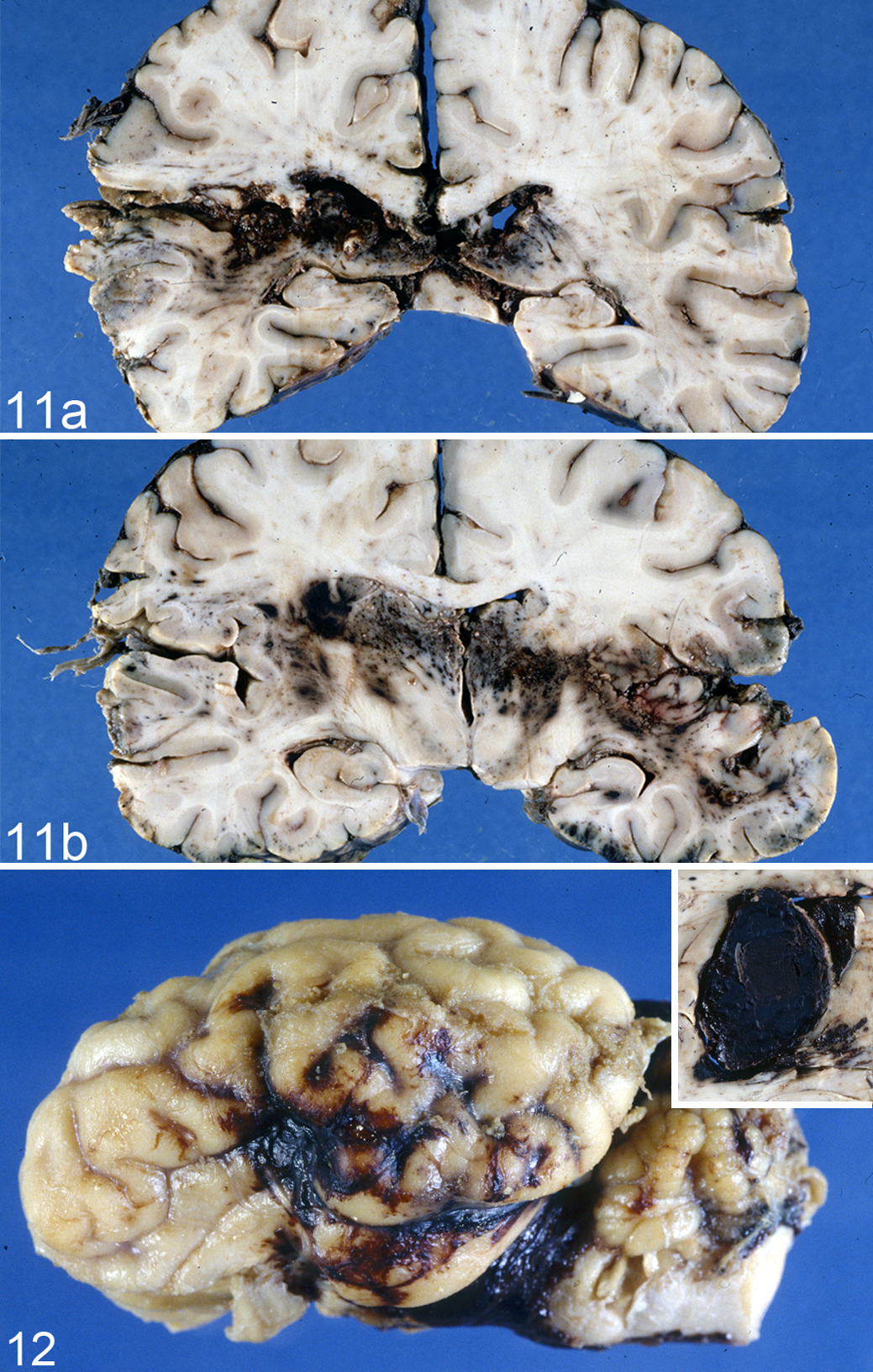
When a missile passes through the brain, KE is transferred to the contiguous tissue, which is propelled radially from the missile track, forming a large temporary cavity. This temporary cavity is mainly responsible for a fatal outcome after missile injury to the brain, especially with a high-velocity projectile, unless vital structures are damaged by tissue destruction in the immediate vicinity of the wound track. The size of the temporary cavity is determined by the velocity of the missile and density of the tissue penetrated. High-velocity (>1000 ft/s) military rifles create a very large temporary cavity, up to 30 times the diameter of the permanent wound track, whereas slower bullets from civilian-type firearms produce a much smaller cavity; stab wounds are of very low velocity and produce only a permanent track. The brain is very sensitive to the stretching effects of cavitation (and the large internal pressures generated in its expanding walls), as it is relatively inelastic. Inelastic tissues such as brain, liver, spleen, and bone are more sensitive to temporary cavity effects than elastic tissues such as muscle, lung, skin, and bowel, with damage further potentiated in brain gunshot wounds by the inelasticity of the constraining skull. 32 When the limit of neural tissue elasticity is attained, the tissue collapses to the point from which it was originally displaced. This cycle may be repeated several times before the disrupted tissue settles around the permanent wound track, enhancing tissue damage. These oscillating movements of the cavity propagate pressure waves widely throughout the brain, resulting in distortion and damage to blood vessels and nerve fibers remote from the primary missile track. 86,105,106
Microscopically, there is destruction of brain tissue in the permanent missile wound track, which subsequently collapses or is compressed by surrounding edema. This permanent wound cavity is surrounded by a hemorrhagic zone, whose shape and extent are determined by the blood supply to the injured region. This bleeding zone is in turn surrounded by a zone of necrosis, which mainly involves neurons and axons, and its formation is secondary to temporary cavitation. More widespread AI also occurs distant to the permanent cavity, although distinguishing the effect of mechanical trauma from secondary ischemic-hypoxic injury can be problematic. In addition, cerebral edema, the most common complication of missile brain wounds, can cause herniation, and open gunshot wounds favor bacterial contamination. The latter may lead to suppurative meningitis and cerebral abscessation; in such cases, the prognosis is extremely poor. 105 –107,120
Gunshot injuries to the head in living victims (vital injuries) must be distinguished from those inflicted postmortem. If hemorrhage has occurred and/or there is a zone of hemorrhage and/or edema around the permanent missile track, postmortem injury is unlikely. Evidence indicating the vitality of a wound includes reactive changes, such as emigration of leucocytes, APP-immunopositive injured axons, and perineuronal/vascular edema. The signs of cerebral edema may appear with survival of 20 to 30 minutes.
The following histologic criteria have been used to date/age cerebral gunshot wounds: neuronal vacuolation and shrinkage (morphologic changes commencing at 30 minutes), white matter edema (50 minutes), leucocyte emigration (70 minutes), oligodendrocyte swelling (90 minutes), macrophage reaction (17 hours), and erythrocyte-containing macrophages and neuronophagia (19 hours). 100 As in nonmissile brain injury, APP-immunopositive AI can be detected as early as 1.75 to 3 hours postinjury. 15
With forensic cases involving stabbing of the brain by knives or instruments such as screwdrivers, ice picks, chisels, pens, nails, and crossbow bolts/arrows, the head is struck at low velocity. Penetration occurs only where the skull bone is very thin, for example, in the orbit and temporal bone; where the bone is thicker, the object is more likely to break than penetrate the skull. In cases of skull penetration, brain damage is limited to a permanent wound track, without temporary cavitation, but there may be associated intracranial and/or intracerebral hemorrhage. 30
Captive bolt stunning of domestic livestock is the most frequently used method of euthanasia in abattoirs and for humane killing of moribund animals. It may also be used to kill animals illegally and has, on occasion, been involved in human suicides. Both penetrating and nonpenetrating mushroom-head captive bolt pistols (humane stunners) are used, 38,40 and these head wounds must be differentiated from those produced by bullets from higher-velocity weapons.
With a penetrating captive bolt projectile, the inner and outer tables of the skull are fractured, with a subjacent impact contusion (Fig. 12). By contrast, nonpenetrating mushroom-head stunners sometimes fracture only the outer table of the skull, although impact contusions are frequently present. Focal vascular and AI is more severe with the penetrating weapon, while such neural damage is more widely distributed after a nonpenetrating head impact. The latter causes rapid angular (rotational) acceleration of the head after impact of the large mushroom-shaped stunner head with the skull, while the area of the head impacted by the cylindrical bolt of the penetrating stunner is smaller and movement of the head correspondingly reduced. 38,40 However, in the only study (in sheep) to compare brain damage in livestock resulting from penetrating and nonpenetrating captive bolt stunning, the overall severity and mix of focal and diffuse injury suggested that both weapons were an acceptable form of euthanasia. 42 Brain damage is additionally influenced by the region of the head subjected to impact from a nonpenetrating mushroom-head stunner, being greatest in sheep with a frontal impact, followed by an occipital, then a temporal approach. 44
In summary, one of the most common forensic conditions encountered by veterinary pathologists is traumatic head-brain damage—either blunt, closed, nonmissile type or penetrating, open, missile type. These neural injuries are characterized by the great diversity and complexity of the many distinct but interactive constitutive lesions. Neurotrauma, unless immediately fatal, is a dynamic, evolving process, and even if lesions appear to be focal, the effects of trauma are usually more diffusely distributed, structurally and functionally, in the brain and dictated by underlying biomechanical events.
Footnotes
Acknowledgement
I thank Professor Peter Blumbergs, consultant neuropathologist, SA Pathology, Hanson Institute Centre for Neurological Diseases, for assistance with this review and kindly providing the human TBI images.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.