
Abstract
Fluorescent in situ hybridization, immunohistochemistry, and Grocott's methenamine-silver nitrate staining were compared as diagnostic methods for Pneumocystis carinii pneumonia in formalin-fixed lung tissue from foals and pigs. An oligonucleotide probe targeting 18S ribosomal RNA of P. carinii was designed for in situ hybridization, and a commercially available monoclonal antibody was used for immunohistochemistry. Samples from six foals and 10 pigs with P. carinii pneumonia, as verified by Grocott's methenaminesilver nitrate staining, were examined concurrently with samples from seven animals with pneumonia caused by other pathogens. Fluorescent in situ hybridization showed distinctive positive reactions for P. carinii in all test samples. The immunohistochemical procedure, however, only revealed P. carinii in the foals. The number of P. carinii organisms observed by fluorescent in situ hybridization and immunohistochemistry far exceeded the number of organisms stained by Grocott's methenamine-silver nitrate staining. The results show that fluorescent in situ hybridization targeting ribosomal RNA can provide a specific diagnosis of P. carinii pneumonia in foals and pigs.
Pneumocystis carinii is an opportunistic fungal pathogen that may cause fatal interstitial pneumonia in immunocompromised hosts. In domestic animals, P. carinii pneumonia (PCP) has been reported in immune-deficient or malnourished foals, 1,14,23,24,29 pigs, 9,11,18,19,25,26 goats, 21 dogs, and cats. 15,27 Although P. carinii from humans and different animals are morphologically indistinguishable, studies on genotypic and phenotypic parameters indicate that P. carinii organisms are species specific. 5,11,24,28,31
Prior to the availability of molecular techniques, the verification of the diagnosis was dependent on the histomorphologic identification of P. carinii cysts in sections or smears by special staining methods such as Grocott's methenamine-silver nitrate (GMS) or toluidine blue O. However, these methods are only indicative, directed to common fungal polysaccharide moieties of the cyst wall. Differentiation from other fungi may be a problem. In situ hybridization has successfully been used to detect P. carinii in lungs from rats and humans, using biotinylated oligonucleotide probes targeting ribosomal RNA (rRNA) 13,16 and in lungs from humans and ferrets by fluorescent in situ hybridization targeting specific small-subunit rRNA genes. 28 With the exception of detection of P. carinii in two pigs, in situ hybridization has so far not been properly evaluated as a diagnostic tool in piglets or in foals. 25
Here, we describe application of fluorescent in situ hybridization (FISH) targeting 18S rRNA of P. carinii in formalin-fixed lung samples from horses and pigs with PCP and an immunohistochemical method (IM) using a commercially available anti-human P. carinii monoclonal antibody. This antibody has previously been used immunohistochemically to demonstrate P. carinii in humans and in one foal. 29 The two methods are compared with GMS staining.
Material and Methods
Animals and histology
Pneumonic lungs from eight foals and 15 pigs submitted for laboratory examination were tested. Tissue samples from six of the foals and 10 of the pigs had previously been verified as positive for PCP by GMS. Lung samples from the two foals and five pigs with pneumonia caused by other pathogens were tested as negative controls in parallel (two foals with Rhodococcus equi, two pigs with chronic Actinobacillus pleuropneumoniae, and three pigs with Streptococcus suis). The foals were between 2 and 6 months of age, and the pigs were 1–3 months old. All foals had a history of longstanding, incurable pneumonia.
The samples were fixed in 10% neutral buffered formalin, embedded in paraffin, sectioned 5 µm, mounted on Super Frost∗/plus slides (Menzel-Gläser®, Braunschweig, Germany) and kept at 4 C until used. Sections of each sample were stained by GMS or used for IM or FISH. Sections used for micrographs were initially processed for in situ hybridization and fluorescence microscopy and later counterstained by IM or GMS.
Immunohistochemistry procedure
After deparaffinization the slides were rehydrated and washed three times for 5 minutes each in Tris-buffered saline (TBS, pH 7.6) before antigen demasking in 1 mM ethylene-diaminetetraacetic acid (pH 9.0) for 20 minutes in a microwave oven (at 850 W). Endogenous peroxidase activity was inhibited by incubation in TBS with 0.6% H2O2 (v/v) for 15 minutes.
The slides were incubated in a moist chamber with TBS with 5% (v/v) normal swine serum (TBS + NSS), followed by incubation with a mouse monoclonal antibody against human P. carinii (DAKO-M778, DAKO A/S, Glostrup, Denmark) in TBS + NSS overnight, and then washed in TBS. Incubated with biotinylated rabbit anti-mouse antibody (DAKO-E465, DAKO A/S) in TBS + NSS for 30 minutes followed by incubation with peroxidase-conjugated streptavidin (DAKO-P397, DAKO A/S) in TBS + NSS for 30 minutes. The reaction was developed for 15 minutes with diaminobenzidine (DAB) (DAKO-S3000, DAKO A/S) counterstained with Mayer's hematoxylin, and mounted with Pertex (HistoLab, Västra Frölunda, Sweden).
Oligonucleotide probes and FISH
The oligonucleotide probes used were specific for P. carinii Carinii1, 5′-GGTAATCCAGGAGGGAAGG-3′, targeting nucleotides 681–705 on the 18S rRNA. The oligonucleotide probe for P. carinii was selected from three published 18S rRNA sequences available by using the function “Probe Design” in the software ARB (this database is freely available at the Internet at http://www.biolchemie.tu-muenchen.de). The three 18S rRNA sequences all were derived from rats. No sequence from pigs, foals, or humans were available. The general eukaryotic probe EUK516, 5′-ACCAGACTTGCCCTCC-3′, targeting nucleotides 548–563 of the 18S rRNA of Saccharomyces cervisiae, a sequence that is identical in all eukaryotic organisms, 2 was used as a positive control. The nonspecific probe NON-EUB338 was used as a negative control for nonspecific binding of probe to tissue. 2 The probes were labeled with either fluorescein or CY3 (Biological Detection Systems, Pittsburg, PA). The labeled probes were purchased from Hobolth DNA Syntese (Hiller⊘d, Denmark).
The specificity of the probes was tested against all available sequences in GenBank using the BLAST program 7 and against the Ribosomal Database Project database using the program CHECK_PROBE. 20 Prior to hybridization, the sections were deparaffinated in xylene and transferred to 99% ethanol for 10 minutes. Before the hybridization solution was applied, the sections were circumscribed with a hydrophobic PAP-pen (Daido Sangyo Co., Tokyo, Japan). The hybridization was carried out at 45 C with 30 µl of hybridization buffer (100 mM Tris pH 7.2, 0.9 M NaCl, 0.1% sodium dodecyl sulfate) and 75 ng of probe for 16 hours in a moisture chamber. The samples were washed in 100 ml pre-warmed (45 C) hybridizaiton buffer for 15 minutes and then in 100 ml of prewarmed (45 C) washing solution (100 mM Tris pH 7.2, 0.9 M NaCl) for 15 minutes. The samples were rinsed in water, air-dried, and mounted in Vectashield (Vector Laboratories, Burlingame, CA) for microscopy.
Epifluorescence microscopy
An Axioplan2 epifluorescence microscope (Carl Zeiss, Oberkochen, Germany) equipped for epifluorescence with a 75-W Xenon lamp and filter sets XF23 and XF34 (Omega Optical, Brattleboro, VT) was used to visualize fluorescein and CY3, respectively. Filter set XF52 was used to visualize red and green fluorescence simultaneously. Fluorescence micrographs were taken with a Zeiss MC200 camera using Kodak™ Ektachrome 400 film.
Results
Pathology
The lungs of the P. carinii-infected foals appeared enlarged, diffusely consolidated, and hepatoid. Histologically, the alterations were characterized by subchronic to chronic interstitial pneumonia and alveolar floating, with macrophages, single giant cells, fibrinoid eosinophilic material, and hyperplasia of type II pneumocytes. Foaming acidophilic “honeycomb” material was absent.
The pig lungs appeared enlarged, firm, and meaty. The histologic changes were characterized by an interstitial pneumonia and slight thickening of septa by macrophages, monocytes, and lymphocytes. In the alveolar lumen, a proteinous exudate mixed with macrophages and desquamated epithelial cells was observed. A foamy, acidophilic “honeycomb” material distending the alveolar space was found in the lung sections of five pigs.
P. carinii detection
FISH with the P. carinii-specific oligonucleotide probe Carinii1 showed distinctive positive reactions in all samples from the test animals. No reactions were observed in the samples of the negative control lungs. The results of FISH, (IM, and GMS staining) in foals and pigs are summarized in Table 1.
Comparison of Grocott's methenamine-silver nitrate staining (GMS), immunohistochemistry (IM), and fluorescent in situ hybridization (FISH) for the detection of P. carinii in foals and pigs with pneumonia.
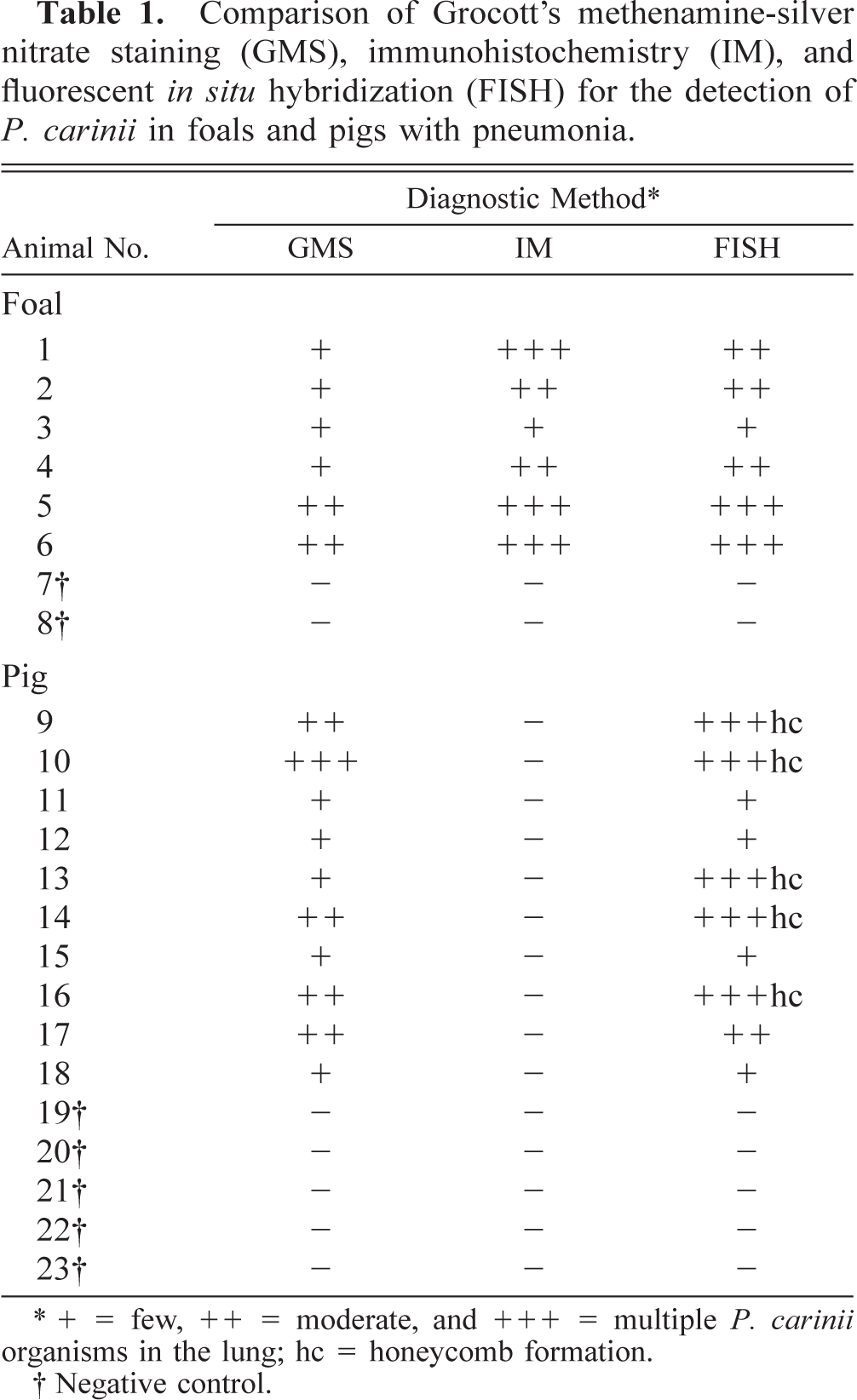
+ = few, ++ = moderate, and +++ = multiple P. carinii organisms in the lung; hc = honeycomb formation.
Negative control.
In the pig lungs, trophozoites appeared by FISH as homogeneously orange to red, oval to spherical bodies of 2–5 µm lining the alveolar epithelium or in the alveolar exudate. Cysts, confirmed by an argyrophilic staining of the cyst wall in GMS, similarly appeared oval to spherical and 4–6 µm. The detection of P. carinii in the same section initially by FISH and then after counterstaining with GMS is shown in Fig. 1. Some cysts were homogeneous orange to red, whereas others contained four or five distinctive intracystic bodies. In addition, empty crescent-shaped cysts were seen in GMS-stained sections. In cases of honeycomb formation, large clusters of P. carinii organisms appeared by FISH like bunches of bright orange balls with an almost fluorescent translucent central area (Fig. 2). The FISH sections revealed more P. carinii organisms than did GMS staining, especially in areas with the honeycomb formation. Application of the IM procedure in the pig lungs failed to give positive reactions.
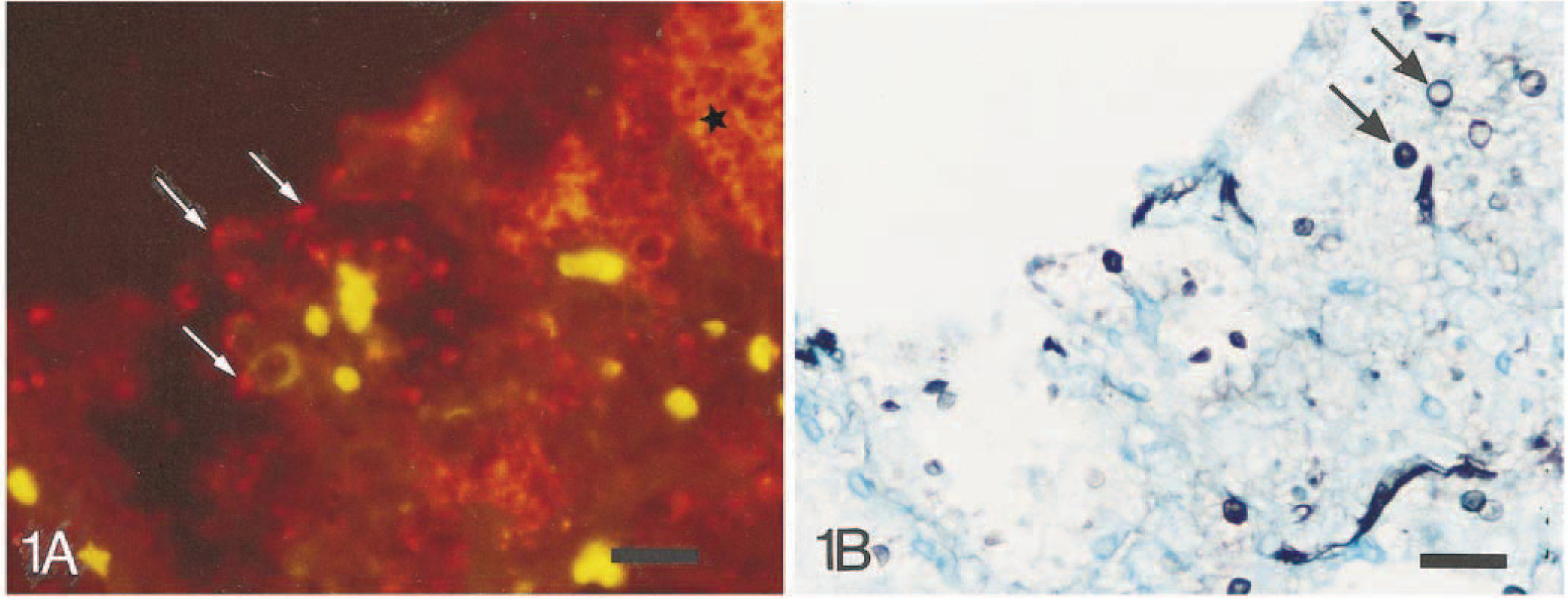
Lung; pig. Identification of P. carinii. Fig. 1A FISH using the CY3-labeled specific oligonucleotide probe Carinii1 and red/green filterset. P. carinii organisms are orange to red bodies (arrows). Note honeycomb formation (∗). Erythrocytes appear as bright yellow spots because of autofluorescence. Fig. 1B Same section counterstained by Grocott's methenamine-silver nitrate (GMS) only revealing cysts (arrows). Bar = 20 µm.
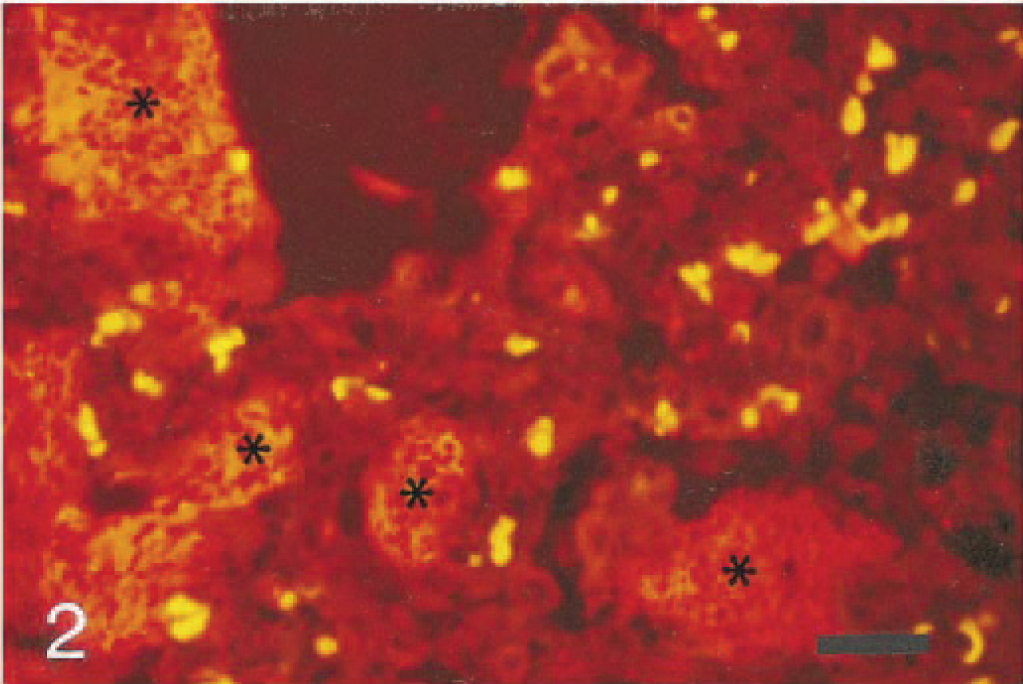
Lung; pig. Large clusters of bright orange P. carinii organisms (∗) distending the alveolar lumen with extensive honeycomb formation. FISH, CY3-labeled probe and red/green filterset. Bar = 50 µm.
Most of the P. carinii organisms revealed by FISH in the foal lungs were 2–3 µm long, polymorphic to oval, homogeneously orange to red trophozoites; the number of cysts was low, as verified by GMS. Figures 3 and 4 show the detection of P. carinii by FISH and subsequent counterstaining by IM on the same section from a foal. A parallel section of the lung stained by GMS revealed only single cysts (Fig. 5). Occasionally, cysts containing three to five distinctive intracystic bodies were observed. In most of the foals, the intensity of the fluorescent signal was decreased as compared with the signal obtained in the pig lungs. The IM sections revealed P. carinii as brown, 2–5-µm-long, single surface–stained polymorphic organisms that often tended to cluster together, partly lining the alveolar epithelium (Fig 3B). In addition, smaller brown fragments were seen scattered in the alveolar exudate. Compared with FISH, the number of positive reactions by IM was generally higher (Table 1). The number of P. carinii cysts in the GMS-stained sections was lower than that obtained with both IM and FISH. In sections of foal lungs with a small number of P. carinii organisms, a positive diagnosis was more easily obtained by IM and by GMS than by FISH; fluorescent microscopy required a higher magnification for examination (40× or 63× versus 20×) because of decreased fluorescent signal. Furthermore, because of intensive autofluorescence in both monofilters in the lung sections from both animal species, the hybridization signals were observed most distinctively using the R/G filterset.
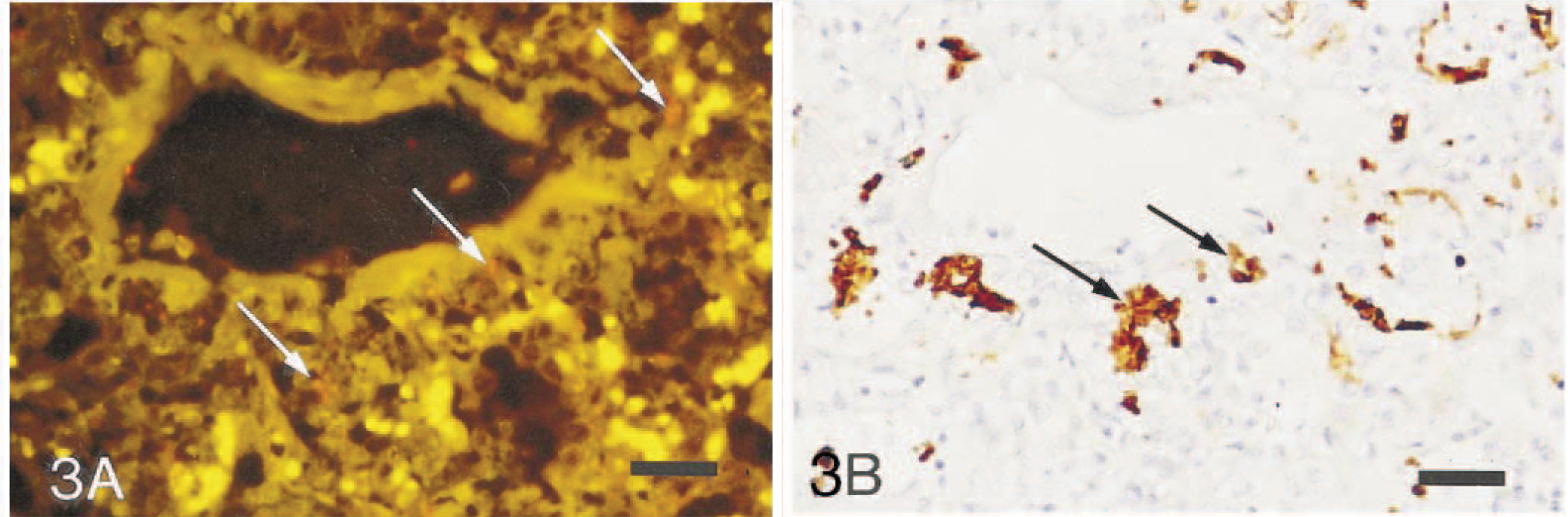
Lung; foal. Identification of P. carinii. Fig. 3A FISH using the CY3-labeled specific oligonucleotide probe Carinii1. The organisms appear orange (arrows). Erythrocytes appear as bright yellow spots because of autofluorescence (red/green filterset). Fig. 3B Same section counterstained by immunohistochemistry using a monoclonal antibody against human-derived P. carinii. The organisms are densely brown bodies (arrows). Bar = 40 µm.
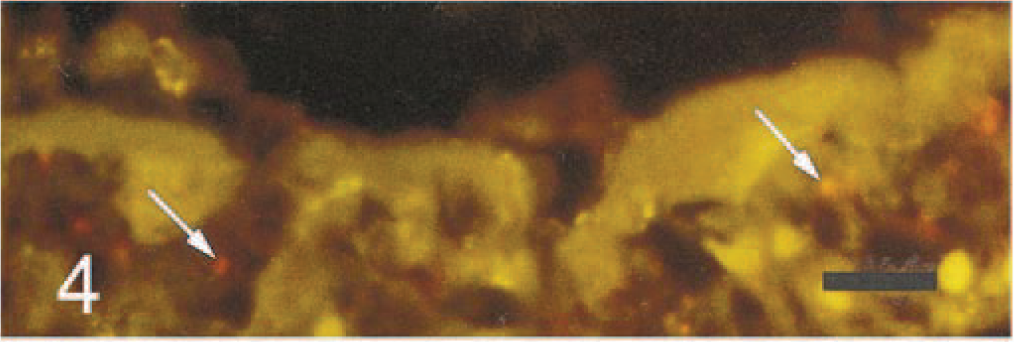
Lung; foal. Higher magnification of section in Fig. 3A showing P. carinii (arrows) in a proteinous exudate lining the alveolar lumen. Bar = 15 µm.

Lung; foal. Demonstration of P. carinii cysts by GMS in a section parallel to that shown in Fig. 4. Only a single cyst is stained (arrow). Bar = 15 µm.
Discussion
Pneumocystis carinii is an opportunistic, ubiquitous fungal pathogen that may cause fatal pneumonia. In domestic animals most PCP cases have been reported in foals and pigs. 1,9,11,14,18,19,23–26,29 Diagnosis of PCP is obtained by histomorphology and identification of the organism, which has not been successfully cultured. The traditional method for recognizing P. carinii is by staining of the cyst walls with GMS or toluidine blue O. 6 However, such methods are only suggestive because they are directed to common fungal polysaccharide moieties of the cyst wall and leave the trophozoites unstained. Thus, specific diagnostic tools for identification are essential.
The results of this study show that in situ hybridizaiton with the designed oligonucleotide probe Carinii1 provides distinctive positive identification of P. carinii in equine and porcine lungs. The present IM using a commercial available anti-human P. carinii monoclonal antibody is, however, only able to reveal the organism in the foals. The proper FISH procedure includes only three steps and is suited for routine diagnostics as a simple technique with visualization of all stages of P. carinii organisms. In situ hybridization has previously been used to detect P. carinii in lung tissue from humans and rats with probes targeting 5S and 18S rRNA. 13,16 Previously, an in situ hybridization procedure targeting 5S rRNA 22 was used on three P. carinii-infected pigs, but a positive diagnosis was made in only two of the animals. All of these procedures, however, are based on the use of biotinylated probes and visualization by a multistep immunohistochemical process, e.g., the avidin–biotin–peroxidase method. To augment the hybridization signal in human tissue, a mixture of three different probes has been used. 16 Because no sequences for P. carinii isolated from pigs and foals were available for this study, the probe Carinii1 was selected to target P. carinii derived from a rat. Similarly, probes used by others 13,16 were designed on the basis of sequences from rat-derived P. carinii. The probe was first labeled with the fluorochrome fluorescein. However, autofluorescence from the lung tissue itself seemed to be less significant when using a CY3-labeled probe and a red/green filter set.
FISH of whole cells with rRNA-targeted oligonucleotide probes has become a highly valuable tool for the specific detection of microorganisms without cultivation. 3,12 Nonculturable bacteria have been identified in their natural environment, e.g., as endosymbionts, in intestinal samples from pigs and in activated sludge. 4,10,30 RNA is naturally amplified in growing cells. An exponentially growing Escherichia coli cell contains 104–105 copies of 5S, 16S, and 23S rRNA per cell. 8 This means that a considerable increase in sensitivity is achieved by targeting rRNA instead of DNA. By probing intact microbial cells, in contrast to nucleic acid, histologic recognition and information about the morphology of the microorganisms can be obtained. In contrast to immunologic methods that rely on the expression of specific markers, which may not be constant, phenotypic variation does not pose a problem when rRNA is used as a target.
Although P. carinii isolates appear to be species specific and antigenically different, 5,11,24,28,31 the same commercial monoclonal anti-human P. carinii antibody used in the present study was used previously for the detection of PCP in one foal. 27 Thus, with the detection of multiple organisms in the six lungs, the results of the present study confirm the applicability of IM with the anti-human P. carinii monoclonal antibody for the detection of PCP in foals. However, P. carinii was not identified in the porcine lungs by IM. Similarly, this antibody has failed to detect P. carinii in rats. 16,17 IM seems to reveal all development stages of P. carinii in foals, and in comparison with FISH the number of positive reactions is higher. The difference between the two tests may reflect the microbial activity of the organisms (the foals probably had been treated for pneumonia with several antibiotics that might have decreased the activity of the organisms). The sensitivity of FISH is correlated with the amount of rRNA in the organisms at the time of fixation, whereas IM detects surface antigens. All the PCP foals had a history of longstanding, incurable pneumonia and treatment with several kinds of antibiotics. The histomorphology confirmed the chronicity of the lung lesions with pronounced cellular inflammatory response. This chronicity and the small number of organisms revealed by FISH indicate that the growth of P. carinii at the time of death in these foals was more or less suppressed.
Application of FISH with the probe Carinii1, targeting the 18S rRNA of P. carinii, showed distinctive positive reactions in foal and pig lungs that were confirmed as positive for P. carinii by GMS. IM using an anti-human P. carinii monoclonal antibody only identified P. carinii in the foals. The number of P. carinii organisms in the FISH- and IM-positive animals was higher than that obtained with GMS staining. FISH targeting 18S rRNA can provide a specific diagnosis of P. carinii pneumonia in both foals and pigs.
Footnotes
Acknowledgements
We thank A. R. Pedersen and U. L. Andreasen for excellent technical assistance.