
Abstract
Low birth weight in humans is predictive of hypertension in adult life, and while the mechanisms underlying this link remain unknown, fetal overexposure to glucocorticoids has been implicated. We have previously shown that prenatal dexamethasone (DEX) exposure in the rat lowers birth weight and programmes adult hypertension. This current study aimed to unravel the molecular nature of this hypertension. However, unknowingly, post hoc investigations revealed that our animals had been subjected to environmental noise stresses from an adjacent construction site, which were sufficient to confound our prenatal DEX-programming experiments. This perinatal stress successfully established low birth weight, hypercorticosteronaemia, insulin resistance, hypertension and hypothalamic–pituitary–adrenal axis dysfunction in vehicle (VEH)-treated offspring, such that the typical distinctions between both treatment groups were ameliorated. The lack of an additional effect on DEX-treated offspring is suggestive of a maximal effect of perinatal stress and glucocorticoids, serving to prevent against the potentially detrimental effects of sustained glucocorticoid hyper-exposure. Finally, this paper serves to inform researchers of the potential detrimental effects of neighbouring construction sites to their experiments.
As research institutions expand and respond to ever-changing building and animal welfare regulations, they are required to undergo either new construction and/or renovation. However, such structural changes are associated with a plethora of nuisances, such as noise and vibration, with each disturbance being capable of powerfully stimulating the hypothalamic–pituitary–adrenal (HPA) axis. 1 The auditory system is permanently open – even during sleep. Its rapid and overshooting excitations in response to noise signals are subcortically connected, via the amygdala, to the HPA axis, resulting in corticotrophin releasing hormone and adrenocorticotrophic hormone release. 2 Animal experiments show noise-induced changes in the sensitivity of glucocorticoid receptor (GR) by increase of heat-shock proteins 3 and ultrastructural changes of the adrenal gland. 4 Increased Cortisol levels have been found in humans when exposed to aircraft 5 or road traffic noise, 6 even during sleep, 7 implying these effects are mainly without mental control. Of course, increased glucocorticoid and sympathetic neural secretion is a perfect short-term stress response, coordinating appropriate metabolic and vascular changes, and thereby assisting the individual to negotiate the stressor. However, over prolonged time periods, such as persistent noise-induced stress responses, can be gravely damaging to health.2,8
Numerous animal studies have documented the programming effects of pre- and postnatal stress on offspring physiology and behaviour, which are remarkably analogous to those induced by fetal glucocorticoid overexposure. Exposing pregnant dams to stressful stimuli results in both maternal and fetal HPA activation.9,10 Moreover, these offspring display an activated HPA axis till weaning, as adults are more anxious and stress-responsive.10,11 These adverse behavioural effects are associated with altered patterns of brain GR expression, as well as alterations in the circadian rhythm of corticosterone secretion, 11 consistent with HPA axis dysregulation. Similarly, prenatal dexamethasone (DEX) treatment results in permanent lifelong hyperactivation of the HPA axis, 12 also associated with altered brain GR expression and increased anxiety-related behaviour. 13 The sympathetic nervous system (SNS) further participates in stress responses, even in fetal life, 14 and prenatal exposure to various stressors affects the development of sympathetic innervation and/or its regulation. 15 One study on the offspring of rats kept in hypoxia during pregnancy showed altered development of sympathetic centres involved in blood pressure regulation, which continued to adversely affect offspring blood pressure throughout adulthood. 16 In line with these observations, we found that prenatal DEX-treated offspring display hypersensitive pressor responses to even the mildest stressor (e.g. being weighed), and that the resulting stress-induced hypertension is mediated by altered sympathetic nerve responsivity. 15 Postnatal manipulations, such as handling newborn pups, also induce modified stress responses, by reducing anxiety through altered HPA feedback sensitivity and hippocampal GR expression. 17
Here we report the phenotypes of offspring treated with either vehicle (VEH) or DEX during the last week of gestation, which were unintentionally further subjected to perinatal stress, as a result of adjacent construction work. The facility in which our animals are housed was refurbished during this time, and was further located next to a newly constructed wing of a large district general hospital.
Materials and Methods
Animals
Adult Wistar rats (200–250 g; Harlan, Bicester, UK, catalogue number Hsd:Wl), housed in open-top cages, were maintained under conditions of controlled lighting (lights on 07:00 to 19:00) and temperature (22°C), and allowed ad libitum access to food (CRM(E) rat and mouse breeder and grower expanded diet: 61.9% carbohydrate, 18.8% protein, 3.4% oil, 0.6% NaCl; SDS, Wiltham, UK) and non-treated tap water. All experiments were performed in accordance with the UK Animals (Scientific Procedures) Act, 1986.
Prenatal treatments
Virgin female rats were housed with adult male rats. Pregnancy was confirmed by the presence of a vaginal plug, checked every morning. Pregnant females were housed singly and randomly assigned to one of two treatment groups (n = 9–10/group). DEX (100 μg/kg/day, dissolved in 4% ethanol-0.9% saline, 200 μg/mL; Sigma-Aldrich, Poole, UK) was given subcutaneously during the final week of gestation (embryonic days 14–21 inclusive). A control group received VEH injections during the same time period.
Litters
Qn the day of birth (postnatal day 1), litters were weighed, sexed and culled to eight pups per litter, retaining equal numbers of male and female pups where possible. Litters were then left undisturbed until weaning (postnatal day 21), apart from routine weekly maintenance. After weaning, male and female pups from each litter were housed in single-sex groups of two to four and left undisturbed until the time of testing.
Blood sampling
Blood samples taken from the tail vein of conscious rats wrapped in a folded tea towel on the investigator's lap took less than 2 min to complete after removal of the rat from its home cage. Applying thumb and index finger pressure to the proximal part of the tail, a scalpel blade was used to make a nick incision in the distal part of the tail vein to initiate blood flow. The tail was subsequently gently milked to ascertain the sample. Blood was collected in Microvette tubes (Sarstedt, Leicester, UK) and stored on ice until centrifugation at 4°C. Subsequently, all the plasma was stored at - 80°C until assayed.
Plasma hormonal assays
Plasma corticosterone was analysed with the use of an in-house specific radioimmunoassay as described previously 18 and modified for microtitre plate scintillation proximity assay (Amersham Pharmacia Biotech, Buckinghamshire, UK). Inter- and intra-assay variations were <10%.
HPA axis activity
To assess circadian variation in plasma corticosterone concentrations, rats were handled daily for a minimum of 15 min by the same investigator for two weeks before experiments. Initially they were allowed to roam freely on the investigator's lap while being stroked by the investigator, before being habituated to being held in a folded tea towel. Blood samples were taken at 08:00 and 20:00.
Oral glucose tolerance test
Offspring underwent an oral glucose tolerance test (OGTT) at six months of age. 19 Animals were fasted overnight, and testing commenced between 08:30 and 09:00 the following morning. An oral glucose load of 2 g/kg was given by gavage, and blood samples were collected at 0, 30 and 120 min. Glucose was determined by the enzymatic (hexokinase) method using a kit (Sigma, Poole, UK). Inter- and intra-assay variations were <2%. Insulin was measured by enzyme-linked immunosorbent assay (Ultrasensitive Rat Insulin ELISA kit; Crystal Chem, Chicago, IL, USA). Inter-and intra-assay variations were <10%.
Phosphoenolpyruvate carboxykinase enzyme activity
At weaning (postnatal day 21), fed animals were killed by decapitation between 09:00 and 10:00, and livers were removed immediately. Phosphoenolpyruvate carboxykinase (PEPCK) activity was measured as previously described. 20
Blood pressure measurement
Animals between two and three months of age were handled daily and accustomed to the measurement routine for one week before commencement of the experiment. Systolic blood pressure was measured by either an automated tail cuff-plethysmography method or carotid cannulation as previously described.21,22
Statistical analyses
All data are expressed as means ± SEM. Data were compared using one-way or multiple analysis of variance (ANOVA) followed by a least significant difference post hoc multiple comparisons test or unpaired Student's t-tests, where appropriate. Values were considered significant when P < 0.05.
Results
Birth phenotype and catch-up growth
As expected, DEX administration throughout the final week of gestation resulted in a significant decrease in birth weight, i.e. VEH 5.9 ± 0.05 g, DEX 5.4 ± 0.05 g (n = 67 for both groups, P < 0.01 by Student's t-test), representing a mean 8.5% reduction in birth weight as a result of prenatal DEX treatment. No differences in gestation length (VEH and DEX; 22 ± 0 days, n = 8 per group) or litter size (VEH; 9 ± 1 pups versus DEX; 8 ± 1 pups, n = 67 per group) were noted. Figure 1 reveals that male and female offspring from both treatment groups gained weight throughout the course of their postnatal life. Contrary to previous studies, DEX-treated offspring only exhibited catch-up growth until approximately three months of age, thereafter it ceased and they remained approximately 11% lighter than controls for the remainder of their life; for example, at 210 days of age, mean body masses were 660 ± 14 and 591 ± 12 g for VEH- and DEX-treated offspring (n = 34 and 39, respectively, P< 0.05 by Student's t-test).
Growth trajectory of offspring to seven months of age. (a) Growth trajectory of male and female dexamethasone (DEX)- and vehicle (VEH)-treated offspring. Data are presented as mean ± SEM, n = 34 (DEX) and 39 (VEH). (b) Catch-up growth of DEX-treated rats: ratio of mean body weights of DEX versus VEH-treated offspring over the same time period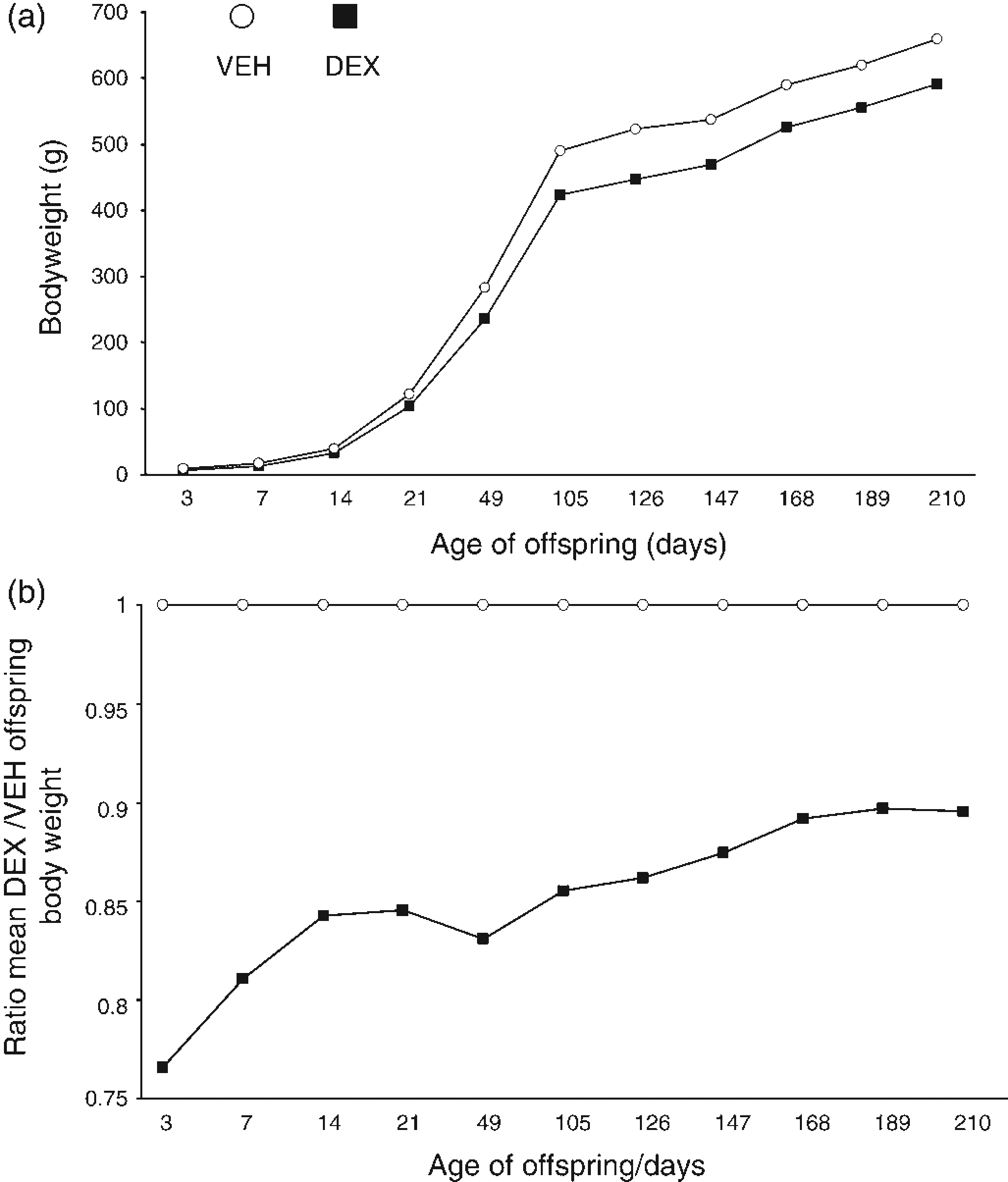
Systolic blood pressure
Although there was a trend for DEX-treated offspring to display elevated systolic blood pressure when measured by carotid cannulation, this did not reach statistical significance in the present cohort (Figure 2). Similarly, no effect of prenatal treatment was revealed by tail-cuff plethysmography at either two or three months of age (Figure 2). However, systolic blood pressure did increase significantly between two and three months of age in both VEH (from 120 ± 7 to 145 ± 5 mmHg, P < 0.05, one-way ANOVA followed by post hoc test) and DEX (from 128 ± 2.5 to 146 ± 5.5 mmHg, P < 0.05, one-way ANOVA followed by post hoc test) treated offspring.
Offspring systolic blood pressure at two and three months of age. Systolic blood pressure of vehicle (VEH) and dexamethasone (DEX)-treated male offspring at two and three months of age, measured by carotid cannulation (average of 2 consecutive days; readings were taken within the first 10 min after connection of the animal to the apparatus) or tail-cuff plethysmography (average of measurements recorded on the last 3 days of 7 days consecutive measurements). Data are presented as mean ± SEM, n = 4–6 per group for carotid cannulation and n = 10 per group for tail-cuff plethysmography, one-way analysis of variance followed by post hoc test, *P < 0.05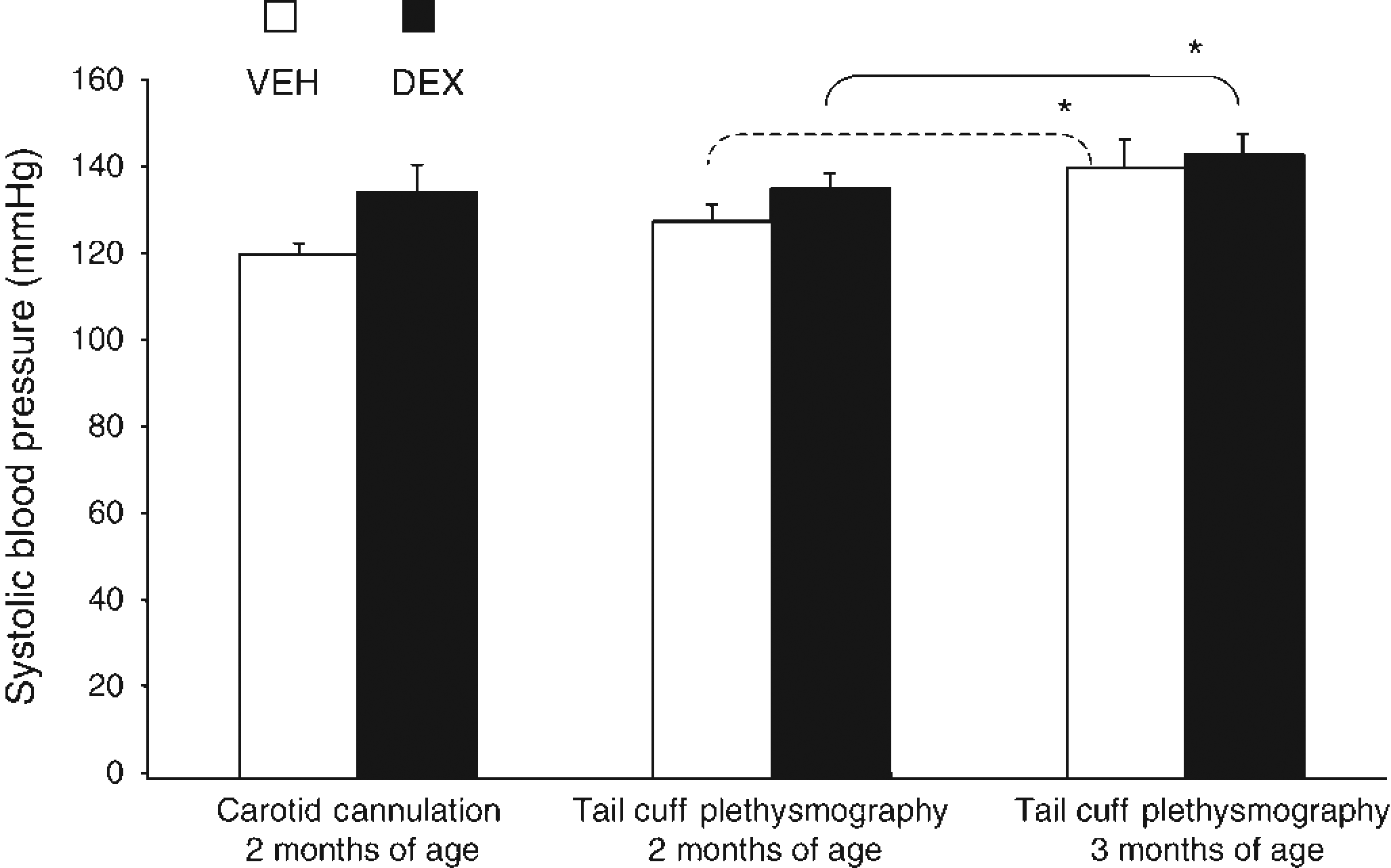
Prenatal DEX, hepatic PEPCK activity and glucose homeostasis
Contrary to previous studies, no difference was observed between the prenatal treatment groups on hepatic PEPCK activity (Figure 3a) at three months of age. This was further associated with a lack of effect of prenatal DEX treatment on either plasma glucose (Figure 3b) or insulin (Figure 3c) during an OGTT, performed at six months of age.
Hepatic phosphoenolpyruvate carboxykinase (PEPCK) activity and plasma glucose and insulin during an oral glucose tolerance test. (a) PEPCK activity measured in microsomal extracts from the livers of three-month-old vehicle (VEH)- and dexamethasone (DEX)-treated offspring. (b) Plasma [glucose] and [insulin] quantified in blood taken at 09:00 from the tail-tip of rats fasted overnight. Plasma glucose before, and at 30 and 120 min after an oral glucose load, measured by hexokinase assay. (c) Plasma insulin before, and at 30 and 120 min after an oral glucose load measured by enzyme-linked immunosorbent assay. All data are presented as mean ± SEM, n = 7–8 per group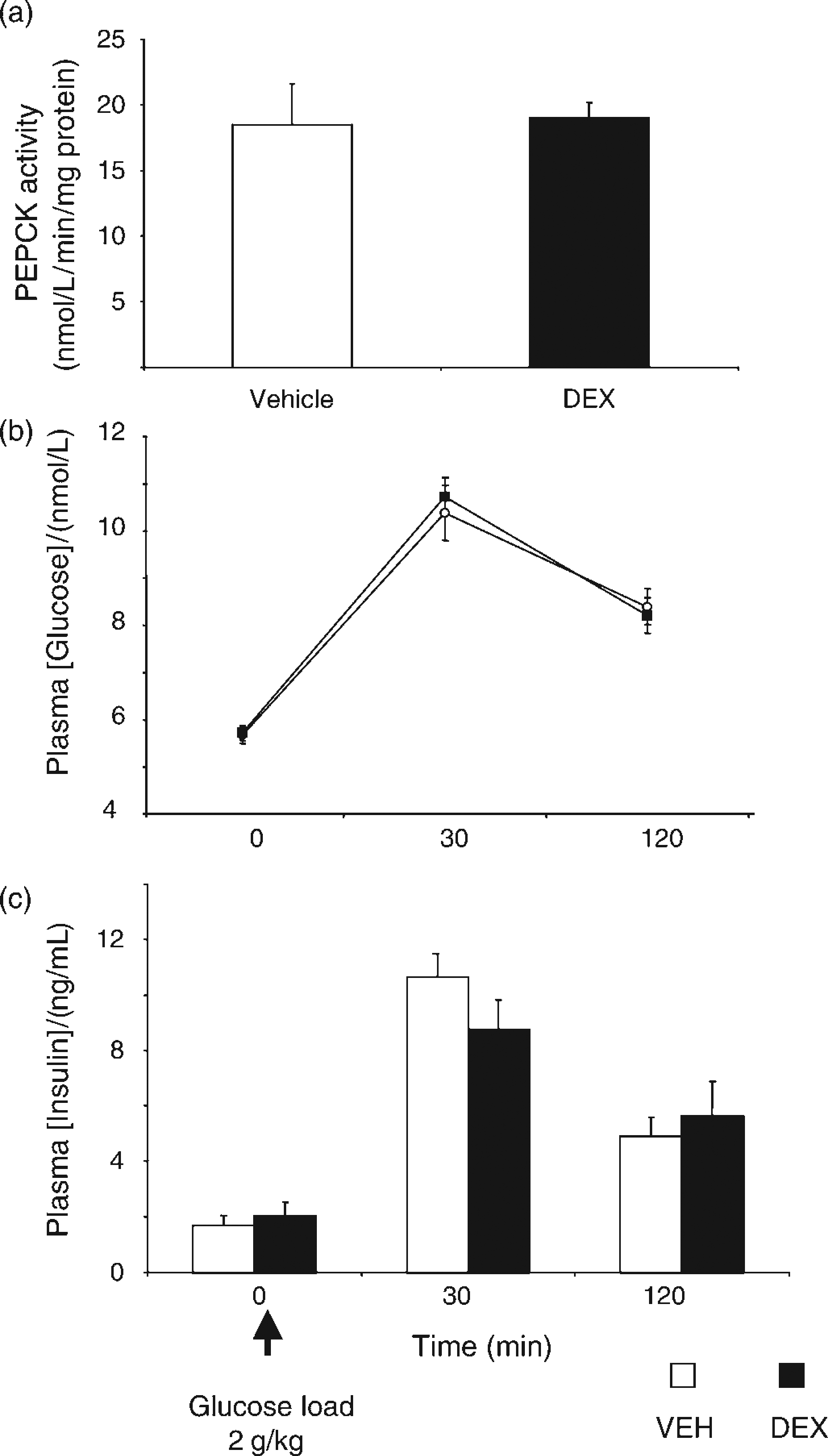
Plasma corticosterone concentrations
At three months of age, both male and female offspring of both prenatal treatment groups displayed greatly elevated increases in basal morning (08:00) plasma CORT (Figure 4), compared with those levels normally observed in our laboratory (Table 1).
Offspring basal plasma corticosterone [CORT] concentrations. Plasma [cort] quantified in blood taken from 08:00 from the tail-tip of male and female offspring. Data are presented as mean ± SEM, n = 7–8 per group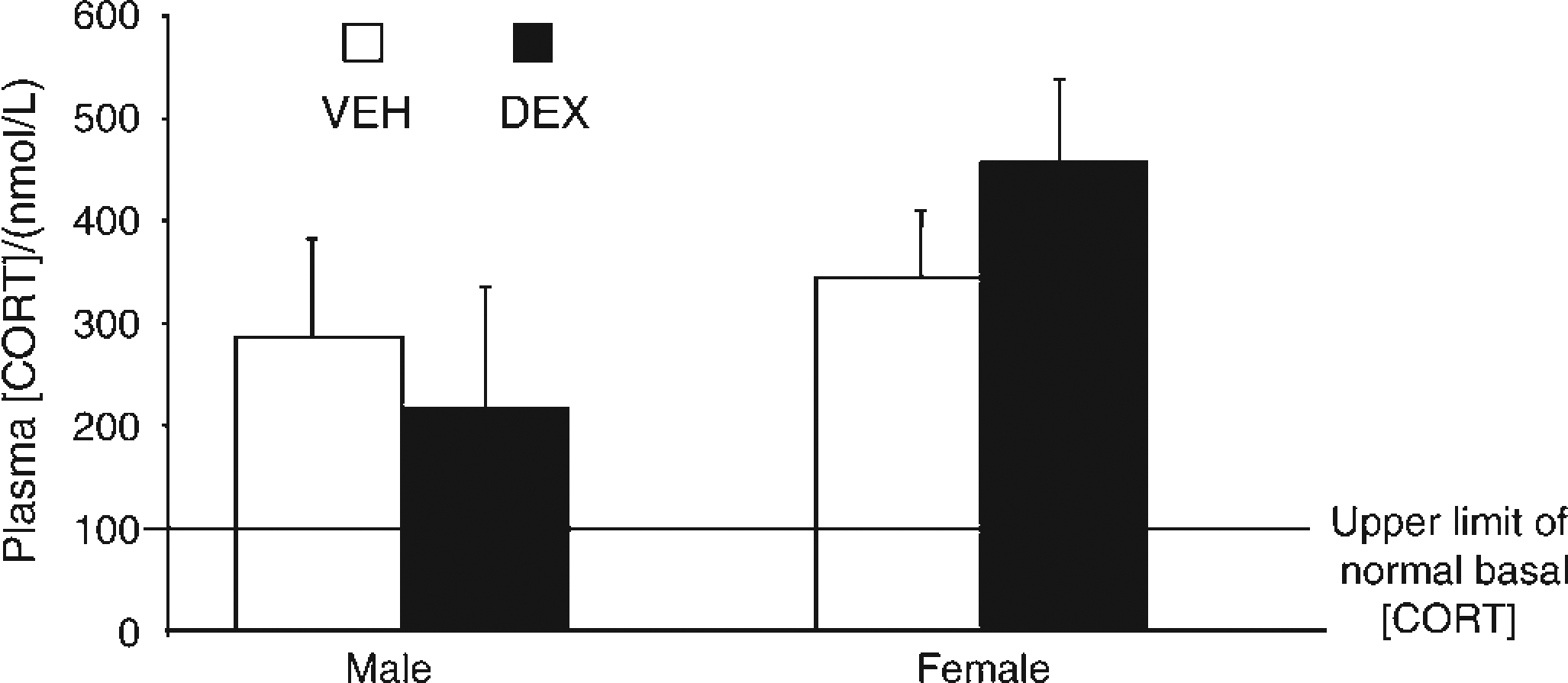
Comparison of prenatally treated offspring phenotypes before, during and after exposure to perinatal environmental stress
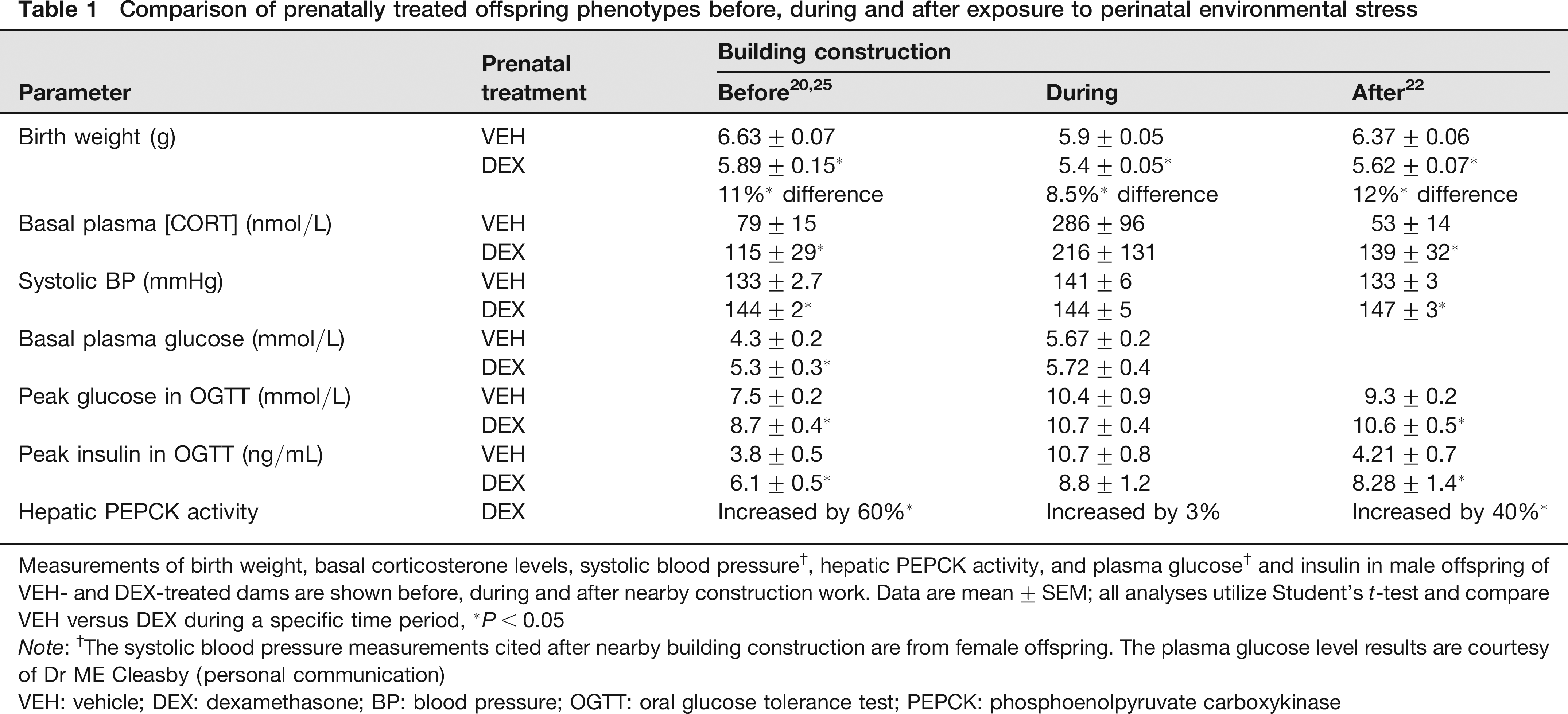
Measurements of birth weight, basal corticosterone levels, systolic blood pressure†, hepatic PEPCK activity, and plasma glucose† and insulin in male offspring of VEH- and DEX-treated dams are shown before, during and after nearby construction work. Data are mean ± SEM; all analyses utilize Student's t-test and compare VEH versus DEX during a specific time period,
P< 0.05
Note: †The systolic blood pressure measurements cited after nearby building construction are from female offspring. The plasma glucose level results are courtesy of Dr ME Cleasby (personal communication)
VEH: vehicle; DEX: dexamethasone; BP: blood pressure; OGTT: oral glucose tolerance test; PEPCK: phosphoenolpyruvate carboxykinase
Detection of environmental disturbance
As each characterizing experiment began to highlight the collapse of our prenatal progamming model, we began an extensive hunt of potential stressors. With no change to animal chow, bedding, water supply or animal carers, we looked to an adjacent construction site as the potential perinatal stressor. During construction of the new hospital wing, corticosterone levels were assayed regularly in non-prenatally manipulated rats. These animals were bred in-house, and revealed persistent hypercorticosteronaemia during the construction, e.g. 08:00 samples: 446.5 ± 158 nmol/L (n = 8) at four weeks and 456.5 ± 142 nmol/L (n = 8) at six weeks during building works.
Plasma corticosterone levels in VEH-treated male offspring, and naïve males after the cessation of adjacent building construction
As demonstrated in Figure 5, VEH-treated offspring, aged eight months revealed significant reductions in their basal corticosterone levels, though they still exceeded the upper limit of the normal. Conversely, naïve animals bought into our research facility had corticosterone levels of 31.5 ± 6 nmol/L (n = 8); values were lying well within the normal basal range for our laboratory. Construction of the neighbouring building was completed at this time of testing.
Corticosterone levels in vehicle (VEH)-treated male offspring and naïve males. Plasma [CORT] quantified in the blood taken from 08:00, from the tail-tip of VEH-treated offspring aged eight months. Shown also are the previous mean [CORT] for these rats when they were three months of age, the upper limit of normal basal [CORT] (represented by a solid black line), and the mean [CORT] measured in naïve males aged three months. Data are presented as mean ± SEM, n = 8 per group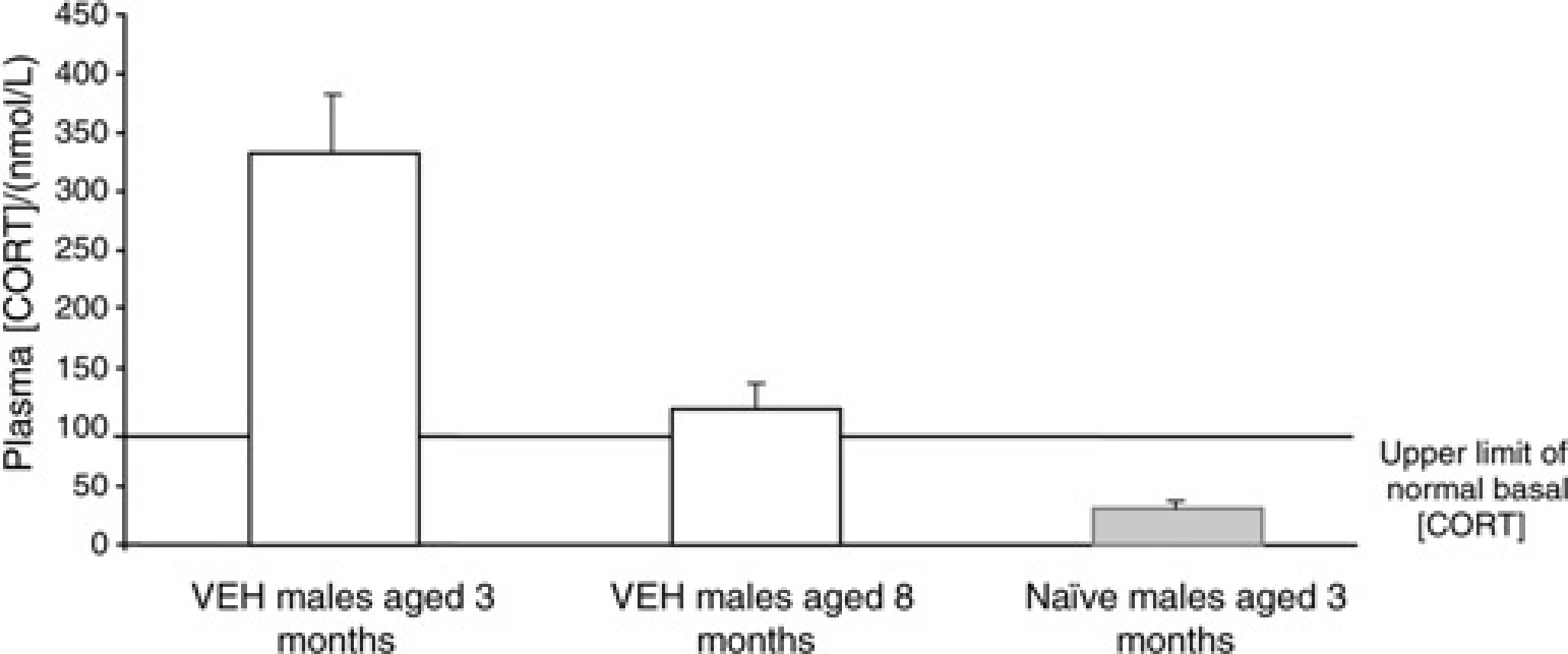
Comparison of offspring phenotypes pre-, during and post-nearby construction
Characterization of this cohort revealed a phenotype that differed substantially from previous and subsequent cohorts of prenatally treated offspring. A summary of the results highlighting the discrepancies between offspring cohorts generated for this study, and those before and after the cessation of building work, is outlined in Table 1.
Discussion
This paper recounts data acquired during a series of investigations intended to unravel the molecular mechanisms underlying the hypertensive phenotype of the DEX-programmed rat. Instead, it reveals how the physiological profile of a prenatal VEH- or DEX-treated animal is confounded by perinatal stress, induced in this case by construction noise. Furthermore, during this same time period the animal facility was frequently but irregularly passed by a plethora of construction lorries and machines accessing the adjacent building site, which would have involved the emission of infrasound. 2 Such environmental disturbances would have been clearly audible to our cohorts, as rodents possess a hearing range that incorporates both ultra- and subsonic frequencies. Moreover, prenatal DEX-treatment programmes greater cochlear sensitivity to noise stress, 23 which is most harmful when emitted irregularly and unpredictably. 24 Thus, these sound frequencies may have induced stress either pre- and/or postnatally to both VEH- and DEX-treated cohorts through the maternal and/or fetal HPA-axis and/or SNS.
Despite in utero growth retardation, the other well-documented adult characteristics of the DEX-programmed phenotype were nonexistent. As highlighted by Table 1, this probably reflects alterations in the VEH-treated offspring that developed a phenotype analogous to that of DEX-offspring, without any appreciable modification in the phenotype of the latter. Given the null effect of perinatal stress on the phenotype of the DEX-treated offspring, it is tempting to speculate that there is an ‘upper limit’ for the effects of prenatal glucocorticoids. Whether this ‘upper limit’ is a result of modified GR and/or MR expression, or is a function of enhanced glucocorticoid metabolism, remains to be established. Alternatively, it is conceivable that a factor exists, which antagonizes the effects of glucocorticoids, thereby limiting their adverse effects in the long term.
Birth weights of both cohorts, but especially of VEH-treated offspring, were significantly reduced when compared with previous25,26 and later15,22 studies, implying that the initiation of environmental stress occurred before birth. Subsequent catch-up growth of DEX-treated offspring also failed to achieve completeness, and they remained on average 11% lighter throughout adulthood, when compared with controls, though the implications of this are controversial. While complete26,27 and lack of catch-up growth28,29 have been demonstrated in several programming models, each reported cohort still developed a ‘programmed’ phenotype. These studies support the present view that birth weight and, therefore, subsequent catch-up growth form relatively crude markers of exposure to an adverse intrauterine environment, rather than underlying a specific cause of pathophysiology in the offspring. In a previous study, 22 birth weight and postnatal catch-up growth were observed in both male and female DEX-treated offspring, yet the adult ‘programmed’ phenotypes differed substantially, consistent with this hypothesis.
At two and three months of age, no differences in systolic blood pressure were detected by either tail-cuff plethysmography or carotid cannulation. This contrasts with previous reports that utilized these techniques to demonstrate hypertension in DEX-treated adult offspring.22,25,30,31 While there was an increase in systolic blood pressure between the time of measurement at two and three months of age, it occurred to the same degree in the offspring of both cohorts, and possibly reflects perinatal stress programming.22,25,30-32 Interestingly, it is the VEH-treated offspring that demonstrate blood pressure values equivalent to those normally associated with prenatal DEX-programming,22,25,31 while the DEX-treated offspring display typically expected values, again suggesting that perinatal noise stress did not add to their phenotype. In terms of mechanism for the hypertension associated with perinatal stress programming, it is most likely mediated through elevated glucocorticoids, 25 altered sympathetic nerve responsivity 15 and/or an activated renin-angiotensinogen system.15,22,32,33 Furthermore, while tail-cuff plethysmography and carotid cannulation are associated with considerable stress artefacts, the nature of their varying effects on both VEH- and DEX-treated offspring does not implicate them as causative factors in the hypertension detected here. 15
In further contrast to previous19,26 and subsequent22,34 findings, prenatal DEX treatment did not affect plasma glucose or insulin levels measured at three time points during an OGTT. However, increases in both these plasma variables following the oral glucose load indicate that nothing went awry with the procedure. 20 In line with birth weight and blood pressure data from the present cohorts, the elevated plasma glucose and insulin concentrations measured in VEH-treated offspring mirrored those typically expected of prenatal DEX-treated offspring. Furthermore, the comparable insulin–glucose profile of these offspring cohorts is consistent with their identical hepatic PEPCK activities. Such elevated levels of activity are normally associated with the hyperglycaemia found exclusively in prenatal DEX-treated offspring.22,26
Highly elevated levels of plasma corticosterone, indicative of stress, were observed in both cohort offspring, and again, unlike previous13,25 and later15,22 findings, no differences between cohorts were detected. During construction, rat corticosterone levels were assayed at regular intervals, serving as environmental stress monitors. These rats, though born in the animal facility, were not subjected to any in utero manipulation. However, they too repeatedly displayed equivalent levels of corticosterone, suggesting that perinatal stress had programmed the HPA axis of each animal born within the facility, which could be further acted upon by construction disturbances. Indeed, following the cessation of building works, plasma corticosterone values fell dramatically, but still remained in excess of the upper normal basal limit, revealing continued lifelong HPA dysfunction. At this time naïve adult males were introduced to the animal facility, and displayed appropriate basal plasma corticosterone levels, suggesting that exposure to the stressors during the perinatal period is critical to programming later dysfunction.
In conclusion, it is evident that perinatal stress induced by environmental disturbance is sufficient to confound prenatal DEX-programming experiments. This perinatal stress successfully introduced low birth weight, hypercorticosteronaemia, insulin resistance and hypertension to VEH-treated offspring, such that the typical distinctions between both treatment groups were completely ameliorated. The lack of an additional effect on DEX-treated offspring is suggestive of an ‘upper limit’ effect of prenatal stress or glucocorticoids, serving to prevent against the potentially detrimental effects of sustained glucocorticoid hyper-exposure. Finally, perinatal stress is further able to programme HPA-axis dysfunction in offspring that were not additionally subjected to any other in utero manipulations.
With the demonstrated ability to modify complex physiological systems, it is quite conceivable that construction work may serve as a major contributor to the reports of ‘biological variability’. As our other studies have illustrated,15,22 under well-controlled conditions, biological variability is actually quite negligible, and those environmental variables, which may go unnoticed by many researchers (particularly those that utilize animal tissues harvested by others), can cause pronounced, misleading, and costly alterations in otherwise robust model systems.
Footnotes
Acknowledgements
Expert animal assistance was provided by Mr Willy Mungall. This study was supported by a Wellcome Trust CVRI studentship (DO'R), the Wellcome Trust (JRS and MCH) and the Medical Research Council (CJK).