
Abstract
Serofibrinous pleuropneumonia in a piglet was examined microbiologically and immunohistopathologically. Large numbers of Morganella morganii were isolated from the pneumonic lesion, but no other pathogens were identified. A large amount of M. morganii antigen was demonstrated, and its distribution was closely associated with the histologic lesion. This finding suggests that pleuropneumonia in piglets might be caused by M. morganii.
Keywords
Morganella morganii is considered part of the tribe Proteeae within the family Enterobacteriaceae. M. morganii was originally isolated from human infants with diarrhea 7 and rarely causes lower respiratory tract infection. 6 In 1981, patients in a cardiac surgery unit developed septicemia caused by M. morganii. 12 Following this incident, M. morganii has been identified as a significant cause of nosocomial infection. However, there have been no reports of disease caused by M. morganii in domestic animals and birds, but disease has been reported in reptiles 2,4 and seals. 11 This is the first report of a sporadic case of pleuropneumonia in piglet associated with M. morganii, which was identified by immunohistopathology.
Five clinically healthy 10-day-old pigs were transferred to pathogen-free isolation facilities from a conventional farm on 18 March 1998. The conventional farm kept a herd known to be infected with Actinobacillus pleuropneumoniae serotype 2, Haemophilus parasuis, Mycoplasma hyopneumoniae, Pasteurella multocida capsular type A, porcine reproductive and respiratory syndrome virus (PRRSV), and Aujeszky's disease virus. Pigs were moved to a sterilized iron box in which a cloth was spread. Pigs were transported to an isolation facility with open pens that were ventilated by positive pressure fans that supplied filtered air. Strict sanitation procedures were followed. Pigs ate an unmedicated sterile diet three times daily in pens kept at a controlled temperature. One day after moving the animals, one pig showed anorexia and dullness. Three days later, the pig became recumbent and prostrate. The pig died 7 days after arrival at the facility. No antibiotic treatment was administered, and no clinical signs were observed in other animals.
Necropsy was done within 10 minutes of death. The right lung contained diffuse dark red consolidated areas with focal fibrinous adhesive pleuritis involving all lobes, and the spleen was slightly brown. There were no macroscopic lesions in other organs or tissues, including the left lung lobes.
A fresh sample of the right lung was stamped onto blood agar and incubated under aerobic and anaerobic conditions. A heavy growth of gram-negative rods was observed in the samples under both conditions. Other bacteria were not isolated. The organism was identified as M. morganii by two biochemical kits for the identification of bacteria (Bio test 1go, Eiken Kagaku, Tokyo, Japan; Enteogram, Waco, Osaka, Japan). The strain was sent to Dr. Kazumichi Tamura (Department of Bacteriology, National Institute of Infectious Diseases, Japan), who confirmed its identity as M. morganii (Table 1).
Characteristics of isolated bacteria from right lung of a piglet with pleuropneumonia.

Virus isolation was also performed according to established techniques using porcine kidkey (CPK) cells, hamster lung (HmLu-1) cells, and porcine alveolar macrophages. PRRSV was isolated from the spleen, serum at the time of displacement, and serum at necropsy using porcine alveolar macrophage cultures. Viral RNA of PRRSV was detected from the lung by polymerase chain reaction (PCR) assay as previously described. 5
After systematic necropsy, tissues were fixed in 20% neutral buffered formalin, embedded in paraffin, sectioned, and stained with hematoxylin and eosin (HE). Additional sections of liver, spleen, kidney, heart, lung, and jejunum were stained with Berlin blue stain, and the MacCallum–Goodpasture method for gram-positive and gram-negative bacteria. Immunohistochemically, bacterial and viral antigens were demonstrated by Envision+ Rabbit/HRP (Dako, Copenhagen, Denmark). After being deparaffinized, sections were rehydrated, immersed in 3% hydrogen peroxide for 30 minutes to block any endogenous peroxidase activity, treated with normal goat serum for 20 minutes, and then incubated with primary antibodies at room temperature for 30 minutes. Anti-M. morganii, anti-A. pleuropneumoniae serotypes 1, 2, and 5, anti-P. multocida capsular type A, and anti-PRRSV rabbit sera were used as primary antibodies at dilutions of 1:8,000, 1:4,000, 1:4,000, 1:4,000, 1:2,000, and 1:2,000, respectively. Sections were then treated with peroxidase-labeled polymeric conjugate for 30 minutes. 3,3′-Diaminobenzidine tetrahydrochloride served as the chromogen, and the sections were counterstained with methyl green. Antiserum to M. morganii was prepared by injecting a New Zealand white rabbit intramuscularly twice at 2-week intervals with 2 ml of whole, formalin-killed isolate from the lung of the affected piglet. The antigen had been emulsified with an equal volume of Freund's incomplete adjuvant. Two weeks after the second injection, the rabbit was inoculated intravenously with two increasing doses (2 and 3 ml) of antigen alone at 2-week intervals. The serum was collected 4 days after the final inoculation. For the specificity test, the antiserum was tested with the isolate by somatic agglutination, and the dilution was 1:1,280. The antiserum did not cross-react with A. pleuropneumoniae serotypes 1, 2, and 5, Bordetella bronchiseptica, Escherichia coli, H. parasuis, M. hyopneumoniae, P. multocida capsular type A, Salmonella cholerae-suis, or Streptococcus suis type II in pneumonia lesions. Other antisera were used as primary antibody and were prepared in a similar manner. The specificity of each primary antibody was assured by detection of the pathogen from lung tissue of pigs with experimental or natural infection. Tissue sections from noninfected gnotobiotic pigs and preimmune serum from the immunized rabbit were used for negative control purposes.
Histologic examination of the right lung revealed severe serofibrinous pleuropneumonia (Fig. 1). There was severe parenchymal coagulative necrosis circumscribed by a layer of degenerating inflammatory cells, moderate macrophage and neutrophil exudation into alveoli, severe capillary congestion and hemorrhage, severe inflammatory vasculitis with thrombosis (Fig. 2), moderate hemosiderin accumulation in alveolar macrophages, marked pulmonary pleura and interlobular septum thickening with fibrin and inflammatory cells, and lymphatic dilatation by edema. Hemosiderin accumulation was observed in the liver (Kupffer's cells and Glisson's sheath: severe), spleen (red pulp: severe), and kidney (mesangial cells: moderate). Hemosiderin was identified by Berlin blue stain. Minimal lymphocyte and plasma cell infiltration was found in the interstitium of the myocardium. Renal tubules were slightly dilated. Numerous gram-negative rods were identified in the pleura, interlobular septum, and alveoli of the affected lung.
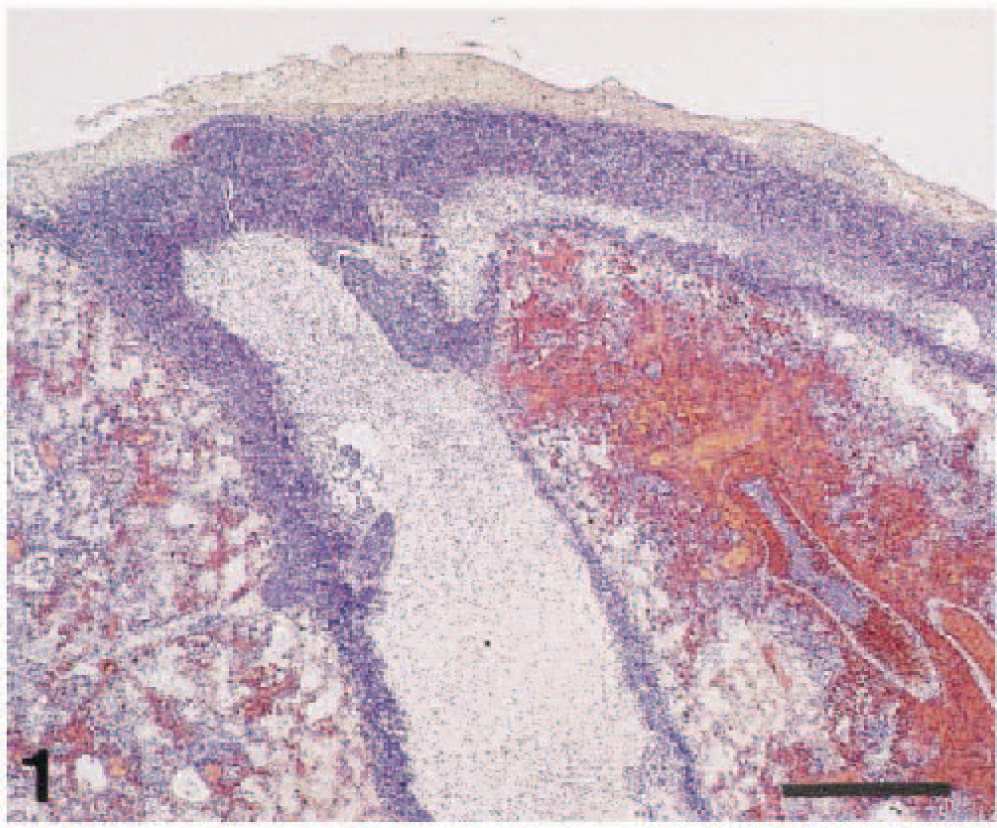
Right lung, intermediate lobes; piglet. Note severe serofibrinous pleuropneumonia with marked edematous interlobular septum, pulmonary pleura thickening, and parenchymal necrotic area with hemorrhage and congestion. HE. Bar = 500 µm.
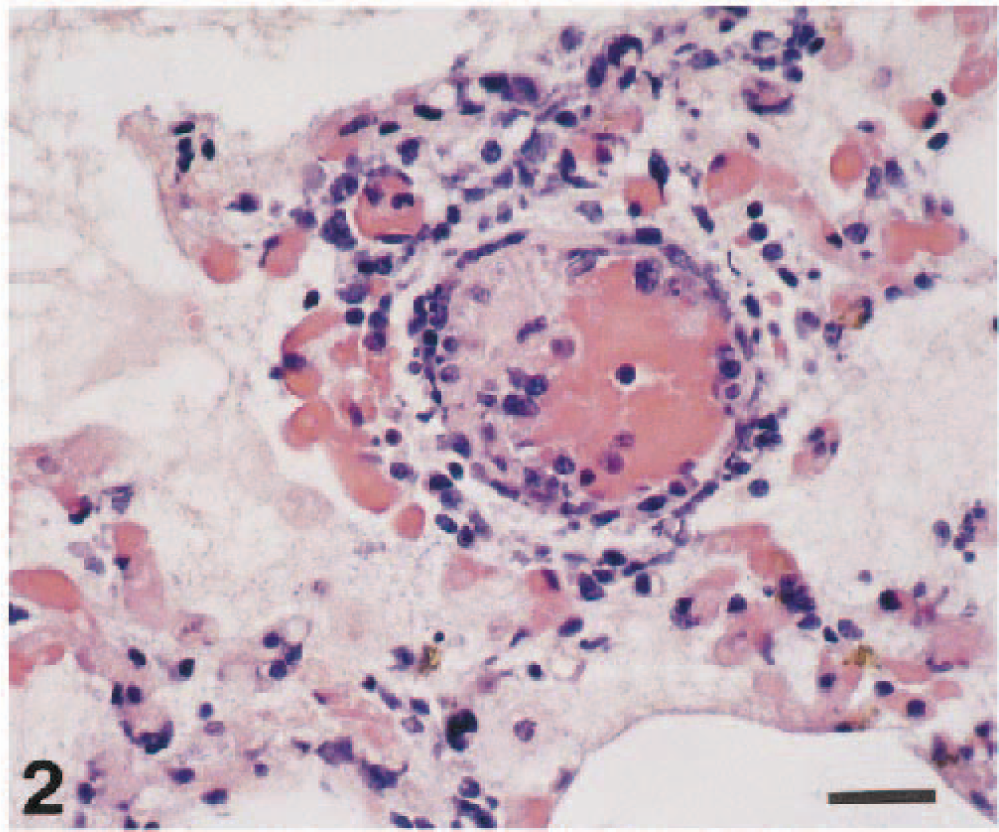
Right lung, intermediate lobes; piglet. In the pneumonic area, there is inflammatory vasculitis with fibrin thrombus, congestion, hemosiderin accumulation. Alveoli contain fibrin and macrophages. HE. Bar = 20 µm.
Immunohistochemically, M. morganii antigen was found in the necrotic lesion, alveoli, interlobular septa, pulmonary pleura (Fig. 3), and tonsillar crypt. Its distribution was closely associated with the histologic lesion. No other bacterial or viral antigen was demonstrated in the lungs.
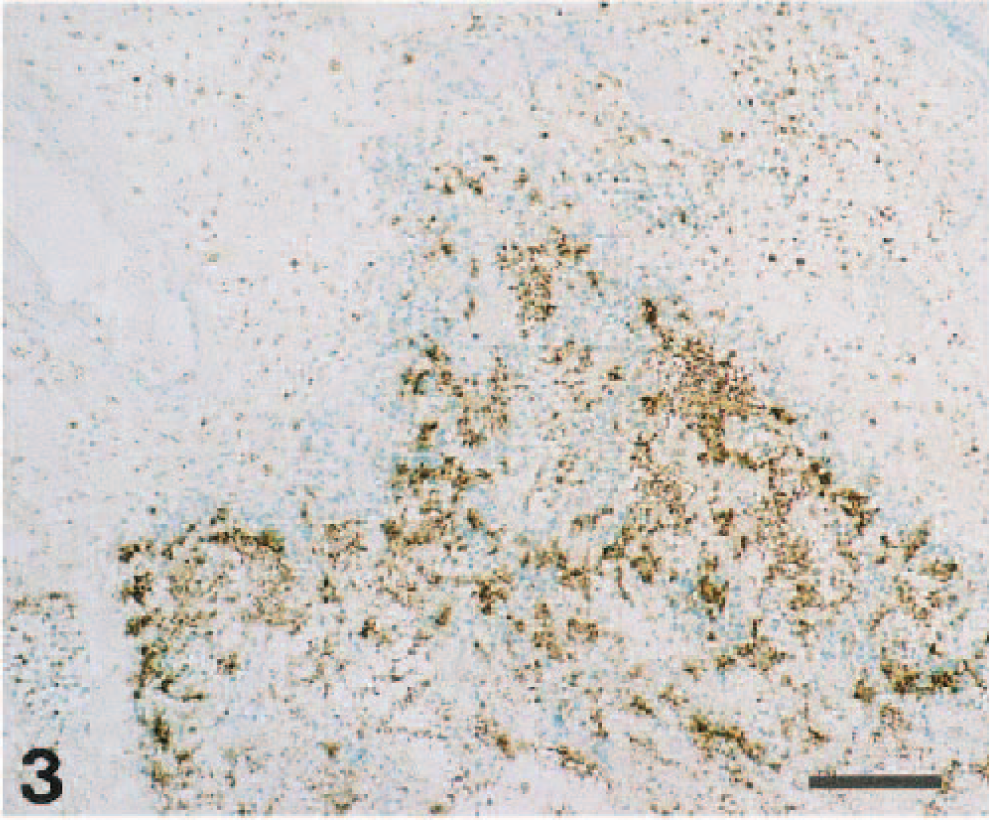
Right lung, intermediate lobes; piglet. M. morganii antigen was localized in the edematous interlobular septum, dilated lymph duct, and alveolar sac. Immunohistochemical stain (indirect method), Methyl green counterstain. Bar = 100 µm.
M. morganii has been recognized as a relatively unimportant human pathogen 6,9 and has not been reported in domestic animals. The affected piglet exhibited anorexia, dullness, and prostration and had serofibrinous pleuropneumonia. M. morganii was isolated from the lungs, but no other pathogen associated with pleuropneumonia in pigs was identified. The clinicomicrobiologic results indicate that this pleuropneumonia was induced by M. morganii infection.
The affected piglet had severe serofibrinous pleuropneumonia composed of parenchymal coagulative necrosis, hemorrhage, vasculitis, thrombosis, and interlobular edema. A large amount of M. morganii antigen was demonstrated immunohistochemically in the lungs, and its distribution was closely associated with the pleuropneumonic lesions. Immunohistopathologically, pleuropneumonia was identified as the characteristic lesion in the piglet infected with M. morganii. Therefore, the pneumonic lesion induced by M. morganii in the piglet could be differentiated from those of A. pleuropneumoniae, P. multocida capsular type A, H. parasuis, PRRSV, and Aujeszky's disease virus infections.
M. morganii is an uncommon pneumonic pathogen. Therefore, immunosuppressive factors might have played a role in the progression of this case. PRRSV infection, 10 transportation, 3 and weaning at an early age 1 have been reported as immunosuppressive factors. In the present case, the sporadic pleuropneumonia occurred in a transported piglet that was weaned when 10 days of age. Results of virus isolation and PCR indicated that this piglet was already infected with PRRSV at the time of transport. Thus, the vasculitis and lymphoplasmacytic myocarditis in this pig might have been caused by PRRSV. 8 Infection with PRRSV and change in the environment might have increased the susceptibility of this pig to M. morganii infection.
Footnotes
Acknowledgements
We thank Dr. M. Narita of the National Institute of Animal Health for his valuable suggestions and comments regarding this manuscript and Y. Ando of the National Institute of Animal Health for preparation of photomicrographs.