
Abstract
Detection and localization of porcine epidemic diarrhea virus (PEDV) was studied by in situ hybridization with a nonradioactive digoxigenin-labeled probe in formalin-fixed, paraffin-embedded tissues from 10 naturally infected piglets. A 377-base pair cDNA probe for viral RNA encoding the membrane proteins of PEDV cell-culture-adapted strain V215/78 was generated by the reverse transcription polymerase chain reaction. In the retrospective study of pigs from herds with diarrhea, the 10 piglets naturally infected with PEDV had positive signals for PEDV by in situ hybridization. When intestinal tissues were hybridized with the PEDV probe, a strong signal was seen in the villus enterocytes of jejunum and ileum but not in the cecum and colon. Positive cells typically had dark brown reaction products in the cytoplasm. Scattered epithelial cells along the ileal Peyer's patches dome areas contained viral RNA. In one piglet, hybridization signal was also found in the duodenum. PEDV was not demonstrated in tissues outside of the intestinal tract. These findings indicate that jejunal and ileal villus enterocytes are the main target of PEDV replication during epizootic outbreaks of the disease.
Porcine epidemic diarrhea virus (PEDV) is a member of the family Coronaviridae. Other porcine viruses in this family include transmissible gastroenteritis virus (TGEV), porcine respiratory coronavirus, and hemagglutinating encephalomyelitis virus.1,24,26,31 PEDV was first reported in Belgium and the United Kingdom in 1978.25,31 Since then, several outbreaks of the disease have been reported in many swine-raising countries, most notably in Europe and Korea.8,19,22
PEDV causes acute enteritis in swine of all ages, and it is often fatal for neonatal piglets. The disease can appear similar to that caused by TGEV.12,31 Although these two viral diseases can be difficult to distinguish clinically, PEDV differs antigenically from TGEV, and antigenic cross-reaction has been described only with feline infectious peritonitis.32 Like TGEV, PEDV causes destruction of villus enterocytes and villus atrophy within the jejunum and ileum.9,10,13,27 Clinical signs of PEDV include anorexia, vomiting, diarrhea, and dehydration. Morbidity and mortality in infected neonatal piglets less than 5 days old approach 100% because of severe diarrhea and dehydration. However, mortality in infected piglets older than 10 days is less than 10%.5,19,25,31
PEDV was initially isolated in 1988 with the Vero cell line and a trypsin-supplemented medium.18 The porcine epidemic diarrhea (PED) virion, projection included, is approximately 130 nm in diameter with a range of 95–190 nm. Many particles have an electron-opaque central area. The club-shaped projections measure 18–23 nm and extend radially from the membrane.5,25 Replication of the virus occurs in the host cell cytoplasm.14,27 A subgenomic 7.4-kb mRNA, co-terminal with 3′ end of the genomic RNA, is translated to form structural proteins such as S and M membrane proteins.11 The nucleotides within M gene are low homologous compared (approximately 50%) with other coronaviruses, including TGEV, but highly conserved among different PEDV strains;12 thus, a complementary DNA probe transcribed from M gene could allow detection of PEDV RNA.
Because PEDV is difficult to isolate in cell culture, different diagnostic methods have been employed for the detection of both PEDV antigen and its antibodies.2,3,7,23 PEDV antigen has been detected in jejunal and ileal villus enterocytes of swine by monoclonal and polyclonal antibody–based immunohistochemical procedures.16,21,29 In situ hybridization is a valuable adjunct to standard RNA extraction techniques for evaluating gene expression in tissues and cells. Its major advantage is the ability to determine which tissues, or cells in a mixed population, are expressing the RNA of interest. In situ hybridization techniques have been used successfully to detect viral nucleic acids with DNA probes complementary to viral RNA.6,15 One objective of this study was to detect PEDV nucleic acids in formalin-fixed, paraffin-embedded tissues with digoxigenin (DIG)-labeled, 377-base, single-stranded DNA probes complementary to a conserved region of the PEDV membrane protein gene. The second objective was to determine the location of PEDV in the porcine intestine in order to better understand the pathogenesis of naturally occurring PEDV infection.
Materials and Methods
Animals
Ten piglets (Nos. 1–10) approximately 3–10 days of age, from 10 different herds, were selected on the basis of clinical signs, characteristic lesions, and positive direct immunofluorescent antibody tests for PEDV infection but negative for TGEV and rotavirus infection. Fluorescent-conjugated polyclonal antibody for PEDV was kindly provided by Dr. M. Pensaert of the University of Ghent, Belgium. Fluorescent-conjugated polyclonal antibody for TGEV and rotavirus was provided by the National Veterinary Services Laboratory (Ames, IA). Samples were obtained from the necropsy cases of the Department of Veterinary Pathology, College of Veterinary Medicine, Seoul National University. All piglets were submitted alive and samples were collected at necropsy. Results of virus isolation from 10 piglets were published previously.20 Only one PEDV (pig No. 3) was isolated and adapted into Vero cells. The rest of the samples failed to isolate and adapt in Vero cells because PEDV is difficult to isolate in cell culture.
Negative tissue (jejunum and ileum) controls were collected from a 1-day-old colostrum-deprived pig not exposed to PEDV or TGEV and from each of 3 piglets that had been naturally infected with TGEV or rotavirus.
Tissue processing
Tissue specimens were collected from lung, tonsil, stomach, duodenum, jejunum, ileum, colon, and cecum of infected and noninfected animals, fixed in 10% (w/v) buffered formaldehyde for 24–48 hours, and embedded in paraffin according to standard laboratory procedures. Sections were cut 3 μm thick, floated on a water bath containing diethylpyrocarbonate-treated water, and mounted on Superfrost/plus slides (Fisher Scientific, Pittsburgh, PA).
PEDV-infected cell cultures for in situ hybridization
PEDV-infected and noninfected Vero cell cultures were used as the starting material for optimization of in situ hybridization. The 100 μl of tissue culture fluid containing 107 50% tissue culture infective dose/ml of a cell-culture-adapted strain V215/78 PEDV, kindly provided by Dr. M. Ackermann of the University of Zürich, was grown in Vero cells in four-well chamber slides (Nunc, Inc., Naperville, IL) as previously described.18 After 12–24 hours of incubation at 37 C, at which time the virus-specific cytopathic effect covered 95% of the monolayered cell, the medium was discarded, the wells were rinsed with phosphate-buffered saline (PBS; 0.1 M, pH 7.4), and the cells were fixed by adding PLP (4% paraformaldehyde, 100 mM
RNA extraction
The cell-culture-adapted PEDV strain V215/78 was used as the standard strain and grown in Vero cells as previously described.18 PEDV-infected Vero cells (75-cm2 flask) were harvested when 70–80% of the cells had undergone cytopathic effects. RNA was extracted from PEDV-infected cells with Trizol LS reagent (Gibco BRL, Grand Island, NY) according to the manufacturer's instructions.
Probe production
The primer from the membrane protein sequence of PEDV used in this study was previously described.22 However, reverse transcription and polymerase chain reaction (PCR) conditions were modified slightly. The sense and antisense primers were 5′-GGACACATTCTTGGTGGTCT-3′ (nucleotides 1,318–1,338) and 5′-GTTTAGACTAAATGAAGCACTTTC-3′ (nucleotides 1,665–1,688), respectively. The primer set resulted in amplified fragments of 377 base pairs.
For the first-strand cDNA synthesis, 1 μl of the PEDV RNA (5 ng/μl) was supplemented in a total reaction volume of 20 μl with 1× RT buffer (50 mM Tris-HCl, 8 mM MgCl2, 30 mM KCl, 1 mM dithiothreitol, pH 8.3), 0.5 mM (each) deoxynucleotide triphosphates (dNTPs), 2.5 μM random hexanucleotide mixture, 20 U of RNase inhibitor, and 50 U of Moloney murine leukemia virus reverse transcriptase. After incubation for 15 minutes at 42 C, the mixture was incubated for 5 minutes at 99 C to denature the products. The mixture was then chilled on ice.
The composition of the PCR mixture (150 μl) was 30 μl of cDNA (5 ng/μl), 2 μl of each primer (250 nM), 15 μl of 10× PCR buffer (10 mM Tris-HCl, 40 mM KCl, 1.5 mM MgCl2, pH 8.3), 1.2 μl of each dNTP (0.2 mM), 2.5 units of Taq polymerase in a volume of 29 μl (Bioneer Corp., Cheungwon, Korea), and 67.2 μl of distilled water. The PCR reaction for PEDV was done under the following conditions in a thermal cycler (Perkin-Elmer-Cetus, Norwalk, CT): 1 cycle of 2 minutes at 94 C, 2 minutes at 58 C, and 2 minutes at 72 C; 40 cycles of denaturation at 94 C for 1 minute, annealing at 58 C for 1 minute, and elongation at 72 C for 1 minute; and 1 cycle of 2 minutes at 94 C, 2 minutes at 58 C, and 2 minutes at 72 C.
PCR products were purified with Wizard PCR preps (Promega Biotech, Madison, WI). Purified PCR product was labeled by random priming with DIG-dUTP by means of a commercial kit (Boehringer Mannheim, Indianapolis, IN) according to the manufacturer's instructions.
In situ hybridization
Sections were dewaxed in xylene and rehydrated in PBS (pH 7.4, 0.01 M) for 5 minutes. Deproteinization was carried out in 0.2 N HCl for 20 minutes at room temperature. Tissues were then digested at 37 C for 20 minutes in 100 μg/ml proteinase K (Gibco BRL) in PBS (pH 7.4, 0.01 M) and fixed in 4% paraformaldehyde in PBS for 10 minutes. After rinsing twice with PBS, the slides were acetylated in 300 ml of 0.1 mM triethanolamine-HCl buffer (pH 8.0) to which 0.75 ml of acetic anhydride (0.25%) had been added. After 5 minutes, an additional 0.75 ml of acetic anhydride was added, and 5 minutes later the slides were rinsed in 2× saline sodium citrate (SSC) (1× SSC contains 50 mM NaCl and 15 mM sodium citrate, pH 7.0). The slides were allowed to equilibrate for 60 minutes in a standard hybridization buffer that consisted of 5× SSC with 50% deionized formamide, 10× 2% buffered blocking solution (Boehringer Mannheim), 0.1% N-lauroylsarcosine, and 0.02% sodium dodecyl sulfate. Hybridization was done overnight at 45 C. The DIG-labeled probe (0.1 ng/μl) was diluted in 300 μl of the standard hybridization buffer, heated for 10 minutes at 95 C on a heating block, and quenched on ice before being applied to the tissue sections. Approximately 50 ng of the DIG-labeled probe was added to the standard hybridization buffer (50 μl), which was then layered over the section. Fluid was held in place by a coverslip, and the edges were sealed with rubber cement. After overnight hybridization, sections were thoroughly washed twice in 4× SSC for 5 minutes at room temperature, twice in 2× SSC for 10 minutes at 37 C, twice in 0.2× SSC for 5 minutes at room temperature, and once in maleic acid buffer (100 mM maleic acid and 150 mM NaCl, pH 7.5) for 5 minutes at room temperature.
For detection of hybridization, sections were incubated with anti-DIG conjugated with alkaline phosphatase (Boehringer Mannheim) diluted 1:250 in 0.1 M Tris-HCl, pH 7.4, and 0.15 M NaCl with 1% blocking reagent (Boehringer Mannheim). After three washes in buffer, substrate consisting of nitroblue tetrazolium and 5-bromocresyl-3-indolylphosphate was layered over the sections. Color was allowed to develop for 5–8 hours in the dark, and the reaction was stopped by dipping slides briefly in tri-ethylenediaminetetraacetic acid (EDTA) buffer (10 mM Tris-HCl and 1 mM EDTA, pH 8.0). Sections were counterstained with 0.5% methyl green, and the slides were then washed with distilled water for 1 minute, allowed to dry completely, dipped into the absolute xylene, and coverslipped with Canada balsam mounting medium (Hayashi Pure Chemical Industries Ltd., Osaka, Japan).
Results
Microscopic lesions
The most consistent and predominant lesion in the diarrheic piglets was moderate to severe multifocal to diffuse villus atrophy in the distal portion of the jejunum and proximal portion of the ileum. Villi often were fused and covered with a degenerated or regenerated flattened epithelium. Rarely, villi were seen that lacked an epithelial lining. Vacuolated enterocytes were seen on the tips of villi or spread over the entire villi in the jejunum. Moderate numbers of exfoliated enterocytes were seen on scattered villi. The lamina propria had small numbers of eosinophils and neutrophils. The crypts of Lieberkühn appeared normal. The cecum and colon showed no apparent changes except for vacuolation of the superficial enterocytes. No lesions were seen in other tissues.
In situ hybridization
Results of in situ hybridization are summarized in Table 1. PEDV sequences were detected by use of an anti-DIG–alkaline phosphatase conjugate when PEDV-infected Vero cells were hybridized with a specific DIG-labeled PEDV cDNA (377 bp) probe (Fig. 1). The PEDV nucleic acid signal was specific because this was completely absent from mock-infected Vero and CRFK cells and TGEV-infected CRFK cells. The 10 piglets naturally infected with PEDV had positive signals for PEDV by in situ hybridization. A strong hybridization signal for PEDV was seen in the villus enterocytes of jejunum (Fig. 2) and ileum but not in the crypts. Although the signal intensity varied within and between anatomic structures in any one section and between piglets, jejunal villus enterocytes had stronger hybridization signals than ileal villus enterocytes (pig Nos. 5, 8, and 10). Positive cells had dark brown or black reaction product, and there was no background staining. Hybridization signals were confined to the cytoplasm (Fig. 3).
Results of in situ hybridization for porcine epidemic diarrhea virus (PEDV) in the small intestinal villi of piglets naturally infected with PEDV.
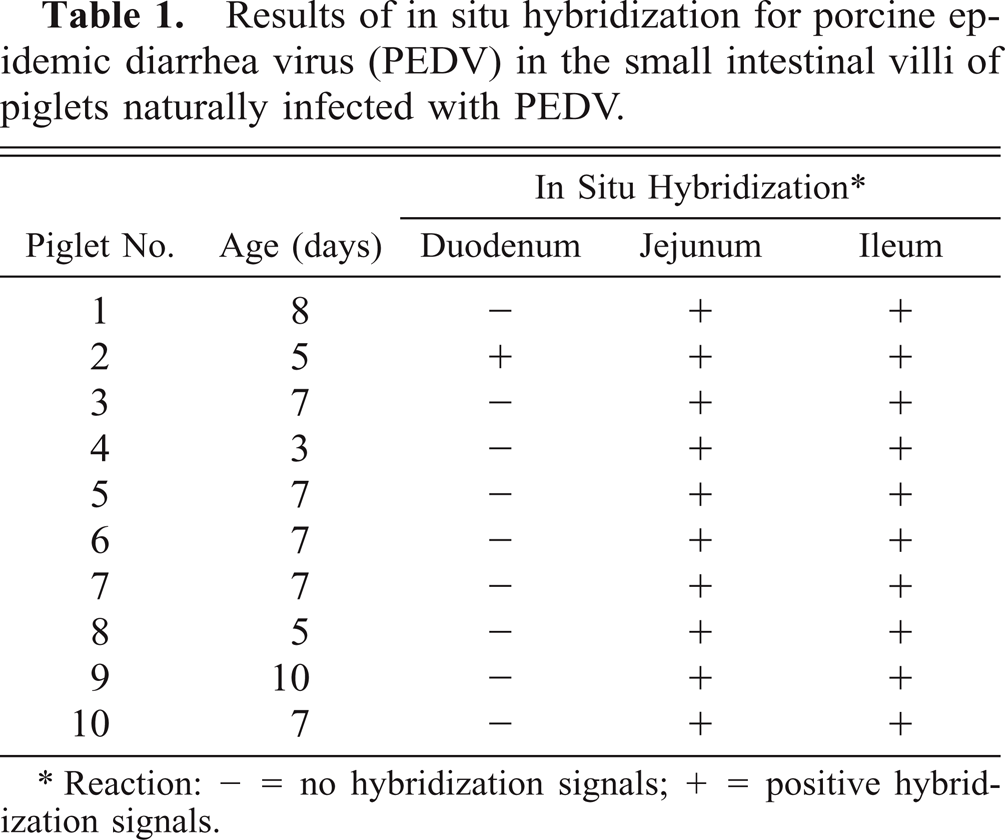
∗Reaction: − = no hybridization signals; + = positive hybridization signals.
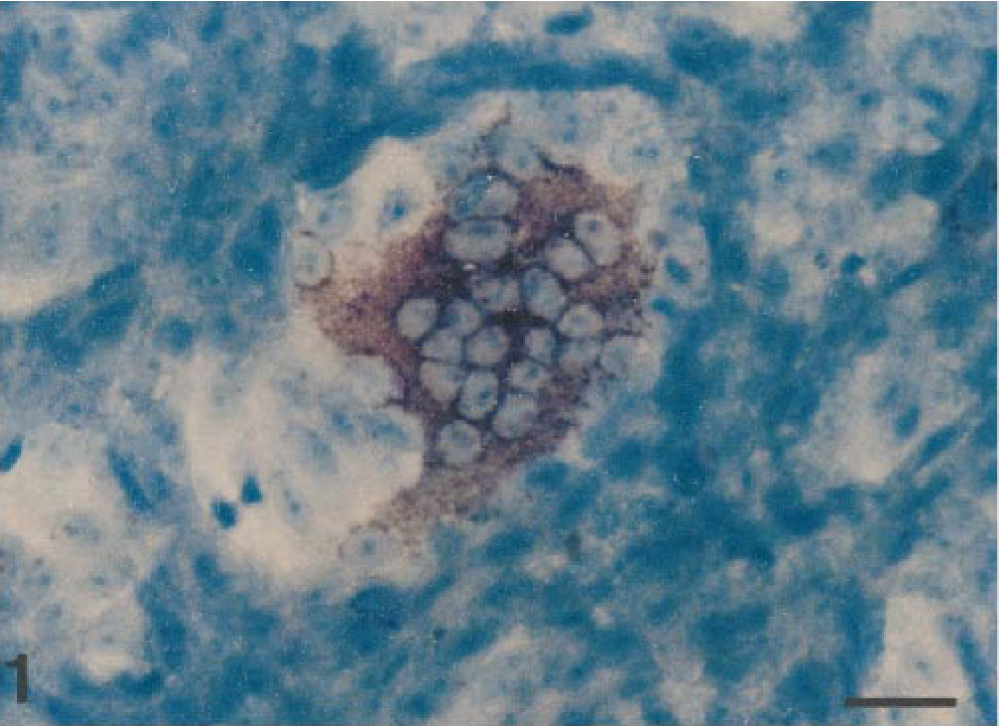
PEDV-infected Vero cells hybridized with a DIG-labeled cDNA probe (377 base pair). PEDV RNA (dark brown reaction) appears in the cytoplasm of a syncytial cell. In situ hybridization; DNA probe, nitroblue tetrazolium/5-bromocresyl-3-indolylphosphate, methylgreen counterstain. Bar = 80 μm.
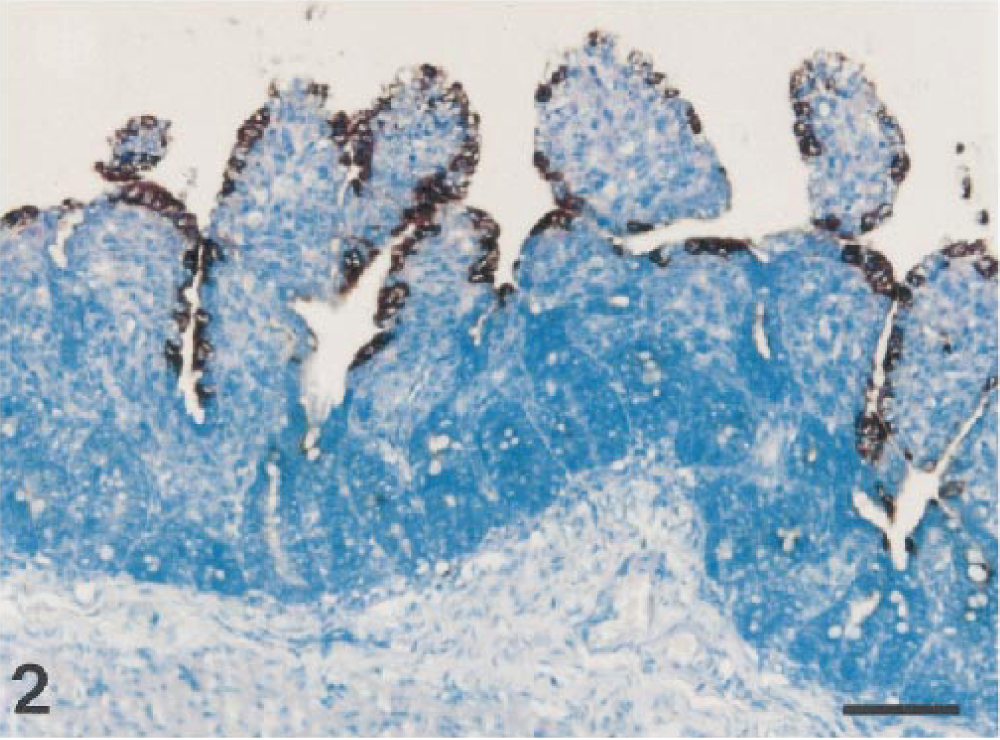
Jejunum; piglet No. 1. PEDV RNA (dark brown reaction) is present in the epithelium covering moderately atrophic villi. In situ hybridization; DNA probe, nitroblue tetrazolium/5-bromocresyl-3-indolylphosphate, methylgreen counterstain. Bar = 80 μm.
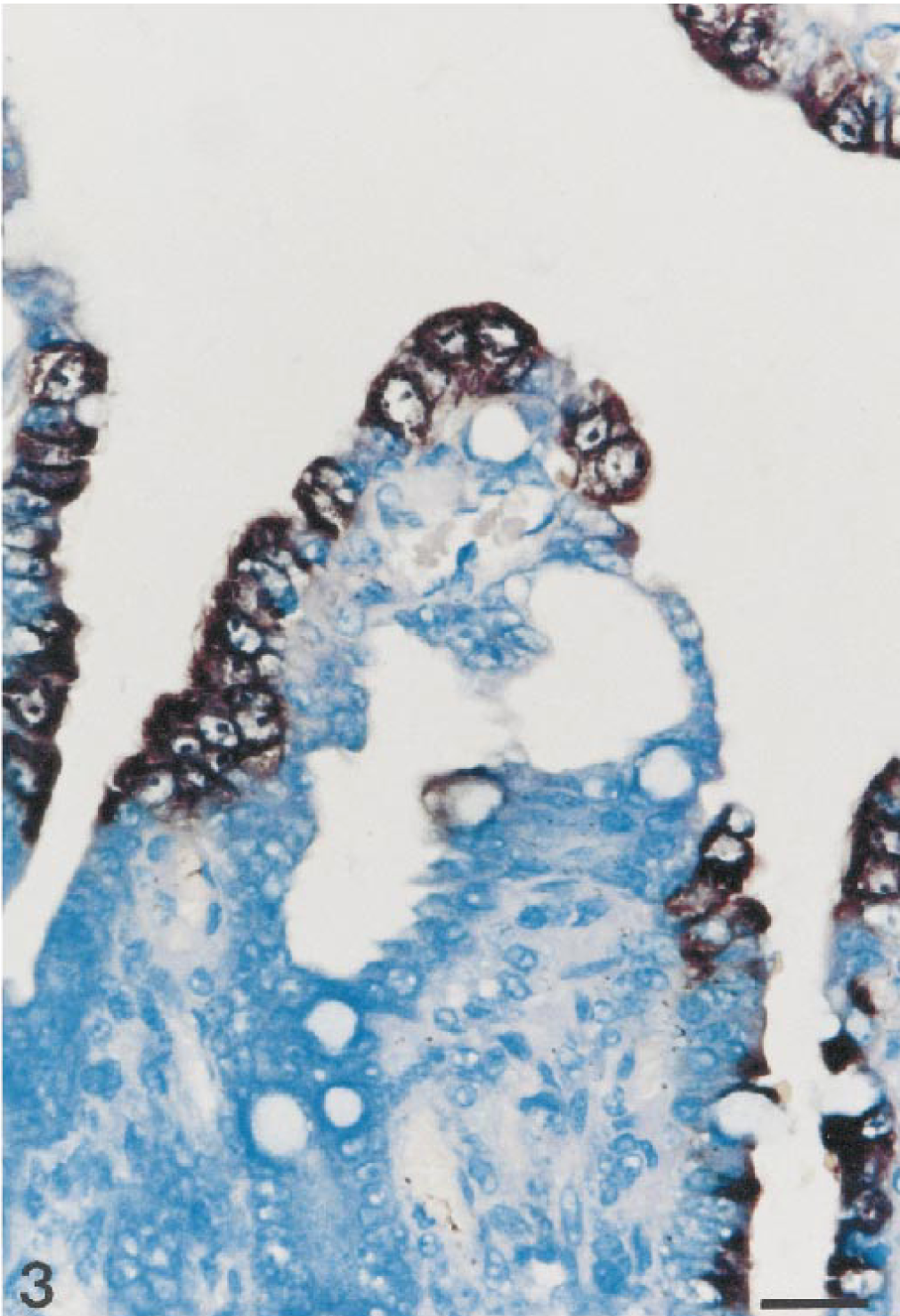
Ileum; piglet No. 6. PEDV RNA (dark brown reaction) is present in the cytoplasm of villus enterocytes. In situ hybridization; DNA probe, nitroblue tetrazolium/5-bromocresyl-3-indolylphosphate, methylgreen counterstain. Bar = 55 μm.
In the jejunum and ileum, there were strong hybridization signals in the villi, typically in areas of moderate to severe villus atrophy or vacuolation. Positive cells were arranged continuously in the enterocytes covering the villi of the small intestine. Cuboidal and degenerated or regenerated enterocytes with or without vacuolated cytoplasm were detected in some areas. In the lumen, numerous exfoliated enterocytes had strong hybridization signals. Hybridization signals were not detected in epithelial cells lining the crypts of Lieberkühn. The ileal dome epithelial cells occasionally showed hybridization signals. Most positive cells were scattered across the dome. In piglet No. 2, hybridization signal was also found in the duodenum. Only a few focal areas with hybridization signal were seen in the duodenal enterocytes.
Hybridization signals were not detected in epithelial cells of the cecum, colon, or rectum. Lung, tonsil, and stomach specimens were negative for in situ hybridization, and PEDV nucleic acids were not detected in any tissue from TGEV- or rotavirus-infected piglets or from the negative control pigs with no exposure to PEDV.
Discussion
The results of this study demonstrate that PEDV nucleic acids can be detected in formalin-fixed, paraffin-embedded tissue specimens of piglets with the epizootic form of the disease with a nonradioactive-labeled DNA probe complementary to a conserved region of the membrane protein gene. PEDV antigen has been detected in jejunal and ileal villus enterocytes of swine by monoclonal and polyclonal antibody–based immunohistochemical procedures.16,21,29 But by using DNA probes and in situ hybridization, there is no possibility for error that could be caused by antigenic cross-reactivity or by the alteration of binding sites caused by tissue processing. Formalin fixation can denature antigens, and this ultimately can lead to false-negative results.17,28 In contrast to immunohistochemistry, in situ hybridization is less susceptible to structural alteration caused by fixation.4,6 Although antigenic cross-reactivity might not be a problem with in situ hybridization, the specificity of nucleic hybridization has its own limitations. To avoid nonspecific hybridization of the probe, both reverse transcription and the PCR must be performed under stringent conditions.
There were villus atrophy and fusion of the enterocyte replicating PEDV as an indication of a direct pathogenic effect of virus. Ultrastructural studies have described considerable damage to the villus enterocytes because of infection with PEDV.13,14,27,29 These changes compromise the digestion and absorption functions of villus enterocytes. Destruction or damage to these cells by PEDV may induce villus atrophy and fusion, which decreases intestinal surface area, resulting in intestinal maldigestion and malabsorption, severe diarrhea, and dehydration.5,14 PEDV is normally described to affect the proximal and distal jejunum and also the ileum.9,10,27 In one piglets (pig No. 2), hybridization signal was also found in the duodenum. Because of the limited number of animals, explaining this difference in sites of replication of PEDV was not possible. This difference could be because of different strains of virus, difference in the age of the piglets, difference in the duration of infection, or difference in susceptibility of breed. Further studies are needed to explain this difference in sites of viral replication.
Histopathologic changes caused by PEDV should be differentiated from other enteric viral diseases such as TGEV and rotavirus. Comparison of histologic intestinal lesions in PEDV- and TGEV-infected piglets in this study did not reveal significant differences. Both virus infections can cause severe villus atrophy in the proximal and distal jejunum and also in the ileum. This is in contrast to rotavirus infection, where villus atrophy is found only in the distal jejunum and ileum.27 Because the nonradioactive-labeled DNA probe for PEDV did not cross-react with small intestines of a naturally TGEV-infected piglet in the present study, in situ hybridization is useful to differentiate PEDV- from TGEV-infected piglets. PEDV-positive cells were demonstrated in villus enterocytes of jejunum and ileum.
Ultrastructural studies have demonstrated PEDV release mainly from villus enterocytes of jejunum and ileum but also from superficial enterocytes of colon and cecum.9,10,27 PEDV antigen has been detected by monoclonal and polyclonal antibody–based immunohistochemical procedures in the small intestine, cecum, and colon from naturally infected piglets.28 PEDV antigen also has been detected in the small and large intestines of experimentally infected piglets by direct immunofluorescent antibody test of frozen tissue sections with polyclonal serum.9,10 Virus replication in the large intestine has also been described in calves infected with bovine coronavirus.30 Replication of PEDV in superficial enterocytes of colon and cecum could not be demonstrated in this study. This difference in the site of viral replication may be because of differences in tissue tropism among PEDV strains. The Korean PEDV appeared to have a tropism for villus enterocyte in the jejunum and ileum of the piglet in this study. In a previous study, positive fluorescence for PEDV was observed in the colon of pigs with diarrhea of greater than 6 hours' duration.10 Thus, piglets in this study might have been euthanatized before the disease had progressed to colonic involvement.
In situ hybridization described in this paper successfully detected PEDV nucleic acids in formalin-fixed, paraffin-embedded intestinal tissues from naturally infected piglets. Because probe of PEDV did not hybridize with TGEV-infected cell cultures, this method would be of particular value in countries where both PED and transmissible gastroenteritis are epizootic and where differentiation between PEDV and TGEV infection for diagnostic purposes is often difficult.5
Footnotes
Acknowledgements
The research reported here was supported by the Ministry of Agriculture, Forestry and Fisheries–Special Grants Research Program (MAFF-SGRP), Republic of Korea.