
Abstract
The novel subgroup J of avian leukosis virus (ALV-J) has emerged as a significant cause of myeloid neoplasia and weight suppression in broiler chickens. We investigated viral tropism using RNA in situ hybridization (ISH) in naturally infected chickens. Formalin-fixed tissues were collected from 12-day-old embryos (seven infected, two control) and from 0-week-old (four infected, one control), 3-week-old (five infected, one control), 6-week-old (five infected, one control), and 9-week-old (10 infected, two control) chickens naturally infected with ALV-J in ovo. A 636-base antisense riboprobe complementary to the 3′ and 5′ ends of the pol and env viral genes, respectively, was constructed. Strong positive staining was present in cardiac myocytes, Purkinje fibers, vascular and pulmonary smooth muscle, renal glomeruli, distal tubules, and pituitary glands. Light staining was present in gastrointestinal smooth muscle, thyroid and adrenal glands, and follicular medullae in the cloacal bursa. Staining was not present in any hematopoietic precursors. Tissues from newly hatched chicks exhibited the strongest and most consistent staining, whereas staining in embryos was minimal. RNA ISH confirmed the presence of ALV-J-specific nucleic acid within cytoplasmic inclusions in cardiac myocytes, Purkinje fibers, pituitary glands, and renal glomeruli. Viral tropism for cardiac myocytes and Purkinje fibers may relate pathogenetically to the cardiomyopathy and congestive heart failure described in index chicken flocks infected with ALV-J. Viral tropism for endocrine organs may relate pathogenetically to the weight suppression associated with infection.
Avian leukosis viruses (ALVs) are a distinct group of simple type C retroviruses well known for their ability to induce neoplasia in mature chickens. Ten subgroups, A through J, are currently recognized. 25 B-cell lymphosarcoma originating in the cloacal bursa predominates in natural infection, but a variety of other tumor types also have been described. 25 Although the vast majority of the research on these retroviruses has focused on their oncogenic properties, several studies have focused on their nonneoplastic effects and pathogenesis in younger chickens. Viruses in ALV subgroups A, B, C, and D produced stunting in growing chickens associated with a lymphocytic thyroiditis and lowered circulating thyroxine levels, with the most severe effects associated with ALV subgroup C. 7,8 RAV-1—a subgroup A ALV—induced myocarditis, right-sided heart failure, and ascites syndrome in immature chickens and was associated with the formation of viral matrix inclusion bodies in cardiac myocytes. 15,16 An ALV produced by a lymphoid leukosis cell line also induced ascites when inoculated into immature chickens. 17 Subgroup B helper viruses induced severe immunosuppression in young chickens by causing macrophage dysfunction associated with the persistent synthesis of proviral DNA. 12,28
The novel subgroup J of ALV (ALV-J) has emerged as an important pathogen of meat-type chickens. 4,24,27 Similar to other ALVs, ALV-J induces neoplasia primarily in mature chickens, but unlike infections with other subgroups, myeloid tumors predominate, and ALV-J has a higher tropism than RAV-1 for cells of the myelomonocytic lineage rather than lymphocytes. 1,2,26 The lowered tropism for lymphocytes has been suggested as the cause for a predominance of myeloid tumors instead of lymphosarcoma in natural ALV-J infection. 2,3 The nonneoplastic effects of ALV-J in poultry, particularly in younger chickens, have not been well described. Initial isolates of ALV-J came from meat-type chicken flocks with a high prevalence of ascites and retroviral particles present in the myocardium. 20,24 Severe body weight suppression, evident as early as 1 week of age, has been seen in meat-type chickens following natural congenital infection with ALV-J. 32 The pathogeneses of these effects have not been elucidated.
RNA and DNA in situ hybridization (ISH) techniques have been utilized for detecting the nucleic acid of multiple retroviruses to investigate pathogenetic mechanisms. For example, the widespread tissue distribution of Jembrana disease virus has been implicated as the cause for the high circulating viremia during the acute febrile phase of the disease. 9 Rapid infection of local lymphoid tissue and Langerhans cells detectable by ISH has been implicated as a mechanism for the rapid establishment and dissemination of both human and simian immunodeficiency viruses (HIV and SIV, respectively). 11,21,29,31 ISH has also been used to identify the major source of HIV virions during infection. 23,35 HIV RNA detected by ISH in megakaryocytes has been implicated as a cause for immune thrombocytopenic purpura in HIV patients. 19 ISH has been used to detect both human T-cell leukemia virus type 1 (HTLV-1) and SIV in the central nervous system of experimental hosts with neural lesions, implicating these viruses as having a direct or indirect role in these syndromes. 5,18 The tissue tropism of most ALVs has been largely unexplored, with the exception of ALV-C. Ducks inoculated with ALV-C as embryos and that remained persistently viremic harbored more ALV-C RNA within multiple tissues, as detected by ISH, than did similarly inoculated ducks who were not viremic. 34
The tropism of ALV-J has been explored in experimentally infected hosts. Both immunohistochemical detection of group-specific antigen and ISH detection of viral transcripts have been used to determine viral tropism in chickens experimentally infected with ALV-J in ovo. 2,3 Group-specific antigen and viral nucleic acid expression were greatest in heart, kidney, adrenal gland, and proventricular glandular epithelium. Although the experimental model of ALV-J infection reliably reproduces the neoplastic manifestations typical of natural infection, studies using this model have not reported any other adverse manifestations, such the body weight suppression described in chickens naturally infected in ovo. 32 Differences in clinical manifestation may be due to differences in tissue tropism between natural and experimental infection. In studies using experimentally infected chickens, only a limited number of endocrine tissues were examined, which another study showed to be frequent sites of ALV replication. 13 In the present study, we investigated the pathogenesis of ALV-J infection in chickens naturally infected in ovo by determining the tissue distribution of viral transcripts in these chickens at 12 days embryonation and at 0 (newly hatched), 3, 6, and 9 weeks of age.
Materials and Methods
Tissue collection
Broiler chickens (19 males, 10 females, nine embryos) were produced from established ALV-J–positive and ALV-negative breeder flocks. Their ALV and ALV-J status was verified at hatch by both ALV p27 enzyme-linked immunosorbent assay (ELISA) and reverse transcription polymerase chain reaction (RT-PCR) amplification of viral genomic RNA extracted from plasma as previously described. 30,32 For broilers collected as embryos, viral RNA was extracted from a 250-µl sample of amniotic fluid using the same method as for plasma samples. Broilers were considered negative for exogenous ALV if the ELISA results were negative. Broilers were considered congenitally infected with ALV-J if results of both the ELISA and RT-PCR were positive. Broilers were raised as described previously and were weighed weekly. 32 ALV-J–infected broilers and uninfected broilers serving as negative controls were collected at 12 days embryonation (seven infected, two control) and at 0 weeks (four infected, one control), 3 weeks (five infected, one control), 6 weeks (five infected, one control), and 9 weeks (10 infected, two control) of age. Specimens of pituitary gland, thyroid gland, parathyroid gland, adrenal gland, spleen, thymus, bursa of Fabricius, bone marrow, heart, sciatic nerve, skeletal muscle, lung, proventriculus, duodenum, pancreas, kidney, and gonad were collected from each chicken and fixed by immersion in 10% neutral buffered formalin for 15–24 hours. To facilitate tissue collection from 0-week-old chicks and embryos, whole body sections were collected, which frequently included additional tissues. Yolk sac was also collected from 0-week-old chicks and embryos. After fixation, specimens were either processed immediately or immersed in diethyl pyrocarbonate (DEPC)-treated phosphate-buffered saline (PBS) for no more than 24 hours and then processed. Tissues were processed, embedded using routine methods, and sectioned at 3 µm for both hematoxylin and eosin (HE) staining and ISH.
Riboprobe synthesis
RT-PCR amplification of the 3′ and 5′ ends of the pol and env viral genes of ALV-J was performed using the primers H5 and H7 as described previously. 30,32 ALV-J genomic RNA isolated from the plasma of the infected breeder flock was used as the template for RT-PCR. The PCR product was sequenced (Molecular Genetics Facility, University of Georgia, Athens, GA), and sequence analysis revealed 99.3% identity with the corresponding portion of the provirus of the prototype ALV-J isolate HPRS-103 (GenBank accession No. Z46390). The PCR product was ligated into the pGEM-T Easy transcription vector and cloned using JM109 competent Escherichia coli following the manufacturer's recommendations (Promega, Madison, WI). The vector was purified (QIAprep Spin Miniprep Kit, Qiagen, Valencia, CA) and digested for 2 hours at 37 C using 20 units of the restriction enzyme NcoI (Roche Molecular Biochemicals, Indianapolis, IN) per 1–3 µg of DNA. Following digestion, a 636-base antisense digoxigenin-labeled riboprobe was synthesized using SP6 polymerase (DIG RNA Labeling Kit SP6/T7, Roche Molecular Biochemicals). Digoxigenin incorporation in the riboprobe was confirmed using a dot blot. The riboprobe was suspended in DEPC-treated water with 40 units of RNase inhibitor (Promega) and stored at −80 C.
RNA ISH
RNA ISH was performed as described previously with minor modifications. 6 Tissue sections were deparaffinized in Hemo-D (Fisher Scientific, Pittsburgh, PA), rehydrated with 5 mM MgCl2 in PBS, and digested with 25 µg/ml proteinase K for 15 minutes at 37 C. Hybridization was performed at 42 C overnight with approximately 25 ng of probe in 5× standard sodium citrate (SSC), 50% formamide, 1% N-lauroylsarcosine, 0.02% sodium dodecyl sulfate (SDS), and 5% blocking reagent (Roche Molecular Biochemicals) per slide. Tissues from ALV-J–infected chickens were also incubated with a digoxigenin-labeled riboprobe complementary to the N gene of an isolate of the New Jersey strain of vesicular stomatitis virus (VSV) or incubated without any probe under identical conditions to serve as controls. 10 After hybridization, slides were stringently washed in decreasing concentrations of SSC and SDS. Slides were incubated for 2 hours at 37 C with alkaline phosphatase–conjugated anti-digoxigenin antibody (Roche Molecular Biochemicals) and then developed with the chromagen nitroblue tetrazolium and 5-bromo-4-chloro-3-indolylphosphate for approximately 60 minutes. Slides were counterstained with Mayer's hematoxylin.
Results
Chickens congenitally infected with ALV-J exhibited severe body weight suppression at 3, 6, and 9 weeks of age but not at 0 days of age. A fibrosarcoma was present in the ventral cranium of one 9-week-old congenitally infected chicken, but no other adverse clinical manifestations or tumors were noted in any chickens. Staining in tissues from congenitally infected chickens indicating hybridization with the ALV-J–specific riboprobe appeared as a red-brown precipitate in tissue sections. Tissues from congenitally infected chickens that exhibited staining are summarized numerically for each age group and for each tissue in Table 1. Because of difficulties in collecting each tissue from embryos and chickens, some variation occurred in the total number of birds examined for each tissue. Staining was not observed in any tissues from ALV-J–infected chickens that were hybridized with the VSV riboprobe or in tissues incubated without any riboprobe. No differences were observed in the staining intensity between tissues processed immediately after formalin fixation and tissues maintained in DEPC-treated PBS prior to processing.
Results of in situ hybridization for avian leukosis virus subgroup J in congentially infected 12-day embryos and 0-, 3-, 6-, and 9-week-old chickens. Results are expressed as the number positive over the total number examined for each tissue. ∗
∗ NE = not examined.
Cardiovascular tissues
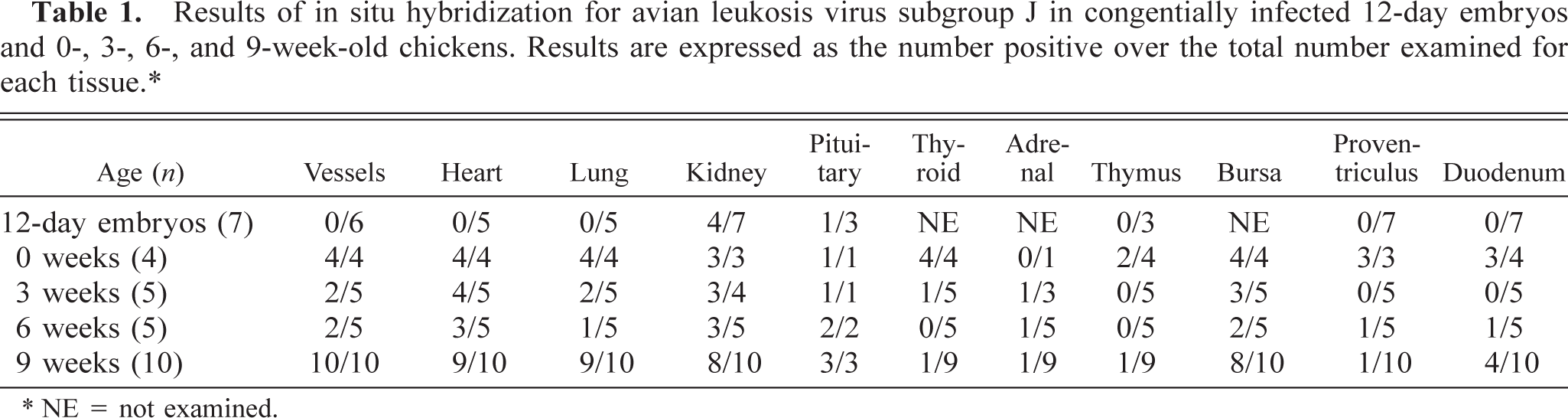
Staining was present in the cytoplasm of cardiac myofibers and Purkinje fibers (Fig 1a). The pattern of staining was variable, with multifocal to coalescing staining throughout numerous myofibers in 0-week-old chicks and discrete clusters of staining in the cytoplasm of myofibers and Purkinje fibers in 3-, 6-, and 9-week-old chickens. Staining was not present in the hearts of any embryos. HE-stained tissue sections from 3-, 6-, and 9-week-old chickens had rare basophilic intracytoplasmic inclusions in both myofibers and Purkinje fibers. The inclusions were round to elliptical and most frequently occurred in chains, often extending the length of the myofiber (Fig 1b). The inclusions were identical in size and shape to areas in myofibers and Purkinje fibers stained using the ALV-J–specific riboprobe. Staining was also present in the cytoplasm of individual smooth muscle fibers of the tunica media of numerous large vessels. Staining was present in all 0- and 9-week-old chickens, fewer 3- and 6-week-old chickens, and none of the embryos.
Myocardium of left ventricular free wall; 9-week-old chicken. Fig. 1a Viral nucleic acid is stained in the cytoplasm of several cardiac myofibers. ISH with ALV-J antisense riboprobe, Mayer's hematoxylin counterstain. Bar = 20 µm. Fig. 1b Several cardiac myofibers contain chains of round to elliptical intracytoplasmic viral inclusions extending the length of the myofiber. HE. Bar = 20 µm.
Pulmonary tissues
Staining was present in atrial septal epithelium and smooth muscle of 0-week-old chicks and in atrial smooth muscle of 3-, 6-, and 9-week-old chickens. Staining was present in all 0-week-old chicks and most 9-week-old chickens but in fewer 3- and 6-week-old chickens. Pulmonary staining was not present in any embryos.
Renal tissues
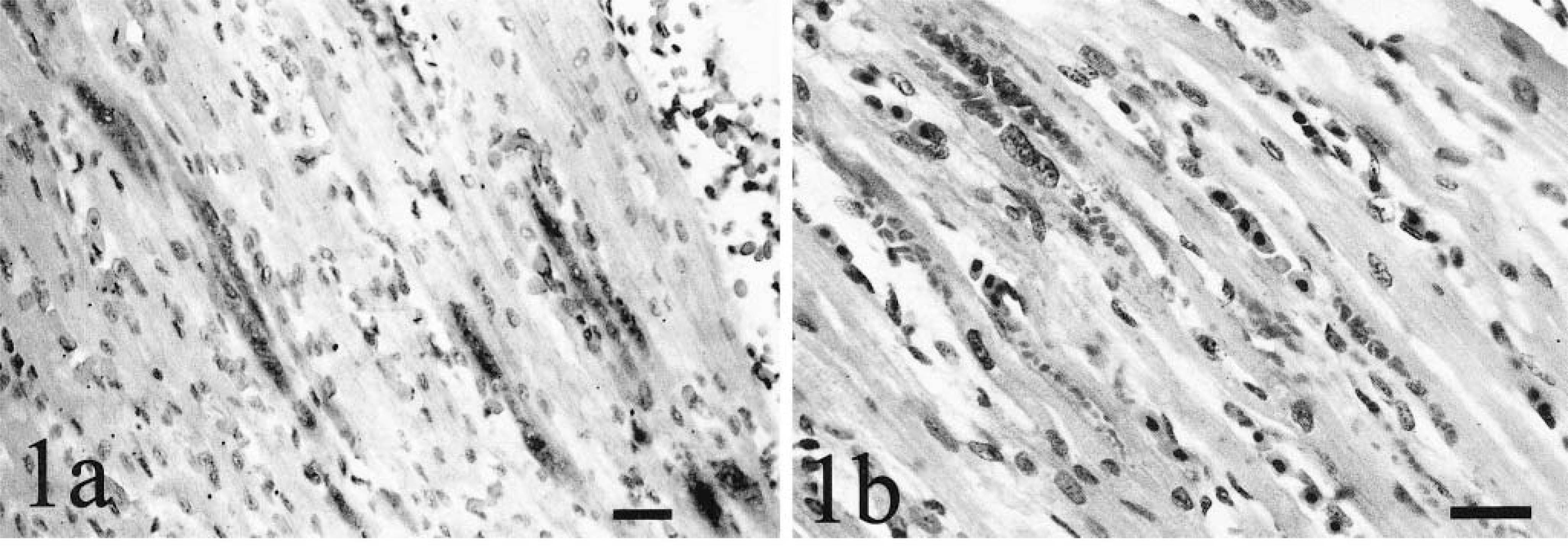
Staining was present in the kidneys of all 0-week-old chicks and in most embryos and 3-, 6-, and 9-week-old chickens. Staining was most frequent in glomerular visceral epithelial cells and less frequent in mesangial cells and tubular epithelium (Fig 2). Distal tubules stained more frequently than proximal tubules. Staining in some distal tubular epithelial cells was polar, with greater intensity in the basal regions and lighter staining in apical regions. In two embryos, staining also was present within tubules and glomeruli of the mesonephros. In the other embryos and all hatched chickens with remnant mesonephric tissue, staining was present only in the metanephros. In one 9-week-old chicken, staining also was present within medullary loop epithelium. Basophilic intracytoplasmic inclusions in visceral epithelial cells were visible in HE-stained sections from 3-, 6-, and 9-week-old chickens and corresponded to areas of ISH-specific staining.
Kidney; 9-week-old chicken. Viral nucleic acid is stained in glomerular visceral epithelium. ISH with ALV-J antisense riboprobe, Mayer's hematoxylin counterstain. Bar = 20 µm.
Endocrine tissues
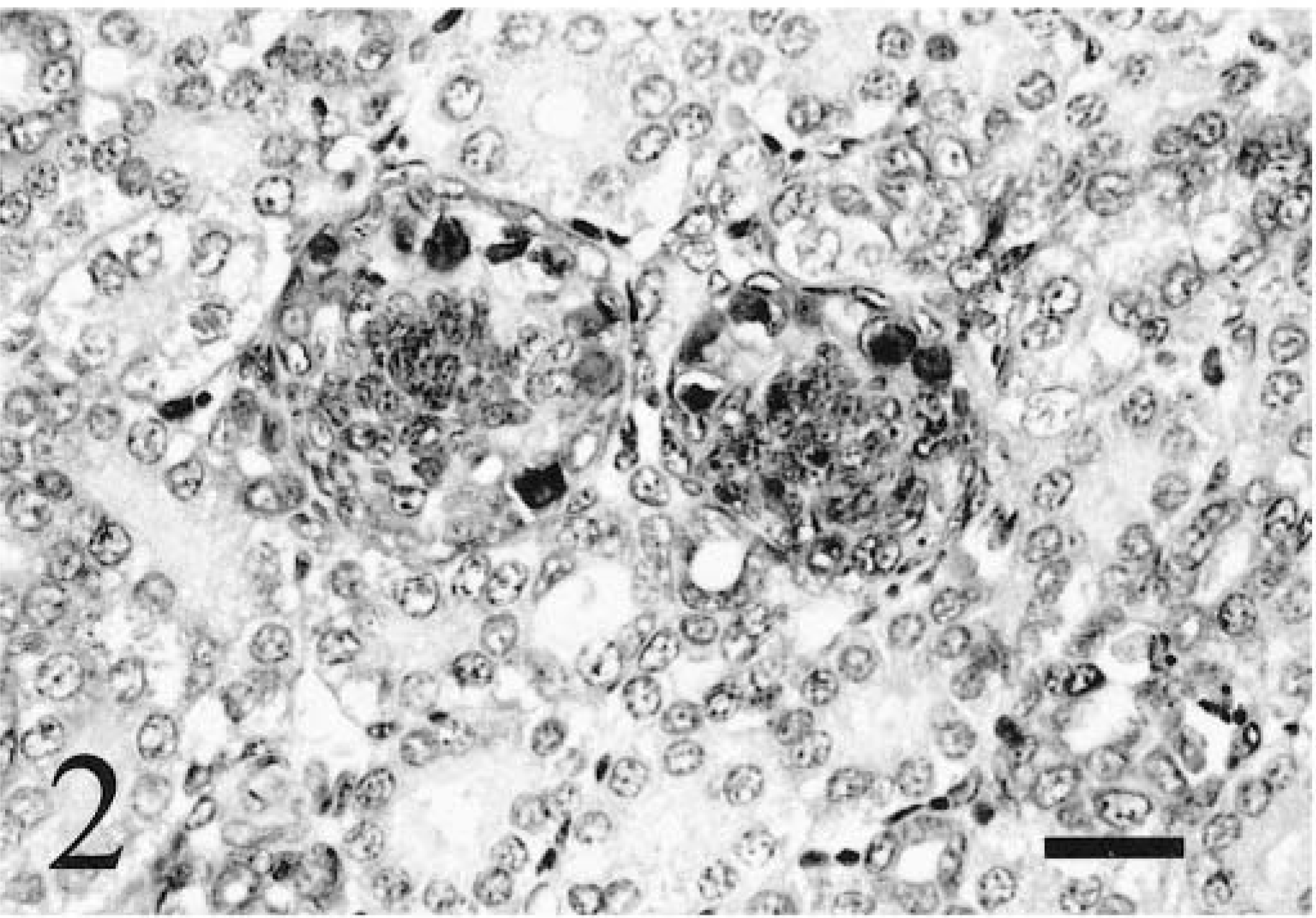
Staining was present in pituitary, thyroid, and adrenal glands of most chickens examined. A limited number of pituitary glands were examined for each age class because of difficulty in collecting this tissue, but all of these tissues in hatched chickens exhibited staining in the adenohypophysis (Fig. 3). One pituitary gland from an embryo also exhibited staining in the adenohypophysis. Rarely, basophilic intracytoplasmic inclusions were present in the adenohypophyses of 6- and 9-week-old chickens. All thyroid glands exhibited follicular epithelial staining in 0-week-old chicks, but only a few glands exhibited staining in 3- and 9-week-old chickens, and none were stained in 6-week-old chickens. Follicle size was extremely variable and markedly smaller in chickens congenitally infected with ALV-J when compared with ALV-negative control chickens. Staining was more prominent in the epithelium of smaller follicles than in larger follicles (Fig. 4). Staining was present in adrenal cortical tissue in some 3-, 6-, and 9-week-old chickens but not in the one gland examined from a 0-week-old chick. Rarely, specific staining in pancreatic islets was present in one 6-week-old and one 9-week-old chicken, but most pancreatic sections collected did not contain islets. Thyroid glands, adrenal glands, and pancreatic tissues were not identified in embryos and therefore were not examined.
Adenohypophysis; 9-week-old chicken. Viral nucleic acid is stained in numerous epithelial cells. ISH with ALV-J antisense riboprobe, Mayer's hematoxylin counterstain. Bar = 20 µm.
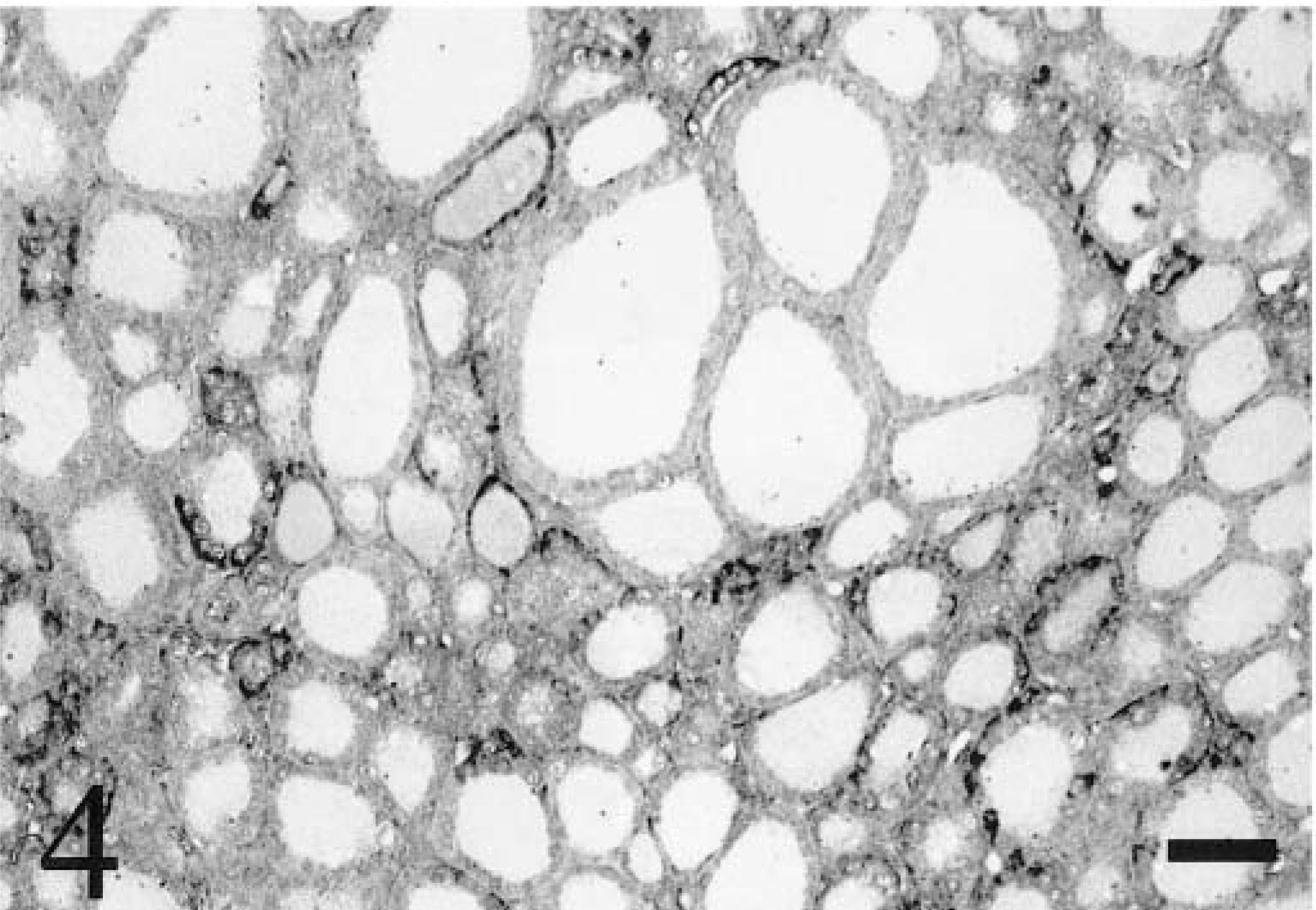
Thyroid gland; 9-week-old chicken. Viral nucleic acid is stained in the epithelium of many follicles. Follicles exhibiting positive ALV-J staining are smaller than those without positive staining. ISH with ALV-J antisense riboprobe, Mayer's hematoxylin counterstain. Bar = 50 µm.
Lymphoid tissues
Staining was present within the cloacal bursa of 0-, 3-, 6-, and 9-week-old chickens, but thymic staining was only present in 0-week-old chicks. In both bursa and thymus, staining occurred in cells morphologically consistent with dendritic cells. Bursal staining was present within follicular medullae but not follicular cortices (Fig. 5). In 3-, 6-, and 9-week-old chickens, medullary staining was not as frequent or intense as in 0-week-old chicks. Rarely, bursal epithelial staining was also present in 0-, 3-, 6-, and 9-week-old chickens. The bursa was not identified in embryos and therefore was not examined. Infrequent thymic medullary cells had staining in two of four 0-week-old chicks. One 6-week-old chicken had staining within dendritic cells of the spleen. One 9-week-old chicken had staining within lymphoid follicular medullae within the spleen.
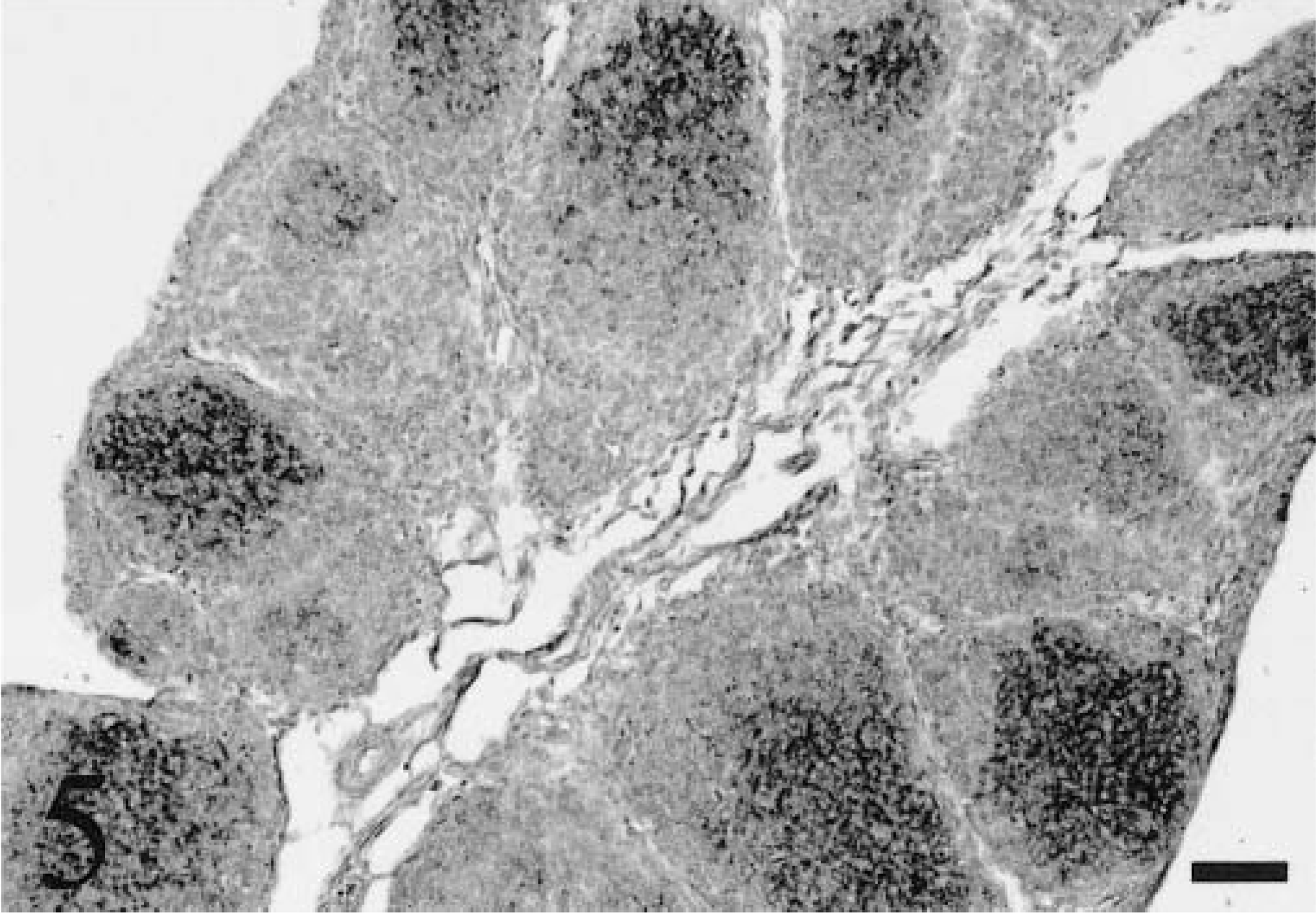
Cloacal bursa; 0-week-old chicken. Viral nucleic acid is stained in the medullae of numerous follicles. ISH with ALV-J antisense riboprobe, Mayer's hematoxylin counterstain. Bar = 50 µm.
Gastrointestinal tissues
Infrequent staining was present in the proventricular muscularis or glandular epithelium in all 0-week-old chicks and in the duodenal muscularis in most 0-week-old chicks. Abundant specific staining was also present in the esophageal muscularis in all 0-week-old chicks. Proventricular and duodenal muscularis staining was not present in 3-week-old chickens, but some staining was present in 6- and 9-week-old chickens. Staining was not present in the gastrointestinal tract of embryos.
Other tissues
Ovary from one 3-week-old chicken had multifocal interstitial staining. Staining was not present in testis, liver, bone marrow, peripheral nerve, skeletal muscle, or yolk sac at any age or in the fibrosarcoma that developed in one 9-week-old chicken. Staining was not present in any tissues collected from ALV-negative embryos or chickens at any age.
Discussion
The SU domain of the env gene of ALV-J determines subgroup specificity, and nucleic acid probes synthesized from this region are specific for ALV-J. 3,30 Nucleic acid probes failed to detect ALV-J proviral DNA but could detect viral transcripts in tissues from experimentally infected chickens. 3 Therefore, we used a riboprobe specific for the same region of the SU domain to detect viral transcripts in chickens naturally infected in ovo. Our results agree with those previously published for chickens experimentally inoculated with the prototype ALV-J strain HPRS-103 and an acutely transforming derivative virus, strain 966, with some exceptions.
We confirmed the presence of ALV-J–specific transcripts in cardiac myofibers as previously reported. 3 Transcripts also were detected in Purkinje fibers and vascular smooth muscle, and intracytoplasmic inclusions were noted in both cardiac myofibers and Purkinje fibers in HE-stained sections from 3-, 6-, and 9-week-old infected chickens. Similar inclusions have been reported in myocardium, ventricular smooth muscle, arterioles, feather follicles, and skeletal muscle of adult chickens naturally infected with lymphoid leukosis virus, and ultrastructural examination of these inclusions revealed ribonucleic acid and protein. 17,18 Immunogold labeling of inclusions within myocardium and Purkinje fibers revealed ALV group-specific p27 protein. 33 This is the first documentation of viral inclusions associated with ALV-J infection and the first demonstration of subgroup-specific nucleic acid within these inclusions in myocardium, Purkinje fibers, renal glomeruli, and pituitary glands. Cardiomyopathy with viral inclusions has been associated with the development of ascites in other ALV infections, and initial ALV-J isolates came from flocks with a high prevalence of ascites and retroviral particles in the myocardium. 16,17,20,24 One clinical manifestation of ALV-J infection may therefore be dilated cardiomyopathy and congestive heart failure. Inclusions were evident in HE-stained sections from older chickens but not in those from newly hatched chicks. Mature nonreplicating myofibers are thought to support abortive replication of lymphoid leukosis virus and to accumulate RNA and viral proteins, forming inclusions. 15
We confirmed previous findings of ALV-J transcripts within the renal distal tubules but additionally report transcripts in glomerular visceral epithelium and, rarely, mesangium. Staining patterns in the tubules of these chickens are consistent with the phenomenon of retroviral budding from the basolateral, but not apical, portions of polarized epithelial cells. 22 Virion production at this site would contribute to viremia but not tubular lumenal excretion of virions. Productive infection of glomerular visceral epithelium provides a mechanism for urinary excretion of virions and implicates urinary shedding as a route for horizontal transmission of the virus. Basophilic inclusions were noted in visceral epithelium in 3-, 6-, and 9-week-old chickens but not in newly hatched chicks, which is suggestive of some degree of abortive production in these cells in older chickens, similar to the proposed mechanism in cardiac myocytes.
ALV-J transcripts were detected in thyroid, adrenal, and pituitary glands. In contrast to previous findings, few adrenal glands in these chickens had detectable transcripts, and transcripts were restricted to cortical tissue, as previously reported. The presence of ALV-J transcripts in thyroid and pituitary glands is a novel finding and is consistent with previous findings for infection with ALV-A. 13 The significance of this finding is unknown because chickens congenitally infected with ALV-A did not exhibit any growth suppression despite widespread evidence of viral replication in multiple endocrine organs. Other ALVs have been associated with stunting, lymphocytic thyroiditis, and lowered thyroxine levels. 7,8 All 3-, 6-, and 9-week-old ALV-J–infected chickens used in this study exhibited severe stunting consistent with previously published observations, but none had lymphocytic thyroiditis. 32 The effects of ALV-J on endocrine function are unknown. The smaller follicle size associated with ALV-J transcripts in follicular epithelial cells implies reduced colloid production, which may be related to growth suppression.
The ability of ALV-J to transform myeloid precursor cells indicates infection of these precursors, but no transcripts were detected in hematopoietic or other tissues in bone marrow or yolk sac. 25 Hematopoietic tissue was also present in the spleens of 0-week-old chickens but contained no detectable transcripts. ALV-J transcripts were present in medullary dendritic cells of the thymus and bursal follicles and in splenic dendritic cells. A similar finding of minimal viral transcription in bone marrow precursors but frequent transcription in tissue macrophages and macrophage-derived cells has been reported for a macrophage-tropic ovine lentivirus. 14 Those authors speculated that viral RNA accumulated in tissue macrophages because of inefficient processing into infectious particles, similar to the mechanism proposed for the formation of inclusions in ALV-infected cardiac myocytes. In the present study, ALV-J infection may have produced differential staining of macrophage precursors and tissue macrophages by a similar mechanism. The frequent staining in bursal follicular medullae may also reflect bursal epithelial uptake of viral particles excreted in urine and feces and subsequent passing of particles into the follicular medullae. Severe immunosuppression associated with the presence of ALV-B in dendritic cells has been documented. 12,28 We are currently investigating the effects of ALV-J on immune function in broiler chickens.
No temporal trends in either transcripts or protein expression in ALV-J infection have been reported previously, but only two age groups of chickens have been examined. In the present study, some temporal trends in ALV-J transcription were detected, although the same tissues were consistently infected at all ages. Detectable transcripts were minimal in embryos, abundant in newly hatched chicks, and variable in older chickens. Because ISH was performed on the tissues from each age group in multiple batches with consistent results, these findings are likely real and not a laboratory artifact. No single explanation seems plausible for these temporal trends in ALV-J transcription. The receptor for ALV-J has not been described, but it may be minimally expressed in embryonic tissues and most frequently expressed in newly hatched chicks. Death of ALV-J–infected cells after hatching would also result in decreased detectable viral transcripts within tissues from older chicks. Because inflammation was not present in any tissues examined in this study, apoptosis of infected cells would be one possible mechanism for this decrease.
ALV-J has the strongest tropism for cardiac myocytes, Purkinje fibers, vascular and pulmonary smooth muscle, renal glomeruli, distal tubules, and pituitary glands in broiler chickens naturally infected in ovo and may form intracytoplasmic viral inclusions in infected cardiac, renal, and pituitary tissue. ALV-J is also present in gastrointestinal smooth muscle, thyroid and adrenal glands, and follicular medullae in the cloacal bursa. Viral nucleic acid was not detected in bone marrow by RNA ISH, although myeloid precursors are frequently transformed during ALV-J infection. The strong tropism of ALV-J for cardiac and endocrine tissue may relate pathogenetically to the ascites and body weight suppression noted in naturally infected broiler chickens.
Footnotes
Acknowledgements
This work was supported by a grant from the US Poultry and Egg Association. We thank J. Stanton, M. Scott, D. Hilt, and M. Jackwood for their technical assistance.