
Abstract
The purpose of this study was to compare weight gain and food intake during high-fat feeding in outbred CD-1 male mice while considering several different experimental designs. This study was completed using data from three separate experiments and was designed to address different experimental design issues. Experiment 1 compared mice housed in groups or singly. Experiment 2 compared adolescent and young adult mice. Experiment 3 examined mice that had been previously exercise-trained prior to diet-induced weight gain. Data from each experiment were analysed using repeated measures analysis of variance and linear regression. While housing and age did not significantly affect weight gain, mice that were previously exercise-trained consumed significantly more kilocalories than sedentary mice while maintaining comparable body weights. We generated a linear prediction model using data from Experiments 1 and 2 that will allow investigators to calculate the weeks of high-fat feeding needed to reach a target body weight. Our key findings characterize the issues related to and affecting experimental design when utilizing an outbred mouse diet-induced weight gain model and will serve as a guide for future researchers.
The prevalence of overweight and obesity has dramatically increased throughout the world. Recent statistics from the International Association for the Study of Obesity suggest that one-half of the population of many European countries can be considered overweight, and one-fifth are obese (body mass index >30). 1 According to the National Center for Health Statistics, approximately one-third of adults in the United States are considered obese. 2 Excessive weight gain is associated with chronic diseases, such as hypertension, cardiovascular disease and type II diabetes mellitus. 3 Numerous published papers have utilized a murine model of weight gain to investigate various diseases and treatments associated with obesity.4-6 These models are cost-effective and allow researchers to collect lifespan data in a shorter period of time than would be required for human research. Many researchers utilize mice that are genetically bred to overeat; however, outbred mice may be better for longitudinal studies, because they tend to live longer and have fewer age-related complications. 7 Since mouse models are routinely used in obesity research, this study has been designed to document the effects of various experimental design considerations on weight gain and food intake in outbred CD-1 mice consuming a high-fat diet.
The purpose of this study was to compare weight gain and food intake during high-fat feeding in outbred CD-1 male mice while considering several different experimental designs (i.e. housing type, age, exercise). We utilized data collected during the completion of three separate experiments. These experiments were designed to compare and contrast the effects of housing (i.e. individual versus small groups), previous aerobic exercise training and the effect of changing a rodent's diet during adulthood on diet-induced weight gain. The primary objective of this approach is to thoroughly identify and describe factors that may influence body weight gain in CD-1 mice. A secondary purpose was to generate a linear model, which predicts the number of weeks of high-fat feeding required to reach a target body weight in CD-1 mice.
Such information may serve as a foundation for future research in this area.
Animals, materials and methods
Animals
The University of Houston institutional committee for animal care and use approved all protocols used in the present study. The experiments in the present study utilized outbred, Crl:CD1(1CR) male mice of varying ages (Charles Rivers Labs, Portage, MI, USA). All mice were free of the following pathogens: mouse parvovirus (MPV)-1, MPV-2, minute virus of mice (MVM), parvovirus NS-1 (NS-1), mouse hepatitis virus (MHV), Theiler's murine encephalomyelitis virus (TMEV), epizootic diarrhoea of infant mice (EDIM), pneumonia virus of mice (PVM), reovirus III (REO) and Mycoplasma pulmonis (MPUL). It has been previously suggested that outbred mice are a better model for longitudinal studies since they typically live longer and experience fewer age-related complications compared with inbred mice. 7 Mice were housed at the Association for Assessment and Accreditation of Laboratory Animal Care-accredited animal care facility at the University of Houston in solid-bottomed microisolator cages with aspen straw contact bedding (Harlan Laboratories Inc, Madison, WI, USA). Mice were housed either in groups of four in a 34 x 18 x 14 cm cage or individually in a 26 x 16 x 12 cm cage. Cages were changed twice weekly. Enrichment was provided in the form of a chew pad and a house in each cage. All mice were housed on a standard 12:12 h light-dark cycle in a temperature controlled room in the University of Houston animal care facility. The animal facility was maintained at a temperature of 22 ± 1°C. Sentinal mice were routinely tested for the following pathogens: ectromelia, EDIM, TMEV-strain GDVII mouse poliovirus (TMEV-GDVII), lymphocytic choriomeningitis (LCM), MHV, MVM, murine norovirus (MNV), MPUL, MPV, NS-1, PVM, REO and Sendai. Four of the 15 sentinal mice housed with mice used for Experiment 2 tested positive for MNV; however, none of the mice used in our experiment tested positive. Prior to the experiments, mice were acclimatized to the animal care facility for at least 14 days. During this time, they consumed a stock diet.
Diets
The stock diet provided during the acclimatization period was purchased from Purina (Lab Diet 5001 Rodent Diet, Purina, Richmond, IN, USA). The high-fat treatment diet (#D12492) was purchased from Research Diets Inc (New Brunswick, NJ, USA). The two diets differed in macronutrient content (Table 1) and were based on the AIN-76A formulation. Since this diet differs in micronutrient levels from the AIN-93 formulation, it is possible that this may have affected our outcome measures. Food was presented as a weekly bolus dose in a food rack in the wire bar lid of the home cage. Water bottles were also held in the wire bar lid. Mice were provided ad libitum access to food and water throughout all experiments.
Macronutrient breakdown of the stock and high-fat diets
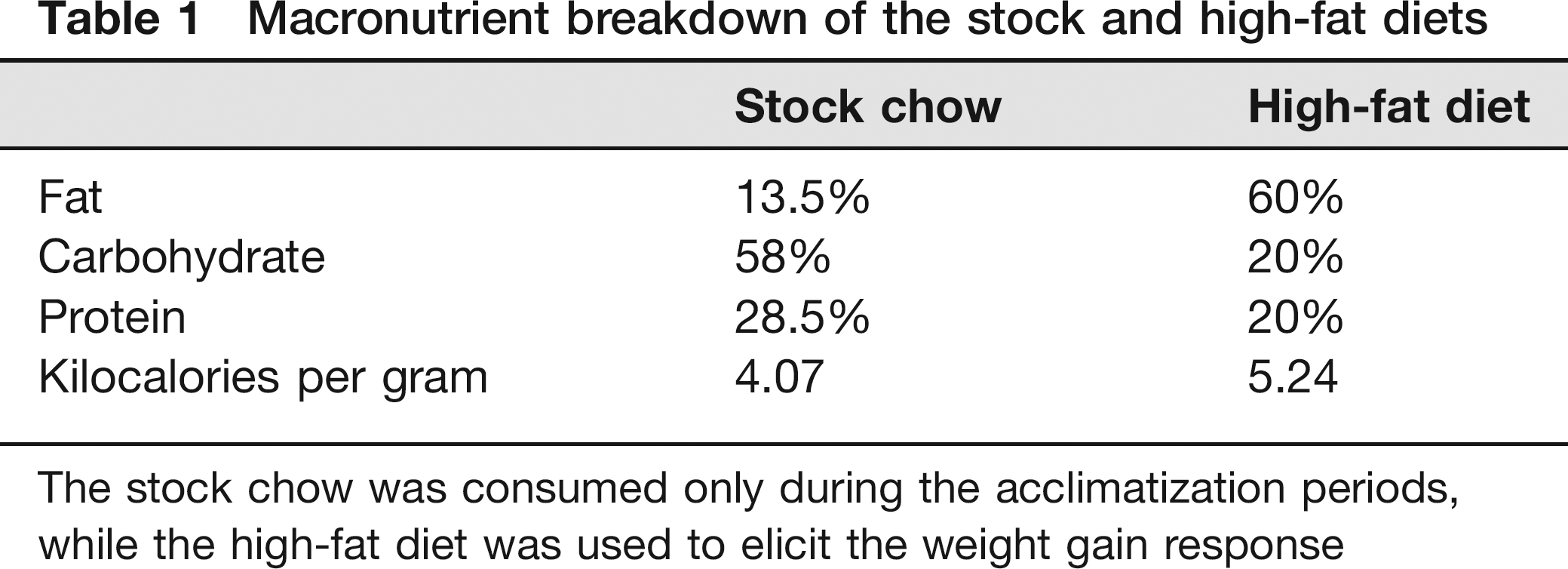
The stock chow was consumed only during the acclimatization periods, while the high-fat diet was used to elicit the weight gain response
Body weight and food intake
Body weight and food intake were measured on a weekly basis using a digital scale. Body weight was reported as a percent change relative to baseline and as an absolute change. Food intake was reported as total caloric intake. During weekly food intake measurement, any food found in the bottom of the cage was collected and weighed along with the food remaining in the food rack.
Experiment 1: effect of individual versus group housing
Mice were housed either individually (HF-I, n = 20 mice) or in groups of four mice per cage (HF-G, n = 12 mice). Both groups were monitored over a period of 12 weeks and consumed an ad libitum high-fat diet for the duration of the experiment (Figure 1). All mice were 6–8 weeks old at the start of feeding and 18–20 weeks old at week 12.
Testing timelines for (a) Experiment 1, (b) Experiment 2 and (c) Experiment 3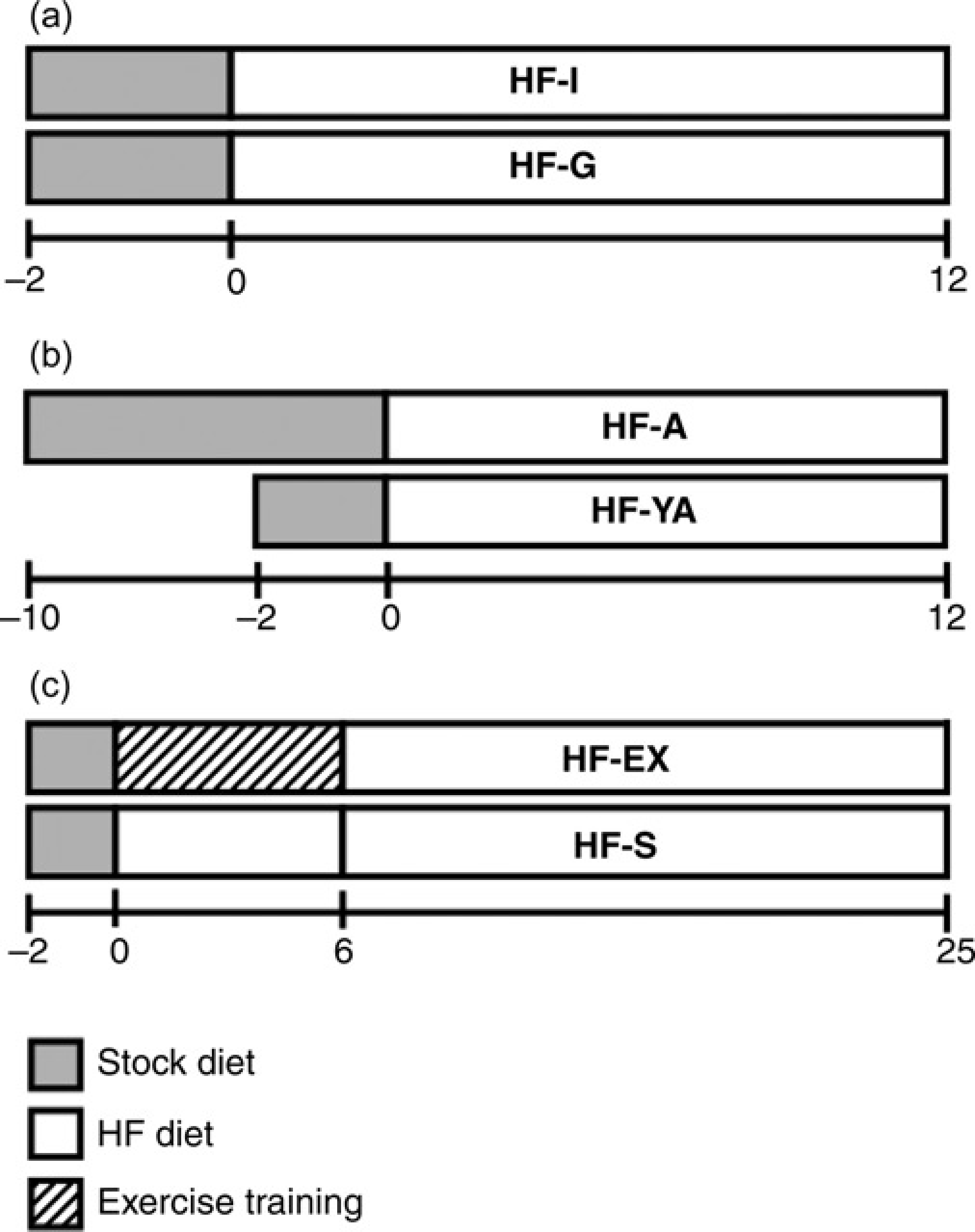
Experiment 2: effect of high-fat feeding during young adulthood versus adulthood
Following a 10-week acclimatization period, mice 19–22 weeks of age were switched from the stock diet to a high-fat diet and monitored weekly for an additional 12 weeks (HF-A, n = 12 mice). Grain-based (stock) and purified ingredient diets have other differences beyond simply macronutrient level and these differences may be proble-matic; however, since stock diets were only used to age the mice, we do not anticipate that this is a major confounder. All mice were 31–34 weeks of age at week 12. This group was compared with a group that began the 12 weeks of high-fat feeding at 8–10 weeks of age (HF-YA, n = 12 mice) making them 20–22 weeks of age at week 12 (Figure 1).
Experiment 3: effect of exercise detraining
Mice were randomly assigned to one of the following two groups (n = 12 mice): high-fat sedentary (HF-S) or high-fat exercise (HF-EX). HF-EX completed five sessions of forced treadmill running per week, while HF-S remained sedentary. Each exercise session consisted of up to one hour of treadmill running (Columbus Instruments, Columbus, OH, USA) at ~21–22 m/min and 1% grade. The mice were exercised in the same order every day to minimize diurnal effects. When mice were unwilling to maintain the prescribed running pace, they were gently pushed by a hand. If mice appeared fatigued or unable to continue, they were removed from the treadmill and the session time was recorded. Exercise sessions were completed in the same room where the animals were housed so that all animals were exposed to noise associated with the treadmill exercise. All exercise sessions were completed within 3 h after the start of the animals’ dark cycle (11:00–14:00). The average weekly running time for HF-EX was 3.5 h, with the group running an average distance of 35.2 km over the course of the entire six-week intervention. Both groups were provided ad libitum access to a high-fat diet. The exercise intervention lasted for six weeks. During weeks 7–25, both groups remained sedentary and continued ad libitum consumption of the high-fat diet (Figure 1). All mice were 8–10 weeks old at the start of feeding and 34–36 weeks old at week 25.
Statistical analysis
Prior to formal statistical analysis, data were assessed for normality and variance using the explore function in SPSS (v.17.0, Chicago, IL, USA). Non-normal data were log-trans-formed prior to statistical testing. For Experiment 1, data were analysed using a 2 (group: HF-I or HF-G) x 4 (time: 0, 4, 8, 12) analysis of variance (ANOVA) with repeated measures on the second factor. For Experiment 2, data were analysed using a 2 (group: HF-YA or HF-A) x 4 (time: 0, 4, 8, 12) ANOVA with repeated measures on the second factor. For Experiment 3, data were analysed using a 2 (group: HF-S or HF-EX) x 19 (time: weekly measurements for week 7–25) with repeated measures on the second factor. Significance was set at P < 0.05 for all measurements. When significance was found, the actual P value was reported. Location of significant differences was determined using separate Student's t-tests with a Bonferroni correction for multiple comparisons.
After ANOVA testing, data from Experiments 1 and 3 were combined and linear regression was used to test models to predict the number of weeks of high-fat feeding needed to reach a specific body weight based on various experimental design considerations. Housing type was dummy-coded for inclusion in the models, with single housing as the referent group. Treatment time measured in weeks was used as an independent factor, along with initial body weight, housing type and age as factors in the models. Separate models were completed for each combination of independent variables to determine the unique statistical improvement that each provided. Improvement in model prediction was determined by testing the R 2 change. Significance for linear regression R 2 change was set at P < 0.05.
Results
Effect of individual versus group housing
A main effect for time existed, in that both groups experienced a significant increase in body weight over the 12-week study period (F = 195.27, P < 0.001). Specifically, HF-G gained 80%, while HF-I gained 62% of their baseline body weight, respectively. An interaction effect for absolute body weight existed only at baseline (F = 8.03, P = 0.002). Due to the difference between groups at baseline, body weight gain was compared by using percent gain from baseline. No significant differences existed between groups in percent weight gain (Figure 2). There was an interaction effect for caloric intake, with significant differences between the groups at baseline, week 8 and week 12 (F = 349.09, P > 0.001). Specifically, caloric intake of HF-I was 84% higher than HF-G at baseline, 76% higher at week 12 and 45% lower at week 8 (Table 2).
Weekly energy intake (kJ): Experiments 1 and 2
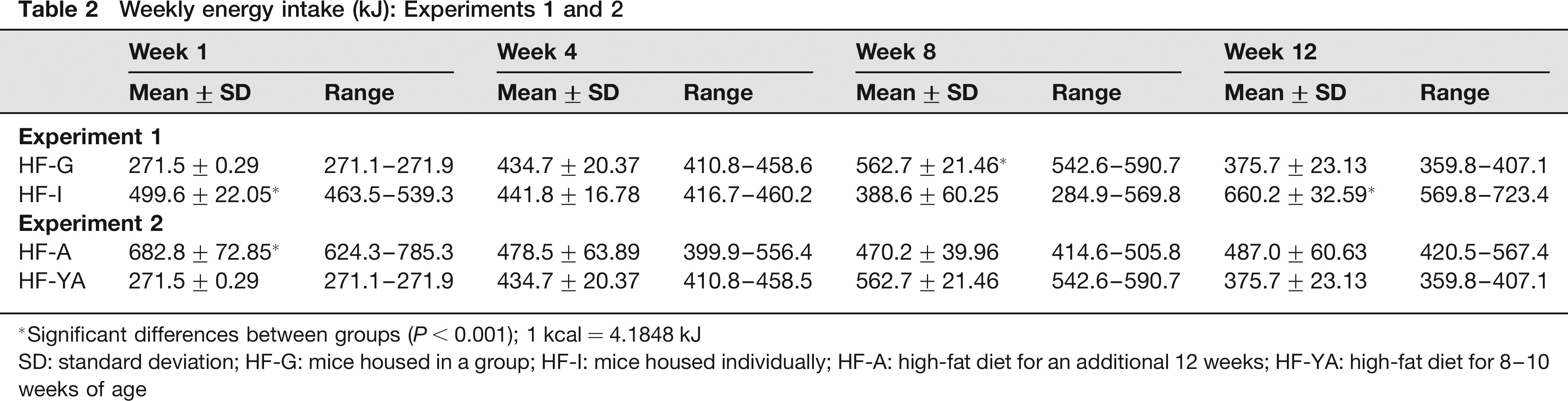
Significant differences between groups (P < 0.001); 1 kcal = 4.1848 kJ
SD: standard deviation; HF-G: mice housed in a group; HF-I: mice housed individually; HF-A: high-fat diet for an additional 12 weeks; HF-YA: high-fat diet for 8–10 weeks of age
Body weights are presented as both absolute gain (top) and percent gain from baseline (bottom) for (a) Experiment 1 : housing type, (b) Experiment 2: age and (c) Experiment 3: previous exercise training. Values are presented as means ± standard error. *Significant difference between groups. **Significant difference from baseline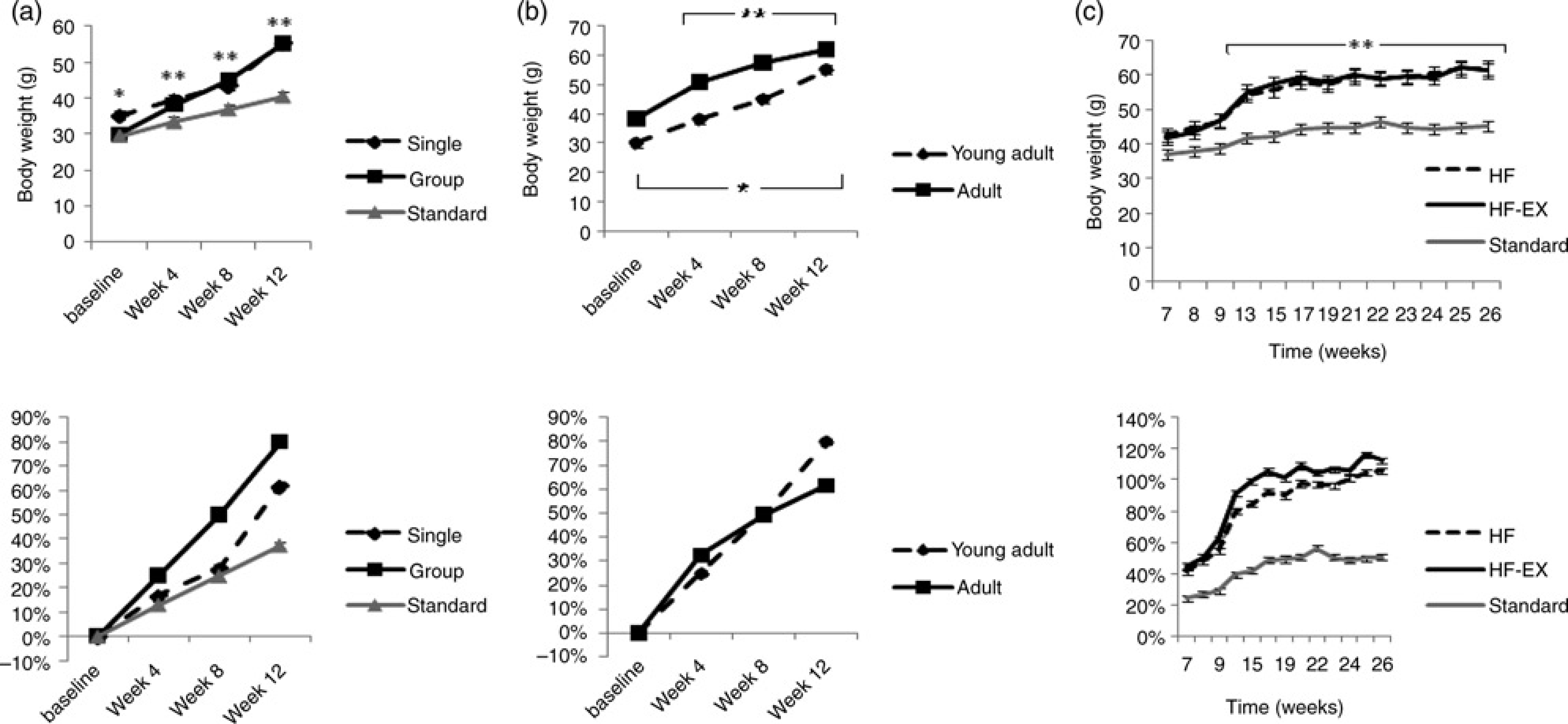
Effect of high-fat feeding during young adulthood versus adulthood
Over the 12 weeks of weight gain, both groups experienced a main effect for time, with HF-YA gaining 82% and HF-A gaining 63% body weight (F = 386.36, P < 0.001). When expressing absolute body weight, we found a significant interaction effect (F = 9.05, P < 0.001). Since HF-A weighed significantly more than HF-YA due to normal ageing, body weight was expressed as a percent change from baseline and the effect was no longer significant (Figure 2). A significant interaction effect for caloric intake existed only at baseline, with HF-A consuming approximately 418.4 kJ or 100 kcal more than HF-YA (F = 359.74, P > 0.001). No significant differences in weekly caloric intake existed between groups at any other time point (Table 2). Given that both adult groups consumed a stock diet for a period of 10 weeks prior to high- or low-fat feeding, this may have altered our outcome measures for this experiment.
Effect of exercise detraining
After the termination of the six-week treadmill running treatment (the detraining period; weeks 7–26), both HF-S and HF-EX experienced a significant increase in body weight (F = 12.95, P > 0.001). HF-S increased by 45% from week 7 and HF-EX increased by 47%. No significant difference in absolute body weight or percent weight gain from baseline existed between groups at any time point throughout the study (Figure 2); however, the average weekly caloric intake of HF-EX was 10% greater than that of HF-S throughout the detraining period (P = 0.021, F = 4.07). Mice in the HF-EX group consistently consumed an average of 46 kJ or 11 kcal more per week than mice in the HF-S group (Table 3).
Weekly energy intake during detraining period: Experiment 3
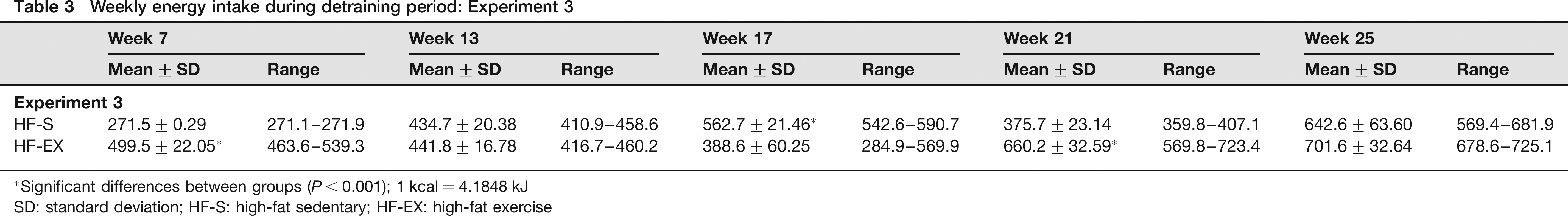
Significant differences between groups (P < 0.001); 1 kcal = 4.1848 kJ
SD: standard deviation; HF-S: high-fat sedentary; HF-EX: high-fat exercise
Linear model for predicting weight gain
The purpose of our linear model approach was to generate a model to predict the weeks of feeding needed to reach a target body weight based on stepwise inclusion of specific variables. A total of four models were analysed to test the impact of each independent variable (Table 4). The inclusion of each of the four independent variables resulted in a significant change in R2. To calculate the weeks of feeding needed, the researcher can enter the target body weight, initial body weight, housing type (0 = single, 1 = group) and age in weeks into the following regression equation:

Linear regression models
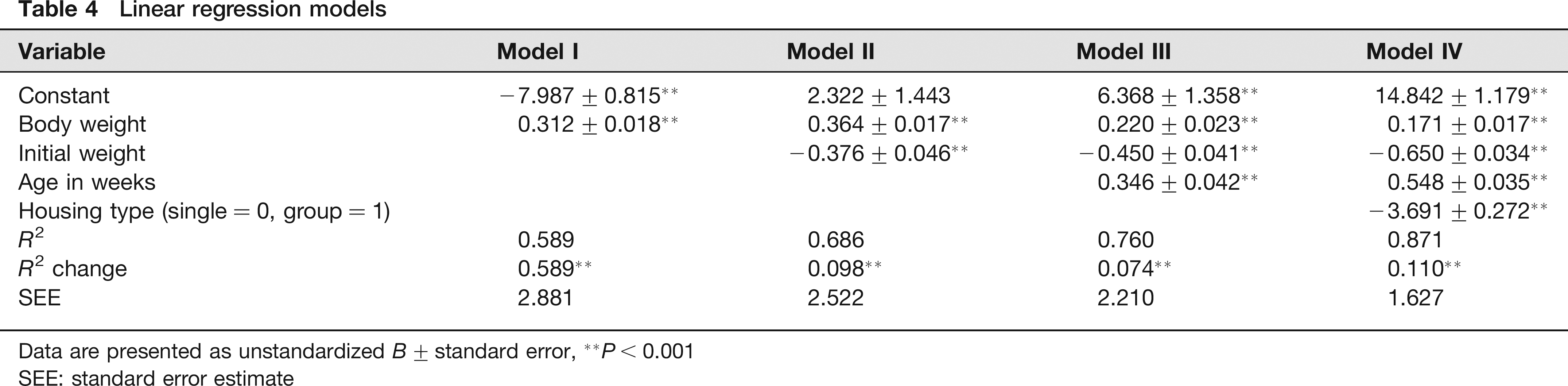
Data are presented as unstandardized B ± standard error,
P < 0.001
SEE: standard error estimate
Discussion
The purpose of this study was to examine experimental design factors that may affect body weight gain during high-fat feeding in CD-1 mice. The key finding of this study is that previous exercise training significantly and negatively influenced food utilization efficiency in response to high-fat feeding in CD-1 male mice. Specifically, mice that had completed a six-week treadmill running protocol consumed more than sedentary mice while maintaining similar body weights. Conversely, housing type and biological factors had no effect on weight gain. We also developed a model that can be used to predict the amount of weeks of high-fat feeding that are needed in order to reach a target body weight while accounting for different experimental design considerations.
A six-week treadmill running pretreatment appeared to increase food utilization efficiency in CD-1 mice consuming a high-fat diet. There were no significant differences in body weight between HF-EX and HF-S, despite the caloric intake of HF-EX being significantly greater throughout the detraining period. The HF-EX group consumed an average of 46 kJ or 11 kcal more per week. Consuming 46 kJ more per week over the course of the detraining period (19 weeks) should have resulted in an additional 25 g of body weight. Prior exercise training appeared to have a persistent effect, increasing feed efficiency and attenuating the excess weight gain. Patterson et al. 8 found a similar effect in a study showing that three weeks of aerobic exercise in young rats prevented excessive weight gain for 10 weeks following exercise cessation. In this study, rats gained 14% less weight than their age-matched, sedentary counterparts while consuming the same amount of calories. A possible explanation for the discrepancy in caloric intake and weight gain between groups in these two studies could be a change in metabolic factors. Previous researchers, including Patterson, have found an increase in thermogenesis with exercise training.8,9 This metabolic effect may persist after exercise cessation, which would account for excess calories consumed by our detrained mice.
Housing conditions did not affect weight gain in CD-1 mice consuming a high-fat diet. Our results are consistent with a previous study by Bibancos et al. 10 who found that six weeks of social isolation did not alter body weight gain. Similar to our design, Bibanco's mice were housed either in groups or singly, with singly-housed mice maintaining auditory and olfactory contact with the other mice. These results contradict those of other studies, which report stress-related suppression of weight gain due to social isolation.11,12 Guo et al. 11 found that long-term (16 weeks) social isolation is associated with decreased weight gain compared with control mice housed in groups. Similarly, Nonogaki et al. 12 found suppressed weight gain in individually-housed mice using a fasting/re-feeding model. It is important to note that these previous studies do not always compare well with the present study because we used a high-fat diet, whereas they used a standard, low-fat diet. It has been previously shown that when mice are presented a highly palatable high-fat diet, food intake and the resulting caloric intake significantly increase. 13 In a separate study, Nonogaki et al. 14 found this to be true for individually-housed mice, as well. While caloric intake was significantly different between groups in the present study, the values were highly variable from week to week. Individually-housed mice tended to consume more kilocalories than group-housed mice. While not measured in the present study, previous research has shown that individually-housed mice are more active in their cages, possibly in an effort to generate warmth in the absence of other mice, 10 which may have accounted for the extra energy consumption. Overall, cumulative caloric intake over the course of our 12-week experiments was similar between individual and group-housed mice eating a high-fat diet. Another plausible explanation for discrepancy between the present study and previous work may be related to variability in weight gain patterns between mouse strains. For instance, C57BL/6J and BALB/cA mice have been shown to be more susceptible to diet-induced weight gain than others; however, both of these are inbred strains of mice.15,16 We selected an outbred stock, since they are generally thought to be better models for longitudinal research and may be more representative of the genetic variation seen in human populations. 7 One limitation of the present study is that we only examined CD-1 mice. Future research should endeavour to compare and contrast weight gain patterns among different mouse strains.
Age at the beginning of a high-fat diet treatment did influence body weight gain in CD-1 mice. The absolute body weight was greater in the young adult group at all time points, but this difference can be attributed to normal, age-related growth. Over 12 weeks of high-fat feeding, both adolescent (2 months) and young adult (4 months) mice gained body weight at a similar rate. As seen in the housing comparison, a significant difference in caloric intake existed at multiple time points throughout the study. When comparing cumulative caloric intake, the groups were similar, and the difference in cumulative caloric intake between groups would not result in a clinically significant amount of weight gain. Wu et al. 17 found age to be a significant modulator of body weight gain when comparing young (2 months) and old (19 months) mice. Similarly, Nishikawa et al. 15 found that middle-aged mice (12 months) were more prone to diet-induced obesity than their young (1 month) counterparts. While both of our groups were under nine months of age at the completion of the study, Wu and Nishkawa's older groups were at least 12 months old at the beginning of high-fat feeding, suggesting that a larger age difference is needed in order to detect an age-effect for weight gain. This age-related difference in weight gain during high-fat feeding is caused by age-associated declines in resting metabolic rate. 18 In mice with only a two-month age difference, no significant effect of age on body weight gain during high-fat feeding appears to exist.
The linear prediction model we have developed will allow investigators to calculate the weeks of high-fat feeding needed to reach a target body weight. In order to develop the most comprehensive and complete model, we included housing type, baseline body weight and weeks of age as factors in the models; each of these factors significantly increased the predictive capacity of the model. With an R value of 0.93, our final model that includes each of these experimental design factors is a strong prediction model that will be a useful tool for new researchers. The model that we developed did not factor in previous exercise training despite its significance to feed efficiency. The main reason that we choose not to include previous exercise training is that the present investigation used a very specific intervention and as such the model would only be applicable if the exact same exercise training protocol was used. Future research is needed to compare and contrast all of the variations in exercise training protocols that are used.
In summary, the present study was designed to investigate how housing conditions, previous exercise training and age influenced body weight gain during a period of high-fat feeding in CD-1 male mice. We have determined that, in our facility, previous exercise history, but not age or housing condition, affects weight gain induced by high-fat feeding, such that previously trained mice consumed more calories while maintaining body weights comparable to sedentary counterparts. The prediction model we developed will allow researchers working with CD-1 male mice to predict quickly the amount of time needed to reach a target body weight, facilitating the use of mouse weight gain models in obesity research. We acknowledge that different sex, strains and diet treatments may yield different results, and future research will explore a wider array of mouse models to further expand our prediction model. Studies similar to the present study are meant to provide a guide to allow researchers to use mouse models more effectively.
Footnotes
Acknowledgements
This study was funded by a TACSM Student Development Research Award to Lisa Esposito, University of Houston Health and Human Performance Department Graduate Student Research Awards to Kelley Strohacker and Lisa Esposito, and a University of Houston College of Education Faculty Development Research Grant to Dr Brian K McFarlin.