
Abstract
Canine osteosarcoma (OS) has been used as a model system for the study of cancer biology and treatment despite the lack of information regarding its pathogenesis. Expression of tumor suppressor genes known to participate in malignant transformation were studied in five different OS cell lines. Each of the cell lines exhibited properties of transformed cells, and those that were tested grew in soft agarose and formed osteoid-containing tumors when injected subcutaneously into nude mice. p53 function was determined to be defective in each cell line as indicated by the lack of induction of p53-responsive genes, p21 and mdm2, following treatment with 5-fluorouracil. p53 mRNA and protein levels were elevated in three cell lines and were extremely low in two cell lines. p53 protein overexpression correlated with the presence of mutations within the DNA binding domain. Four cell lines appeared to contain normal retinoblastoma (Rb) mRNA and Rb protein and no detectable p16 mRNA or protein. In contrast, the remaining cell line contained high levels of p16 mRNA and protein and significantly reduced levels of Rb, p107, and p130 proteins. These results underscore the importance of inactivating p53 and Rb family pathways in canine OS and suggest that unlike human OS, cells derived from canine OS contain mutations that simultaneously inactivate all three Rb family members.
Canine osteosarcoma (OS), like its human counterpart, is an aggressive, highly metastatic tumor. Whereas human OS is relatively rare, canine OS is prevalent, occurring primarily in large and giant breeds.37 Canine OS generally occurs in older dogs, although a significant percentage of young animals develop the disease. Human and canine OS are histologically similar and both are difficult to manage clinically because of the known biological behavior of the tumor.32,42
The pathogenesis of human OS has been associated most frequently with abnormalities of the retinoblastoma (Rb) and p53 tumor suppressor genes. The Rb family consists of three genes, Rb, p107, and p130, that produce proteins that regulate cell proliferation and differentiation.10,14,19,26,47 Normally, Rb protein regulates cell cycle transit by controlling the availability and activity of specific members of the E2F family of transcription factors.47 Inactivation of Rb protein by phosphorylation and the subsequent release of E2F is controlled during mid-G1 by D-type cyclins and cyclin-dependent kinases (cdks) 4 and 6.34,47 In response to growth-inhibiting signals, a cdk inhibitor (cki), such as p16, blocks Rb phosphorylation and prevents DNA synthesis. Human cancers frequently contain mutations that inactivate the Rb pathway either by decreasing the inhibitory activity of Rb and p16 or by deregulating the activity of cyclin D or cdk 4.1,4,16,22 Mutations in p107 or p130 are extremely rare in human cancers.40
p53 acts in part through the Rb pathway to inhibit cell cycle transit. In response to cellular stress and/or DNA damage, activated p53 induces the transcription of mdm2, a negative regulator of p53 protein stability, and the cki p21, which blocks Rb phosphorylation.39 In addition, tumor cells containing activated oncogenes undergo p53-mediated apoptosis in response to DNA damage, hypoxia, or chemotherapeutic agents.13,24,30 Tumors that have functionally inactive p53 and Rb pathways are in general more resistant to apoptosis and likely accumulate additional mutations that contribute to tumor progression.
Little is known about the molecular pathogenesis of OS in dogs. Immunohistochemical studies have indicated that p53 is overexpressed in a high percentage of these tumors.38 Mutations within the p53 gene have recently been identified in canine OS.23,44 In one study, nine of 10 tumors were missing a 3.7-kb Rb-specific genomic fragment that was present in normal controls.43 These results suggest that inactivation of these tumor suppressors is an important event that contributes to the development of canine OS. To directly determine whether p53 and Rb pathways are dysfunctional in canine OS, we studied the expression of these genes at the RNA and protein levels in five established canine OS cell lines.
Materials and Methods
Isolation of osteosarcoma cells and cell culture
Animals diagnosed with OS based upon radiologic and histologic findings were used for the establishment of continuous cell lines. Tumor designation and location and the age of the dogs are as follows: CO2, distal femur, 2 years; CO3, distal radius, 5 years; CO5, distal radius, 10 years; CO7, proximal humerus, 2 years; CO8, proximal humerus, 7 years. Canine OS cells were isolated following trypsin (0.15%) and collagenase (0.3%) treatment of fresh tissue from primary tumors. Care was taken to exclude surrounding connective tissue containing contaminating fibroblasts. Enzymatically released cells were grown in Dulbecco's modified Earle's medium (DME) containing 5% bovine fetal calf serum (FCS), 4 mM glutamine, and penn-strep/fungizone (Life Technologies, Gaithersburg, MD). Low-passage-number cells (<25) were used for all experiments. Control canine fibroblasts (CF2) were isolated from connective tissue adjacent to normal bone. For experiments testing p53 function, cells were treated for 20 hours with 2.5 μM 5-fluorouracil (5FU) and harvested for RNA or protein extraction as described below.
Injection of tumor cells into nude mice
Trypsinized CO2 and CO3 cells were resuspended in DME plus 5% FCS and counted, and 2 × 106 cells in 100 μl DME were injected subcutaneously into the right upper thigh of nude mice (three each) (Charles River Laboratories, Wilmington, MA). Tumors were harvested after 6 weeks. Sections prepared from formalin-fixed, paraffin-embedded samples were stained and used for histologic examination.
RNA analysis
RNAs for Rb, p107, p130, mdm2, p21, p53, and p16 were quantitated by northern analysis in all of the cell lines. Equal OD260 units of total cellular RNA (20 μg) were separated on a 6% formaldehyde, 1% agarose gel. RNA was transferred to nylon membranes (ICN, Costa Mesa, CA) and baked for 2 hours at 80 C. Isolated cDNAs were radioactively labeled by random priming and hybridized in 50% formamide at 42 C. Blots were washed, dried, and autoradiographed with Kodak MS XAR film and an intensifying screen. Densitometry was performed using the Eagle Eye II (Stratagene, La Jolla, CA).
Western blotting
For western blotting, CF2 and canine OS cell lines were harvested in ice-cold RIPA buffer (150 mM NaCl, 1% nonidet-P40, 0.5% deoxycholate, 0.1% sodium dodecyl sulphate [SDS], 50 mM Tris, pH 8.0) containing phenylmethylsulfonylfluoride (1 mM), aprotinin (1%), leupeptin (20 μg/ml), and sodium orthovanidate (1 mM). Equal amounts of protein were electrophoresed through 6% (for Rb, p107, and p130), 8% (for p53), or 12% (for p16) SDS polyacrylamide gels. Protein bands were transferred to nitrocellulose using a semidry electroblotter (Bio Rad, Hercules, CA). Filters were blocked in 4% milk powder in Tris-buffered saline containing 0.05% Tween 20 and incubated with specific antibodies overnight at 4 C. After washing, filters were incubated with the appropriate horseradish peroxidase-conjugated secondary antibody, washed, and then processed for detection using the ECL kit (Amersham, Arlington Heights NJ). Anti-p53 (pAb240), anti-Rb (C-15), anti-p107 (C-18), anti-p130 (C-20), anti-p16 (H-156), and anti-cdk2 (M2) were obtained from Santa Cruz Biotechnology Inc. (Santa Cruz, CA) and used per the manufacturers' instructions.
Serum starvation and Rb phosphorylation state
CF2 and OS cell lines were serum starved in 0.5% and 0.3% FCS, respectively, for 72 hours. At that time, cell proliferation had ceased (CF2, CO2, CO5, CO7) or was significantly reduced (CO3, CO8). Cell extracts were prepared for western blotting as described above. Extracts were electrophoresed through a 6% SDS polyacrylamide gel. Hyperphosphorylated and hypophosphorylated Rb proteins were identified with an anti-Rb antibody based upon position in the gel.
Cloned DNAs
Probes for canine p53, human p21, p107, p130, and mdm2 were obtained by polymerase chain reaction (PCR) using oligonucleotides derived from published DNA sequences. Murine Rb was provided by Dr. Rene Bernards (Massachusetts General Hospital, Charlestown, MA). Human p16 was provided by Dr. Leonard Freedman (Memorial Sloan-Kettering Cancer Center, New York City, NY).
Sequencing of p53
RNA isolated from canine OS cell lines containing detectable p53 protein was used to synthesize cDNA using AMV reverse transcriptase (Promega, Madison, WI). The cDNAs (0.1 μg) were amplified using canine p53-specific primers (forward: 5′-CTGGCTAGACGAAGACTCAG-3′, reverse: 5′ AGGCAGTGCTCGCTTGGTAC-3′) and Taq DNA polymerase (Life Technologies). The 738-bp PCR product, representing amino acids 52-296, was cloned into the T vector (Promega), and DNA was sequenced using the dsDNA Cycle Sequencing System (Life Technologies). Mutant sequences were verified by sequencing at least three independent PCR clones for each cell line.
Results
Properties of OS cell lines
All five canine osteosarcoma cell lines exhibited properties of transformed cells in culture, including loss of contact inhibition and reduced growth factor requirements. CO2 and CO8 cultures consisted predominantly of flat, spindle-shaped cells, whereas CO3 and CO7 cultures contained round to spindle-shaped cells that did not adhere well to the plastic growth surface. In contrast, CO5 cultures were more homogeneous and polygonal in shape. This appearance may relate to the parent tumor being diagnosed as a chondroblastic osteosarcoma. CO2 and CO3 formed colonies in soft agarose.
When injected subcutaneously into nude mice, CO2 and CO3 formed osteoid-containing tumors. CO2 tumors grew slowly and consisted of irregularly shaped, pleomorphic cells. Large areas of osteoid-like material were evident (Fig. 1). In contrast, tumors derived from CO3 grew rapidly and appeared more invasive. Sections from these tumor contained distinct areas of irregularly shaped and spindle-shaped cells (Fig. 2). Osteoid-like material was generally not associated with spindle-shaped cells.
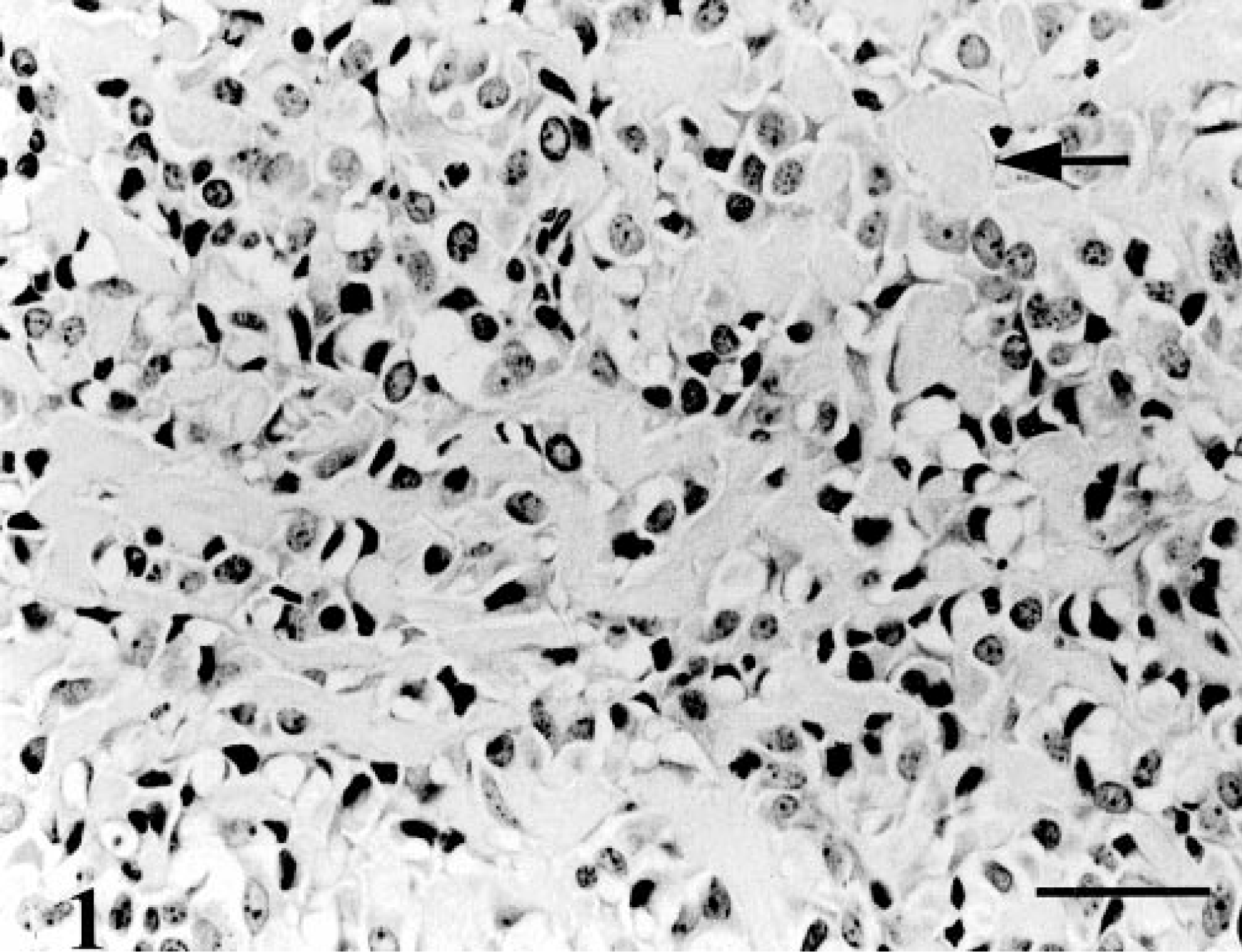
Osteoid-containing tumor; nude mouse injected with cells from CO2. Large amounts of osteoid-like material were present (arrow). HE. Bar = 100 μm.
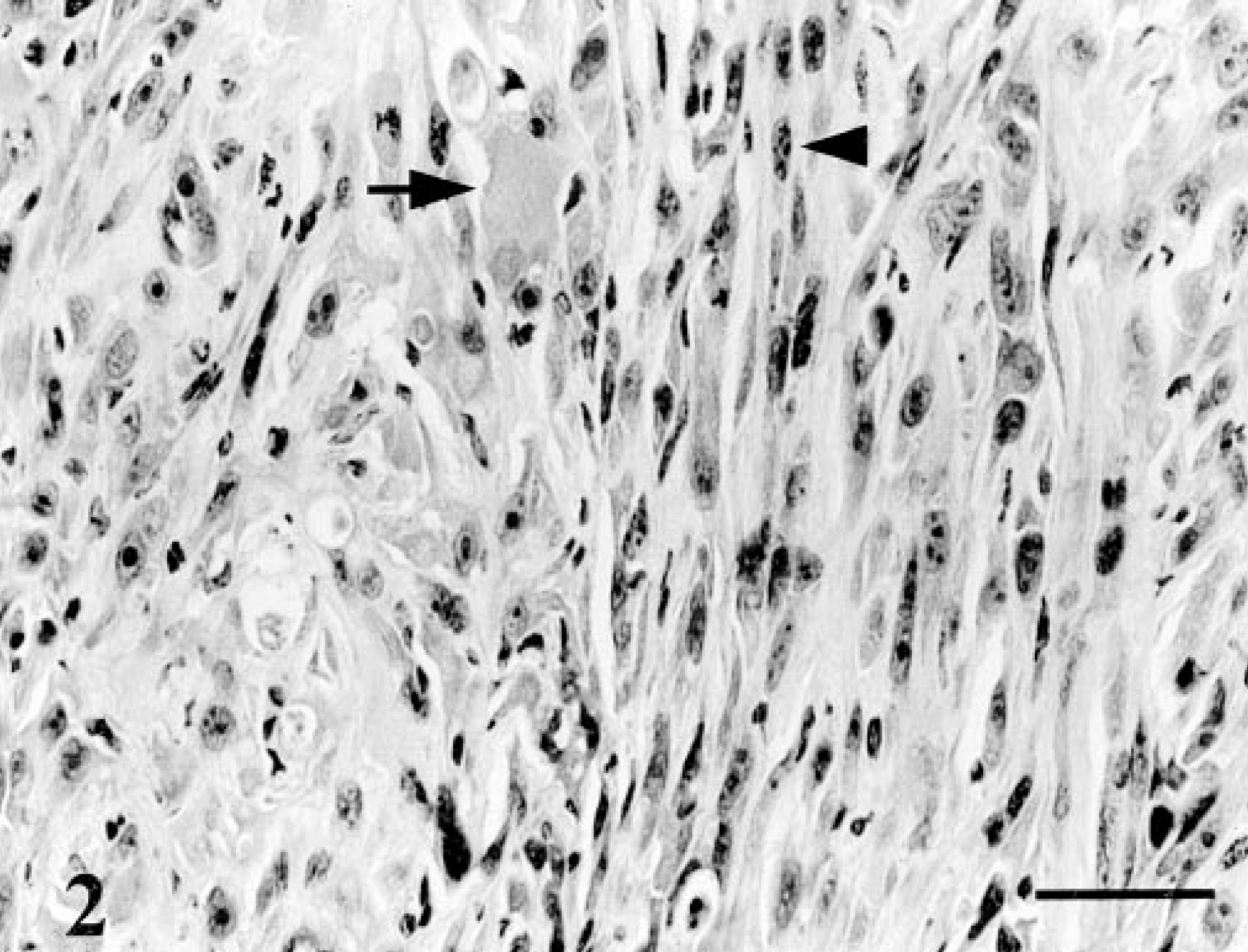
Osteoid-containing tumor; nude mouse injected with cells from CO3. Tumors contained spindle-shaped cells (arrowhead) and some osteoid-like material (arrow). HE. Bar = 100 μm.
Analysis of p53 gene expression
p53 is activated in response to various stimuli, including, agents that damage DNA or promote nucleotide pool imbalance.30 To test for p53 function, OS cells were treated with 2.5 μM 5FU for 20 hours and assayed for the expression of p53-inducible genes, p21 and mdm2 (Fig. 3).29 Normal CF2 were used as a control to verify the p53 wild-type response. In the presence of 5FU, p21 and mdm2 mRNA levels increased by >20-fold in CF2 cells. Although baseline levels of p21 were higher in CO2, CO3, CO5, and CO8 than in untreated CF2 cells, a <4-fold induction of p21 mRNA was detected in any of the OS cell lines. The mdm2 mRNA levels remained relatively constant in all of the OS cell lines and were higher than in untreated CF2 cells. These results indicate that the induction of p21 and mdm2 is defective in these canine OS cell lines.
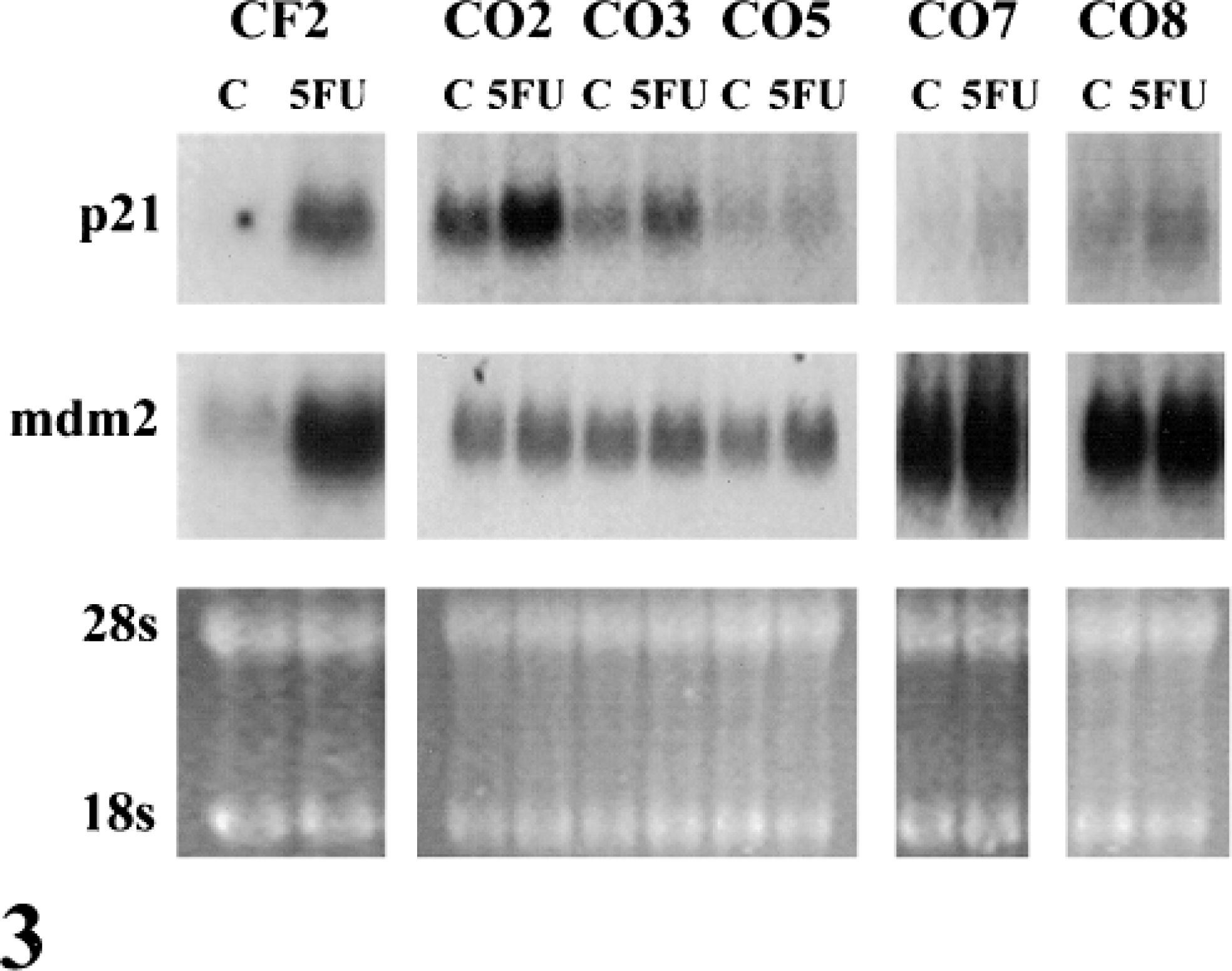
Northern blot of p21 and mdm2. Control primary canine fibroblasts (CF2) and five canine OS cell lines (CO) were treated with 5FU (2.5 μM) for 20 hours, and the induction of p21 and mdm2 mRNA was assayed by northern hybridization. Ethidium bromide-stained ribosomal 28S RNA and 18S RNA bands are shown for each sample as a control for equal loading.
Rearrangements and/or deletions of both copies of the p53 gene are common in human OS.8 In these cases p53 mRNA is usually not detected or is of aberrant size. Three canine OS cell lines (CO2, CO7, CO8) had high levels of normal size p53 mRNA as compared with CF2 cells (Fig. 4). The remaining two cell lines (CO3, CO5) expressed extremely low or undetectable levels of p53 mRNA. This low level of expression likely accounts for the p53 functional deficiency observed in CO3 and CO5.
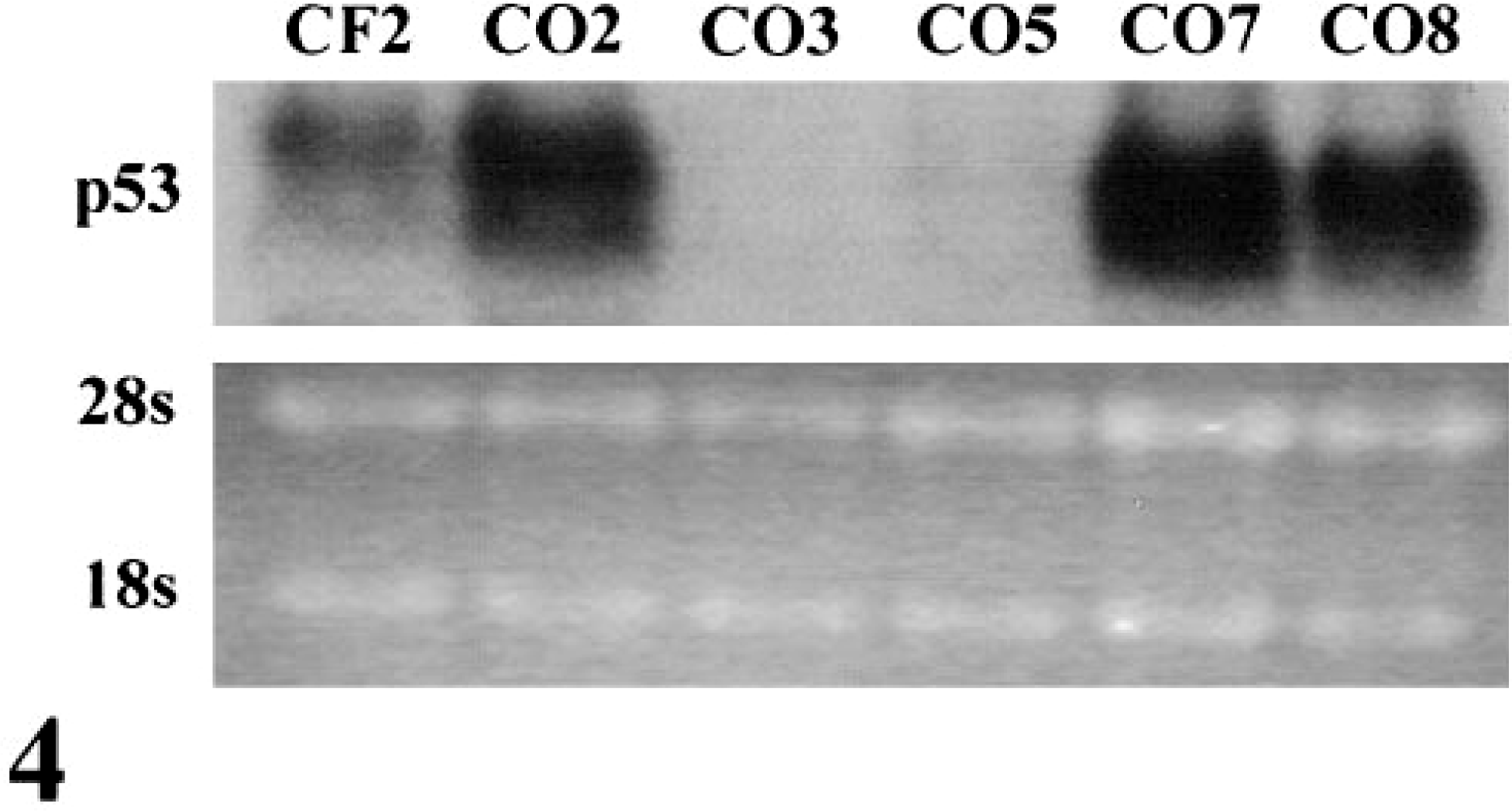
Northern blot of p53 in exponentially growing cells. p53 mRNA levels were very low or undetectable in CO3 and CO5.
Point mutations within the p53 gene are very common in human cancers. A hallmark of many p53 mutations is enhanced protein stability of this normally labile protein, enabling detection by immunohistochemistry and western blotting.28 p53 protein could not be detected in normal CF2 nor in CO3 and CO5 cells (Fig. 5). In contrast, CO2, CO7, and CO8 expressed high levels of p53 protein. The overexpression of p53 protein in 3/5 (60%) of the OS cell lines is consistent with published results indicating that p53 is overexpressed in a high percentage of canine OSs.38

Western blot of p53 protein in exponentially growing cells. p53 protein was overexpressed in CO2, CO7, and CO8.
Using primers derived from the canine p53 gene, regions of p53 known to be frequently mutated in human cancers were amplified and cloned from CO2, CO7, and CO8 cells, and the DNA was sequenced.45 CO2 and CO8 both contained mutations at amino acid residue 269 (a G to A mutation), converting an aspartic acid to an asparagine. CO7 contained a mutation at residue 261 (a GC to AA mutation), converting an arginine to a glutamine. These amino acids correspond to residues 281 and 273, respectively, in the human p53 gene. Mutations at both sites have been identified in human tumors.36 The locations of these mutations within the highly conserved sequence-specific DNA binding domain are consistent with mutations that impair p53 function.27,36
Analysis of Rb family gene expression
The tumor suppressor gene Rb is mutated in a high percentage of human OSs. In the majority of cases, Rb mutations are characterized by loss of Rb mRNA and/or protein.46 During exponential growth, all five OS lines contained elevated levels of full-length Rb mRNA as compared with CF2 (Fig. 6). CO3, CO5, CO7 and CO8 contained correspondingly high levels of normal size Rb protein (Fig. 7). In contrast, CO2 contained markedly reduced levels of Rb protein. The majority of Rb protein in all cases was hyperphosphorylated during exponential growth (Fig. 7).
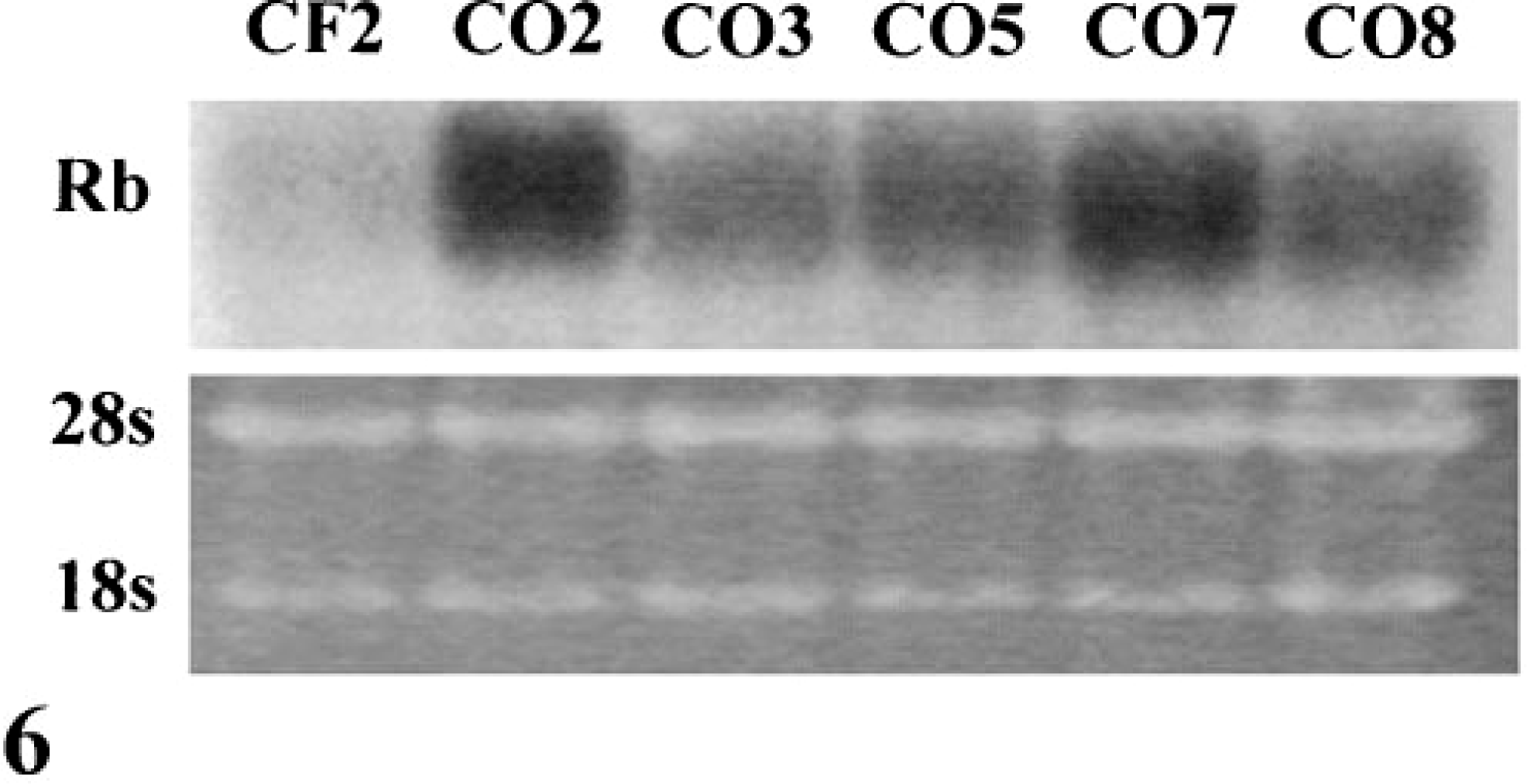
Northern blot of Rb in exponentially growing cells. Rb mRNA was expressed at high levels in all OS cell lines.
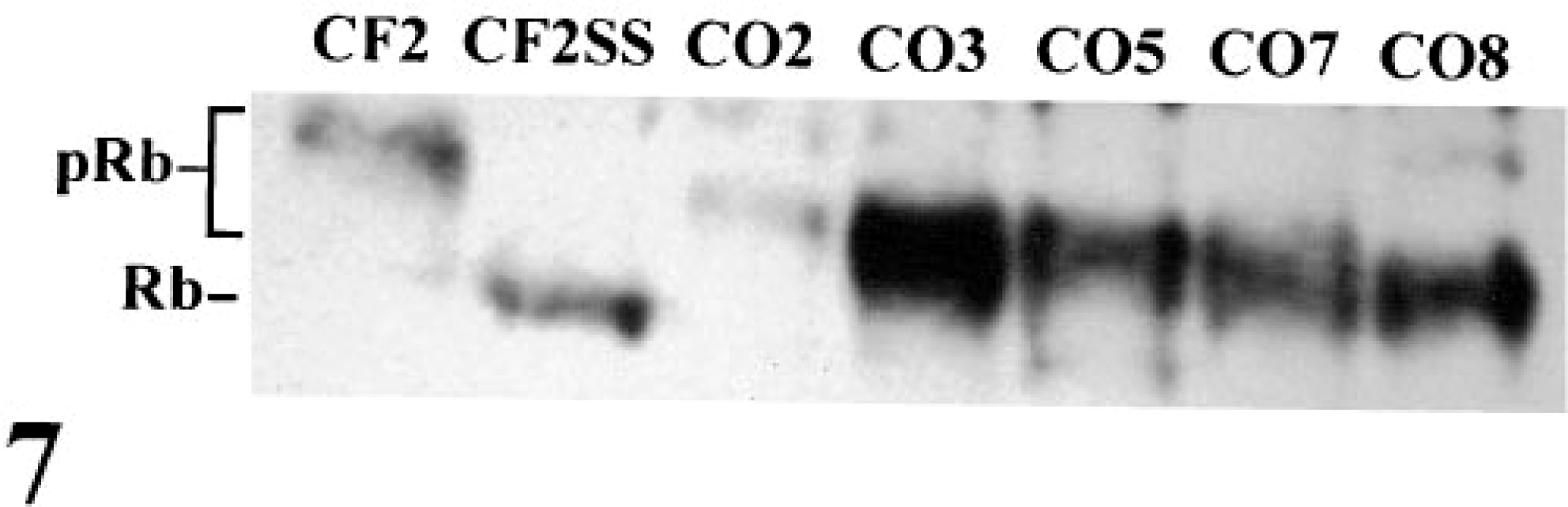
Western blot of Rb protein in exponentially growing cells. Both hypophosphorylated (Rb) and hyperphosphorylated (pRb) forms of the protein were resolved. As a control for the expression of the hypophosphorylated form of Rb, CF2 cells were serum starved (CF2SS) for 72 hours in 0.5% fetal bovine serum prior to the preparation of protein extracts. Rb protein was expressed at low levels in CO2.
The expression patterns of Rb family members p107 and p130 were similar to those observed for Rb. CO2 contained elevated levels of p107 and p130 mRNA (Fig. 8). CO3, CO5, CO7, and CO8 expressed both proteins (Fig. 9) whereas CO2 contained little or markedly reduced levels of p107 and p130. The reduced expression of p107 and p130 in CO2 was specific for these proteins; several other proteins, including cdk2, were expressed at high levels (Fig. 9).
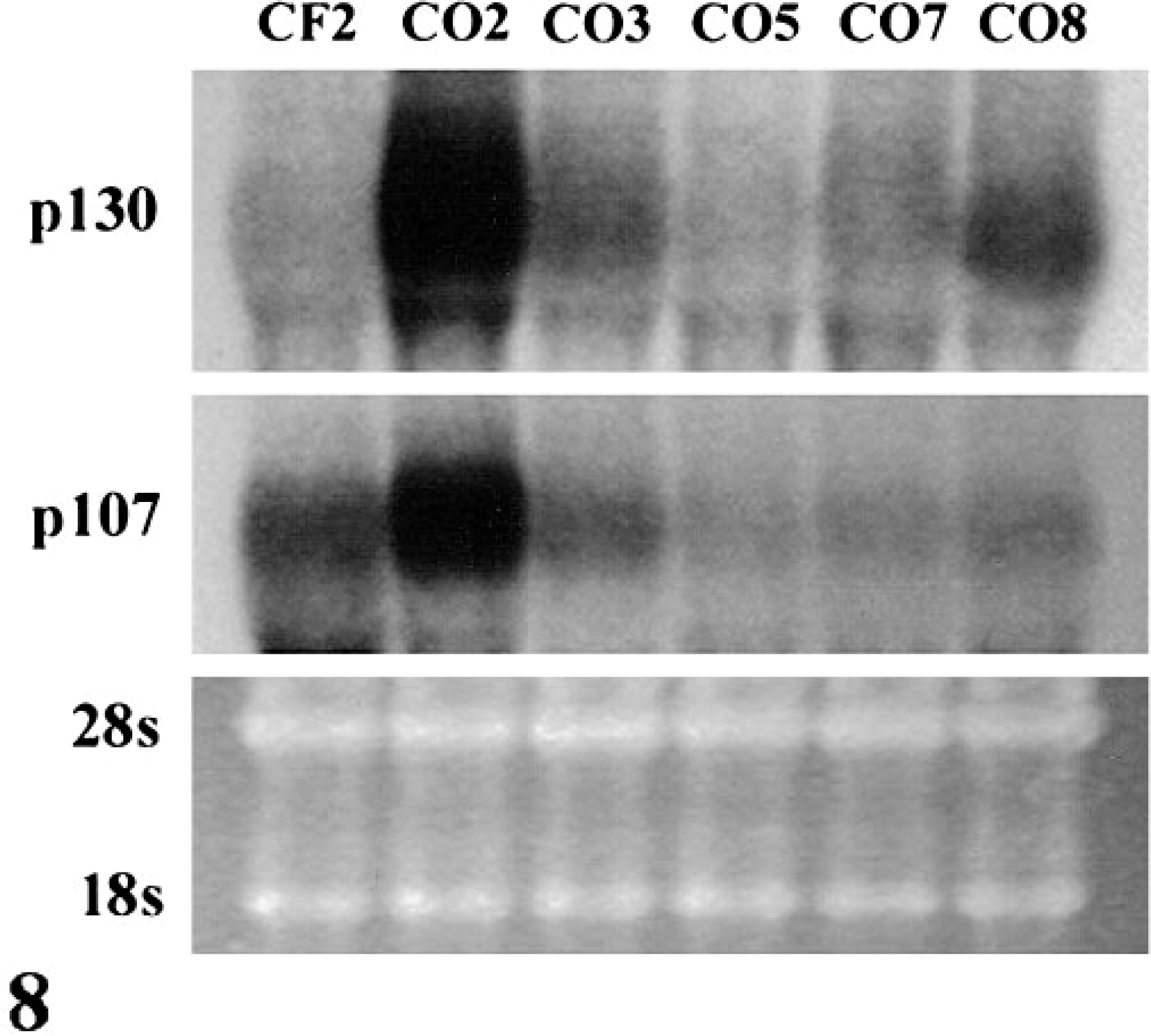
Northern blot of p107 and p130 in exponentially growing cells. p107 and p130 mRNAs were expressed in all OS cell lines.
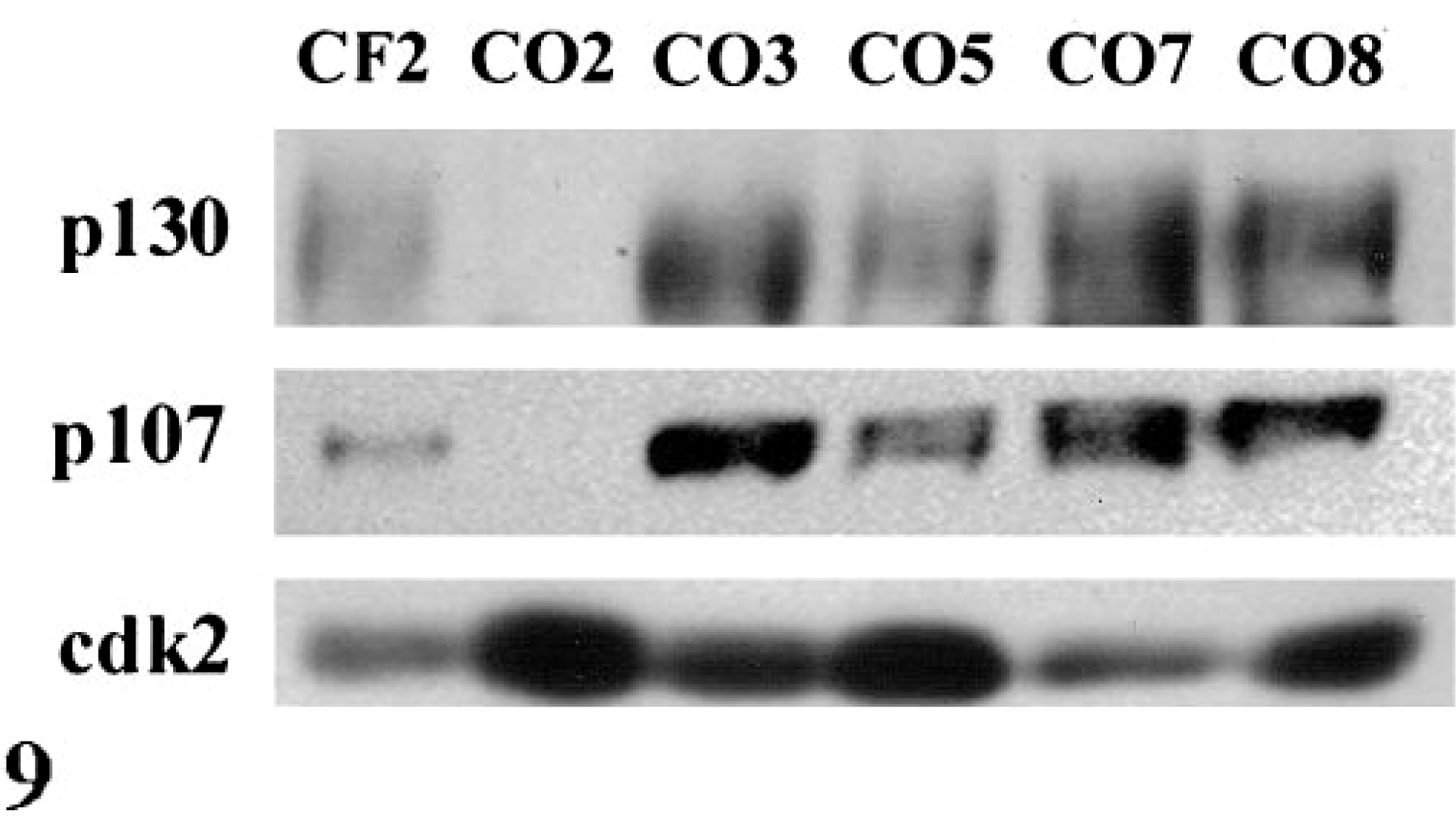
Western blot of p107 and p130 protein in exponentially growing cells. p107 and p130 proteins were undetectable in CO2, whereas cdk2 protein was abundant.
One measure of altered growth control is the failure of hypophosphorylated Rb protein to accumulate following serum starvation.7,12 The normal response, as typified by CF2 cells, is the loss of hyperphosphorylated Rb protein and a concurrent increase in the percentage of hypophosphorylated protein (Fig. 10). All of the OS cell lines contained high levels of hyperphosphorylated Rb protein, relative to CF2, following serum starvation. CO3, CO5, CO7, and CO8 accumulated hypophosphorylated Rb protein, but with the exception of CO5, the hyperphosphorylated form was most abundant.
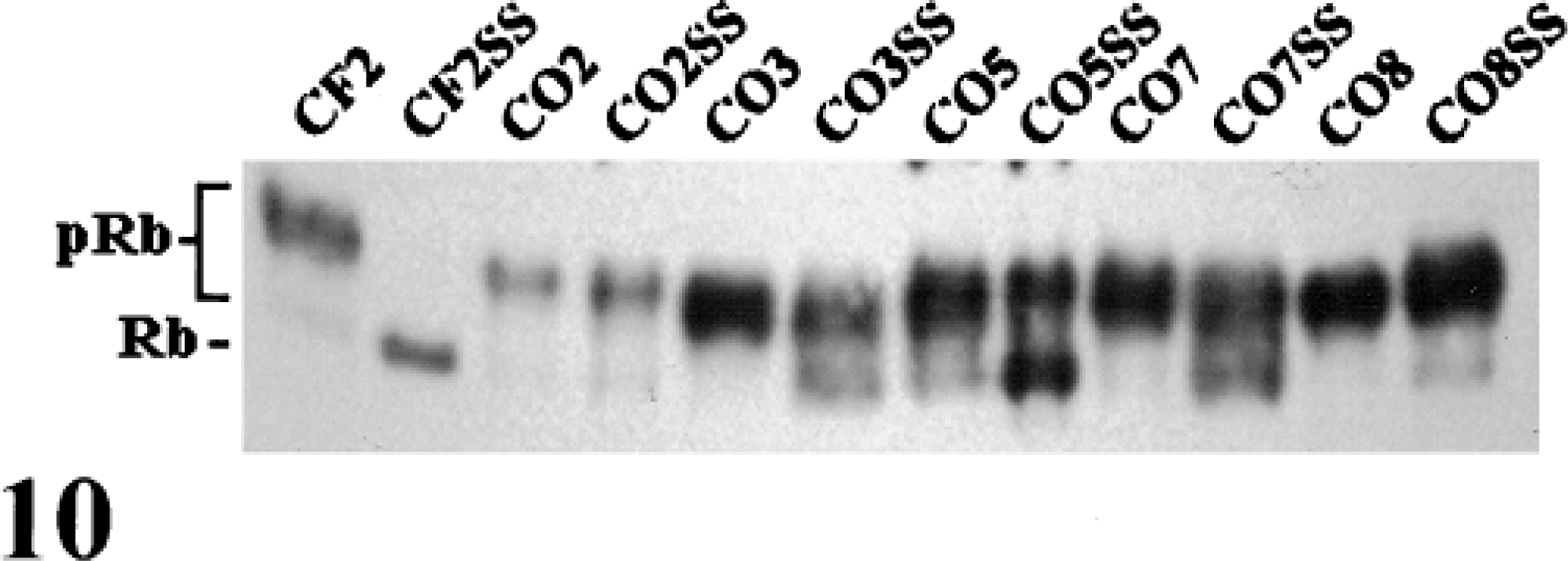
Western blot of Rb protein following serum starvation. Protein extracts from exponentially growing and serum-starved cultures were analyzed for Rb protein by western blotting. The majority of Rb protein in the OS cell lines remained in the hyperphosphorylated state following serum starvation.
Analysis of p16 gene expression
The cki p16 is an important regulator of the Rb pathway and is frequently mutated in human tumor cell lines and tumors.35,39 CF2 contained low levels of p16 mRNA (Fig. 11) and detectable levels of protein (Fig. 12). CO2 contained high levels of p16 mRNA and elevated levels of p16 protein relative to CF2. The p16 mRNA and protein were not detectable in the remaining OS cell lines that contained high levels of Rb protein.
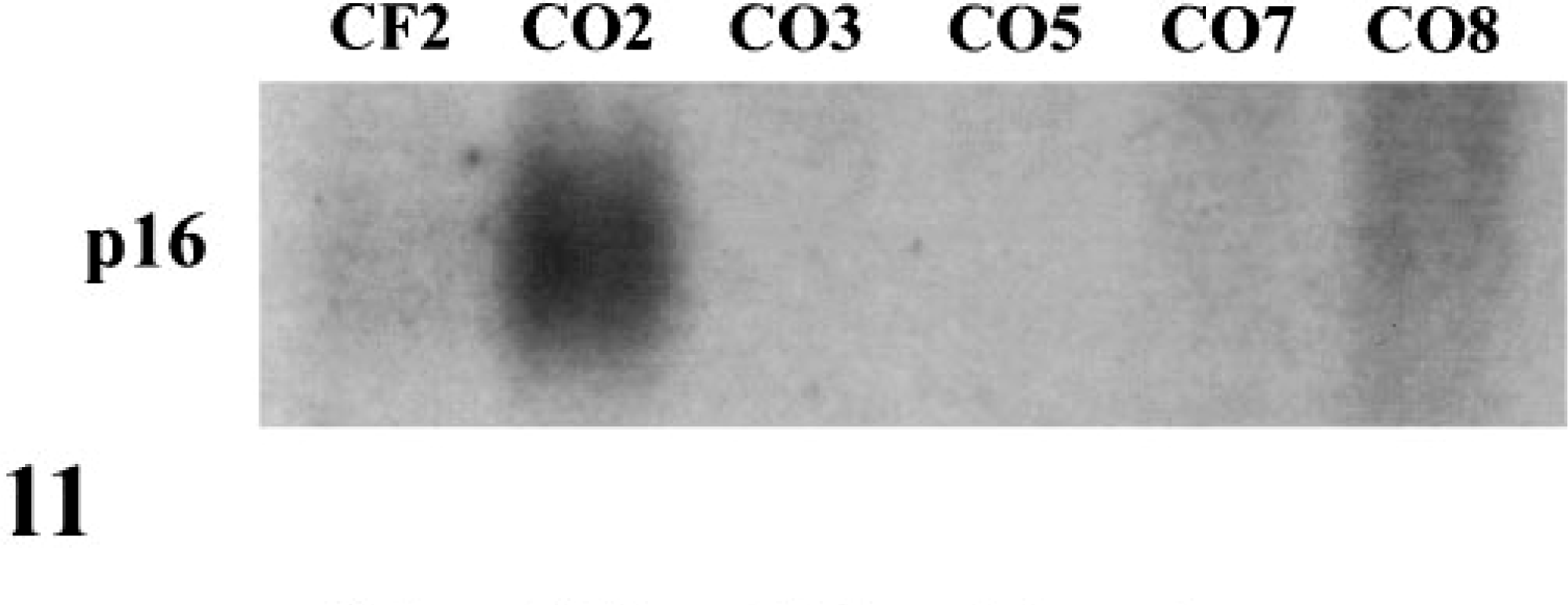
Northern blot of p16 expression in exponentially growing cells. p16 mRNA was undetectable in CO3, CO5, CO7, and CO8.

Western blot of p16 protein in exponentially growing cells. p16 protein was undetectable in CO3, CO5, CO7, and CO8.
Discussion
To determine whether p53 and Rb pathways are misregulated in canine OS, five canine OS cell lines from different animals were established. Several lines of evidence indicate that p53 activity is aberrant in all five canine OS cell lines. First, p53 mRNA and protein levels were undetectable in CO3 and CO5. Second, p53 protein levels were very high in CO2, CO7, and CO8. Elevated levels of p53 protein are common in cells containing mutations within the p53 gene that compromise p53 activity.28 In the absence of a functional p53 protein, its transcriptional target, mdm2, is not present to destabilize the mutant p53 protein. Third, mutations in p53 were detected at amino acid sites 261 in CO7 and 269 in CO2 and CO8. These amino acids are localized to the DNA binding domain of p53, and the equivalent human amino acids are commonly mutated in human cancers.36 Wild-type p53 sequences were not detected in CO2, CO7, and CO8. In an in vivo assay of p53 activity, neither p21 nor mdm2 genes, both transcriptionally induced by wildtype p53, were induced in any of the OS cell lines following 5FU treatment.
Our findings that p53 function is aberrant in all of the canine OS cell lines is consistent with the accepted thinking that p53 is a critical regulatory gene that blocks cell cycle transit and tumorigenesis.29 The importance of p53 as a tumor suppressor for canine OS is further underscored by previous studies indicating that p53 protein is overexpressed in 84% and 56% of OS derived from appendicular and axial locations, respectively.38 Our results suggest that reduced levels of p53 mRNA, possibly resulting from inhibition of transcription and/or alterations of mRNA degradation, may be responsible for compromising p53 activity in tumors in which p53 protein is not detectable by immunostaining. Taken together, these result suggest that mutational inactivation of p53 plays an important role in the pathogenesis of canine OS.
All five canine OS cell lines contained elevated levels of Rb mRNA relative to CF2. Rb or Rb family members may negatively regulate Rb mRNA levels via transcriptional and/or posttranscriptional mechanisms.48 In the absence of an active Rb pathway, Rb mRNA levels would increase to compensate for the loss or inactivity of Rb protein.
The results presented here indicate that the Rb pathway is deficient in all of the canine OS cell lines. Unlike in control fibroblasts, Rb protein in the OS cell lines was found predominantly in the hyperphosphorylated, noninhibitory form following serum starvation. In exponentially growing cells, Rb protein levels were either very low (CO2) or p16 was not expressed (CO3, CO5, CO7, CO8) in these cell lines. In the absence of sufficient Rb or p16, the ability of these cells to respond to negative growth regulatory environmental signals is impaired.
Rb is mutated in many human cancers including OS. In one study, 40/56 (71%) OS were wholly or partially negative for Rb expression.46 In the present study, only 1/5 (20%) canine OS cell lines had reduced expression of Rb protein. In the absence of a cloned canine Rb gene, it is difficult to ascertain whether a subtle mutation exists within the Rb coding region in these cell lines that inactivates the protein. However, the OS cell lines expressing high levels of Rb mRNA and hyperphosphorylated Rb protein are also deficient in p16 expression. Inactivating mutations in both Rb and p16 are rarely found in the same cell because a mutation in either gene is sufficient to disrupt the Rb inhibitory pathway.1,4 It is therefore unlikely that Rb is mutated in these cell lines.
In humans, the occurrence of Rb mutations is restricted to specific tumor types. Individuals that inherit a mutated Rb gene are prone to developing retinoblastoma and OS but not other cancers, such as breast and small cell lung carcinomas, which often contain Rb mutations.14,33 Rb may act as a rate limiting negative regulator of cell cycle transit only in specific cell lineages (e.g., retinoblasts and osteoblasts) and/or only at specific developmental times.9,14,15,18,20,25 Under certain conditions, mutations in Rb would therefore not be effective in deregulating cell cycle control.
In contrast to humans, animals do not readily develop retinoblastoma. Mice genetically engineered to inherit only one functional copy of Rb do not develop retinoblastoma, as do humans inheriting the retinoblastoma susceptibility gene.20 One possible explanation is that Rb family members p107 and/or p130 are able to compensate for the loss of Rb. A retinoblastoma-like disease occurs in mice expressing SV40 large T antigen, which simultaneously inactivates Rb, p107, and p130. 31
Our results suggest that mutations leading to the inactivation of multiple Rb family members are favored in canine OS. Four of the canine OS cell lines do not express p16. This cki blocks the activation of all three Rb family members and is often silenced in tumors by changes in methylation state.2,11,17
In the CO2 cell line, Rb, p107, and p130 mRNA levels were high, whereas protein levels were low or undetectable. It is unlikely that individual mutations in each gene are responsible for this phenotype. Rather, a single mutation acting at the posttranscriptional level likely blocks the synthesis or accumulation of all three Rb family member proteins. The mechanism of action is unknown, but it may involve alterations in protein synthesis or stability. Various proteases have been identified that cleave Rb during apoptosis and following expression of the papilloma immortalizing gene E7.3,6,21,41 It is unclear whether p107 and p130 are also degraded during apoptosis. Neither p107 nor p130 are degraded in E7-expressing cells.5
Canine OS cell lines are distinct from their human counterparts in that they appear to contain normal Rb protein.46 Instead, they likely harbor mutations that may promote tumorigenesis in vivo by simultaneously inactivating all three Rb tumor suppressor gene family members.
Footnotes
Acknowledgements
We are grateful to the surgeons and the Pathology residents at Cornell College of Veterinary Medicine for supplying fresh OS specimens, Dr. James Zimmer and Dave and Mary Maley for donating the osteosarcoma from their beloved dog, Jake, Drs. Andy Goodwin and Achim Gruber for their assistance in injecting mude mice with tumor cells, and Dr. Ron Minor for helpful discussions. This study was supported by a grant from the Morris Animal Foundation (97CA-21).