Salmonella Typhimurium infections in pigs pose an important human health hazard. One promising control measure is the development of live attenuated vaccine strains using defined knockout mutants. Preferably, screening of candidate knockout vaccine strains for attenuation should first be done in models allowing testing of a large number of strains. Thereafter, a limited number of selected strains should be further characterized in an experimental infection model in pigs. The aim of the present study was to develop such models. The invasive and proliferative characteristics of S. Typhimurium were assessed in both a non-polarized and a polarized porcine intestinal epithelial cell line. Neutrophils obtained from porcine blood were used to study the capacity of Salmonella to withstand killing by these phagocytes. The ability to induce an intestinal inflammatory response was investigated in a terminal intestinal loop model. The systemic phase of infection was mimicked by studying the uptake and intracellular survival of S. Typhimurium in porcine pulmonary alveolar macrophages and peripheral blood monocytes. These models should allow screening for attenuated strains. For further characterization, an experimental infection model was established, providing extensive data on the course of an oral infection and the optimal time points for colonization (day 5 postinoculation [pi]) and persistency (days 21–28 pi) in pigs. In conclusion, screening for virulence of S. Typhimurium strains with subsequent confirmation for a subset of strains in a well-defined experimental infection model would significantly reduce the number of experimental pigs required.
Salmonella enterica serovar Typhimurium (S. Typhimurium) infections in pigs cause considerable economic losses and public health problems, as pigs are a reservoir of infection for humans.1 Until today, measures to control Salmonella infections in pigs are often of hygienic origin with no guarantee for a satisfactory result. One promising measure in the combat against Salmonella is the development of live attenuated vaccine strains using defined knockout mutants.2–4 Previously, virulence of Salmonella strains in pigs was assessed using in vivo infection models.5–9 However, screening candidate knockout vaccine strains for attenuation, solely using in vivo infection models is expensive, time-consuming and requires high numbers of experimental animals.
Most of our knowledge concerning interactions of Salmonella with the host is derived from experiments in mice or murine cell types. Although many tools are available for studying Salmonella pathogenesis in murine models, these results often cannot be reproduced in other hosts, compromising extrapolation to human and pig disease.10
In vitro studies using primary isolated cells or cell lines of porcine origin are rare.11–13 Comparative data from these alternative methods to one another and to those of the in vivo infection model are missing. In times of growing animal welfare awareness, there is need for experimental models that replace, reduce or refine the use of experimental animals.14
It was the aim of this study to evaluate and discuss different in vitro and in vivo models to investigate host–pathogen interactions and virulence of S. Typhimurium in pigs. The intestinal phase of infection was reproduced using an invasion and proliferation assay in two porcine intestinal epithelial cell lines, a bacterial killing assay with neutrophils and a terminal intestinal loop model in pigs. For modelling the systemic phase of the infection, a bacterial killing assay with porcine alveolar macrophages and peripheral blood monocytes (PBM) was used. Finally, the course of a S. Typhimurium infection, different clinical score systems and the effect of different infection doses were studied in an experimental infection model in pigs.
Materials and methods
Animals
For the in vitro experiments with pulmonary alveolar macrophages (PAM), a total of three male crossbred pigs, aged 3–4 weeks, were used. For the in vitro experiments with polymorphonuclear cells (PMN) and PBM one female hybrid pig, aged 20 weeks, was used. The intestinal loop model was performed on two female 6-week-old, farm reared Landrace/Large White cross male piglets. For the in vivo experiments, a total of 38 piglets of mixed sexes (commercial closed line based on Belgian landrace), aged four weeks were used. All the pigs used in the experiments came from farms with no history of Salmonella infections and were tested negative for Salmonella in faecal sampling. Pigs arrived at the facility seven days before infection as an acclimatization period. They were kept in pairs in isolation units at 25°C under natural day–night rhythm with ad libitum access to feed and water in high efficiency particulate air (HEPA)-filtered stables. Piglets were provided with a rubber ball as environmental enrichment.
The experiments were approved by the ethical committee of the Faculty of Veterinary Medicine, Ghent University.
Bacteria
Salmonella serovar Typhimurium strain 112910a, phage type 120/ad, isolated from a pig without clinical signs of salmonellosis, was used in all the experiments. The bacteria were stored at –70°C. The inocula for the intestinal loop model were prepared according to the temperature shift method for Salmonella.15 Cultures in Luria-Bertani broth (LB; Sigma-Aldrich, Steinheim, Germany) were centrifuged at 130 rpm for 24 h at 25°C. After diluting two-fold, the cells were incubated for 2 h at 37°C, by centrifuging at 130 rpm. Later, three syringes of 5 mL were filled with the strain. The actual number of bacteria/millilitre was assessed by plating serial dilutions on MacConkey agar (Oxoid, Hampshire, UK) plates.
For all the other experiments, the bacteria were grown in the brain heart infusion (BHI) broth (BHI; Oxoid) for 6 h at 37°C without centrifugation and then washed twice in phosphate-buffered saline (PBS). The number of viable microorganisms was determined by plating 10-fold dilutions on brilliant green agar (BGA; Oxoid).
Interactions of Salmonella Typhimurium with intestinal epithelial cells
All the cell lines were stored in liquid nitrogen until use.
The porcine epitheloid intestinal cell line IPI-2I was derived from the ileum of an adult boar.16 The polarized porcine epithelial intestinal cell line IPEC-J2 was derived from jejunal epithelia isolated from a neonatal piglet.13,17
The IPI-2I cells were cultured in Roswell Park Memorial Institute medium (RPMI) (Gibco, Life Technologies, Paisley, Scotland, UK) containing 10% (v/v) fetal calf serum (FCS; Hyclone, Cramlington, UK), 2 mmol/L l-glutamine (Gibco, Life Technologies), 1 mmol/L sodium pyruvate (Gibco, Life Technologies), 100 units nystatin/mL (Gibco, Life Technologies), 100 units of penicillin/mL and 100 μg of streptomycin/mL (penicillin/streptomycin, Gibco, Life Technologies). The IPEC-J2 cells were maintained on 50% Dulbecco's modified essential medium (DMEM) – 50% (v/v) F12 medium (Gibco, Life Technologies) with 5% (v/v) FCS, 1% (v/v) insulin/transferrin/Na-selenite media supplement (Gibco, Life Technologies) and antibiotics as described above. IPI-2I and IPEC-J2 cells were seeded in 24-well plates at a density of approximately 105 cells per well and were allowed to grow to confluency for at least seven days. These wells were inoculated with Salmonella at a multiplicity of infection (MOI) of 10:1. To synchronize the infection, the inoculated multiwell plates were centrifuged at 365 × g for 5 min. After 25 min of incubation at 37°C under 5% CO2, the wells were washed and the fresh medium was supplemented with 50 μg/mL of gentamicin (Gibco, Life Technologies) was added. After an additional 60 min of incubation at 37°C under 5% CO2, the wells were washed three times.
To assess invasion, the cells were lysed with 0.25% deoxycholate (Sigma-Aldrich) 90 min after inoculation and 10-fold dilutions were plated on BGA plates.
To assess the intracellular growth of S. Typhimurium in the cells after 24 h of incubation, the medium containing 50 μg/mL of gentamicin was replaced 90 min after inoculation with fresh medium supplemented with 15 μg/mL of gentamicin and the number of viable bacteria was assessed as described above.
All the measurements were performed in triplicate and the experiment was carried out on at least two occasions. The results were analysed by a paired Student's t-test. A P value of <0.05 was considered significant.
Interactions of Salmonella Typhimurium with porcine macrophages and monocytes
PAM were collected by broncho-alveolar washes and PBM by ficoll-paque density gradient centrifugation, as described previously.11,18 The cells were maintained in RPMI supplemented with 10% (v/v) FCS, 0.3 mg/mL l-glutamine, 1 mmol/L Na-pyruvate, 1% (v/v) non-essential amino acids, 10 U/mL heparin, 100 U/mL penicillin, 0.1 mg/mL streptomycin and 0.1 mg/mL kanamycin. After overnight incubation in 96-well plates and washing, the cells were inoculated with S. Typhimurium in RPMI at an MOI of 10. The determination of invasion and intracellular replication of S. Typhimurium was carried out using a gentamicin protection assay as described for the intestinal epithelial cell lines. Cells were lysed at 90 min, 3 h or 7 h postinoculation (pi), using 0.5% (v/v) triton X-100 (Sigma-Aldrich). All the measurements were performed in triplicate and the experiments were carried out on three occasions. The results were analysed by a paired Student's t-test. A P value of <0.05 was considered significant.
Interactions of Salmonella Typhimurium with neutrophils
Neutrophils were isolated from heparinized (1500 IU/mL) blood using discontinuous Percoll (Pharmacia, Uppsala, Sweden) density gradient centrifugation according to Dom et al.19
Determination of the percentage of salmonellae killed by PMN was carried out according to Barrio et al.20 with some modifications. The assay was run in Eppendorf® tubes (Netheler-Hinz GmbH, Hamburg, Germany) in a final volume of 500 μL with the following composition: 400 μL Hank's Balanced Salt Solution (HBSS; Gibco, Life Technologies) containing 2.5 × 106/mL PMN and 100 μL HBSS containing 107 cfu/mL bacteria (mixture sample). Control samples consisted of 400 μL HBSS and 100 μL HBSS with 107 cfu/mL bacteria. The tubes were rotated end-over-end at 37°C for 60 min. A sample (25 μL) was taken at 0 and 60 min pi. PMN in the mixture samples were lysed with 0.5% (v/v) triton X-100. The number of bacteria in the mixture and control samples was determined by plating 10-fold dilutions on BGA plates. The percentages of killed bacteria were calculated using the formula described by Barrio et al.20
Intestinal loop model
Pigs were starved overnight before surgery to empty the intestines. Piglets were sedated for intratracheal intubation using 1 mL/20 kg azaperone (Stressnil, Janssen Pharmaceutics, Beerse, Belgium). Induction of terminal anaesthesia was performed by slow intraveneous injection of 1 mL/4 kg alphaxolone/alphadolone (Saffan, Schering-Plough Animal Health, Middlesex, UK). Anaesthesia was maintained with 1–3% isoflurane in conjunction with 1% pure oxygen, using a closed circuit with CO2 and isoflurane absorbers. The piglets were kept warm and heart and respiratory rate were monitored. A continuous ringer-glucose drip was inserted into the dorsal ear vein.
The porcine ligated loop model was carried out as follows. Commencing at the mid ileum, loops of 7–8 cm were ligated using surgical silk, separated by 1 cm spacers. The lumen of the ileum was flushed with 0.9% (w/v) NaCl before the construction of the loops to remove the intestinal contents. The loops were injected with 1–2 × 109 cfu of S. Typhimurium. Approximately 10 mL of blood was removed from the piglets to isolate the PMN. The isolated PMN were labelled with 111Indium oxinate and re-injected intravenously. The influx of PMN in the intestinal wall and in the lumen, as assessed by the counts per minute (cpm) emitted from 111Indium-labelled PMN within the loop, was recorded 12 h after injection of the loops using a gamma counter before euthanasia of the animals. The results were analysed by a paired Student's t-test. A P value of <0.05 was considered significant.
Course of a Salmonella Typhimurium infection in pigs
Twenty-one pigs were randomly divided into seven groups of three piglets, which were orally inoculated with 107 cfu S. Typhimurium in 2 mL PBS, by syringe. One group of three pigs was sham-inoculated orally with 2 mL PBS and served as negative controls. The rectal temperature and clinical condition (anorexia, lethargy, diarrhoea) were monitored and faecal samples were taken directly from the rectum of each pig on several days pi. On days 1, 2, 5, 9, 14, 21 and 28 pi, three pigs were anaesthetized with an intramuscular injection (0.5 mL/10 kg body weight) of 1:1 mixture of xylazine (Xyl-M 2%, VMD, Arendonk, Belgium) and zolazepam/tiletamine (Zoletil 100, Virbac, Carros, France), after which they were euthanized by intracardial injection of 2 mL T61 and necropsied. Samples of various organs were taken for bacteriological analysis.
All the samples were stored at −70°C until further examination. The samples were thawed and weighed, 10 % (w/v) suspensions were prepared in buffered peptone water (BPW; Oxoid) after which the material was homogenized. The homogenized samples were examined for the number of Salmonella bacteria per gram by plating 10-fold dilutions on BGA. If negative at direct plating, the samples were pre-enriched overnight at 37°C in BPW, enriched overnight at 37°C in tetrathionate broth and then plated on BGA. Samples that were negative after direct plating but positive after enrichment were presumed to have <10 cfu/g. Samples that remained negative were presumed to contain 0 cfu/g.
Dose–response study
Twelve pigs were randomly divided into three groups of four pigs, which were orally inoculated with 105 cfu (group 1), 107 cfu (group 2) or 109 cfu (group 3) of S. Typhimurium in 2 mL PBS. The animals were weighed before the inoculation and just before euthanizing. Their rectal temperature and clinical condition (anorexia, lethargy, diarrhoea) were monitored twice daily the first two days pi and daily at day 3 and day 5 pi. On day 5, the pigs were euthanized and necropsied. Samples were taken for bacteriological analysis and bacterial counts in the tissues and faeces were determined as described above.
Results of bacterial analysis and loss of weight for the different groups were statistically analysed using a Kruskal-Wallis test and significant differences (P < 0.05), were compared by a Mann-Whitney test. A P value of <0.05 was considered significant.
Results
Salmonella Typhimurim invades and replicates in intestinal epithelial cell lines
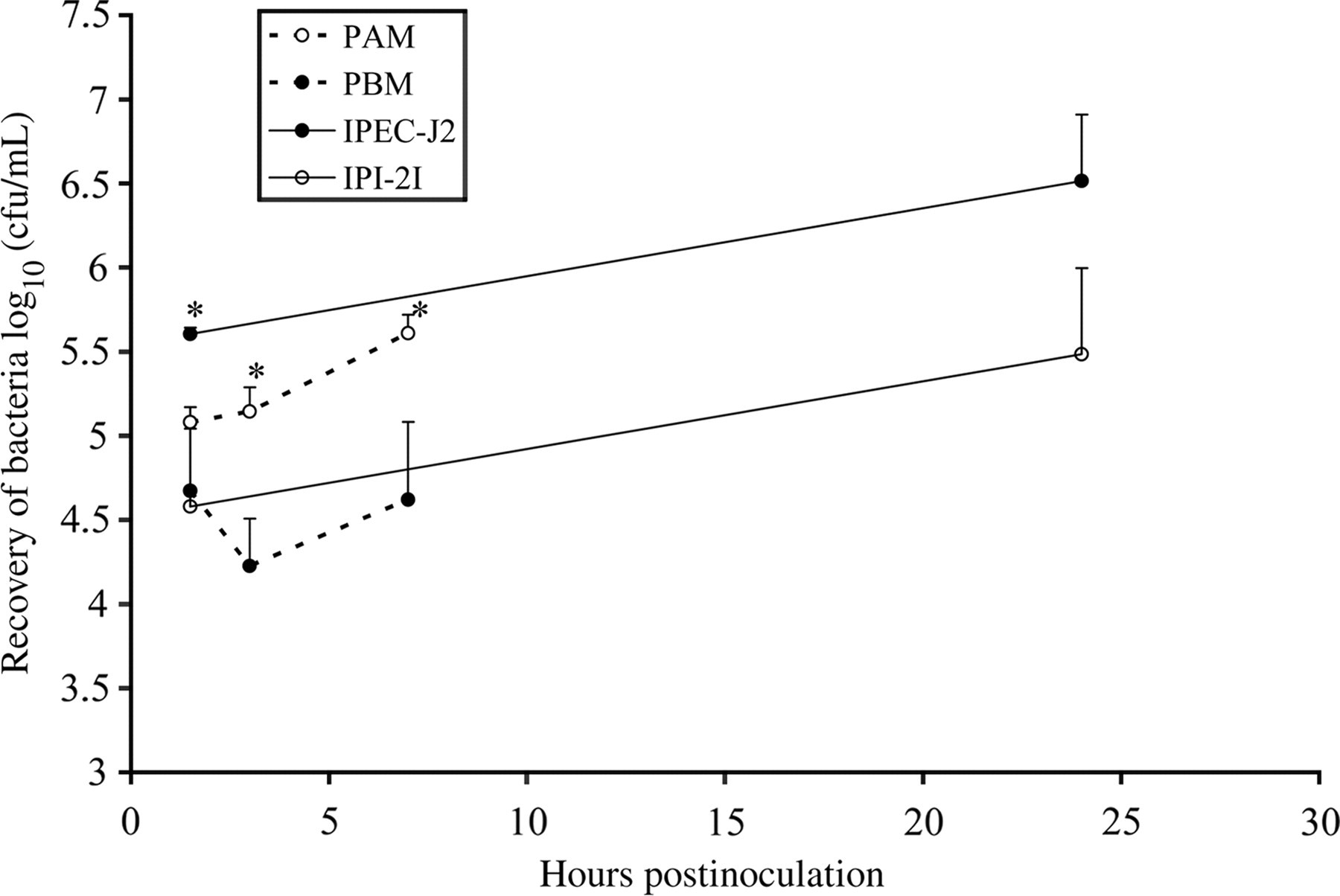
The results of the invasion and proliferation tests in both epithelial intestinal cell lines are summarized in Figure 1. Invasion was significantly (P < 0.05) higher in the polarized epithelial cell line IPEC-J2 compared with the IPI-2I cell line. Intracellular replication rates were not significantly different.
The invasiveness and intracellular replication of Salmonella Typhimurium in intestinal epithelial cells (full lines) and mononuclear cells (dotted lines). The results obtained in the polarized cell line IPEC-J2 and the monocytes are shown in black circles, the results obtained in the non-polarized cell line IPI-2I and the macrophages are shown in open circles. The log values of the number of intracellular bacteria are shown. The results represent the means of at least two independent experiments conducted in triplicate ± standard error of mean. An asterisk refers to a significant (P < 0.05) difference between both epithelial or mononuclear cells
Salmonella Typhimurium replicates in pulmonary alveolar macrophages, but not in peripheral blood monocytes
PAM and PBM were invaded to a similar extent. The bacteria replicated quickly in the PAM, but not in the PBM at 7 h after inoculation. At 24 h pi, the viability of PAM and PBM did not allow accurate titration of intracellular bacteria. The results of the invasion and proliferation tests are summarized in Figure 1.
Salmonella Typhimurim induces enteritis in the loop model and is killed rapidly by polymorphonuclear cells
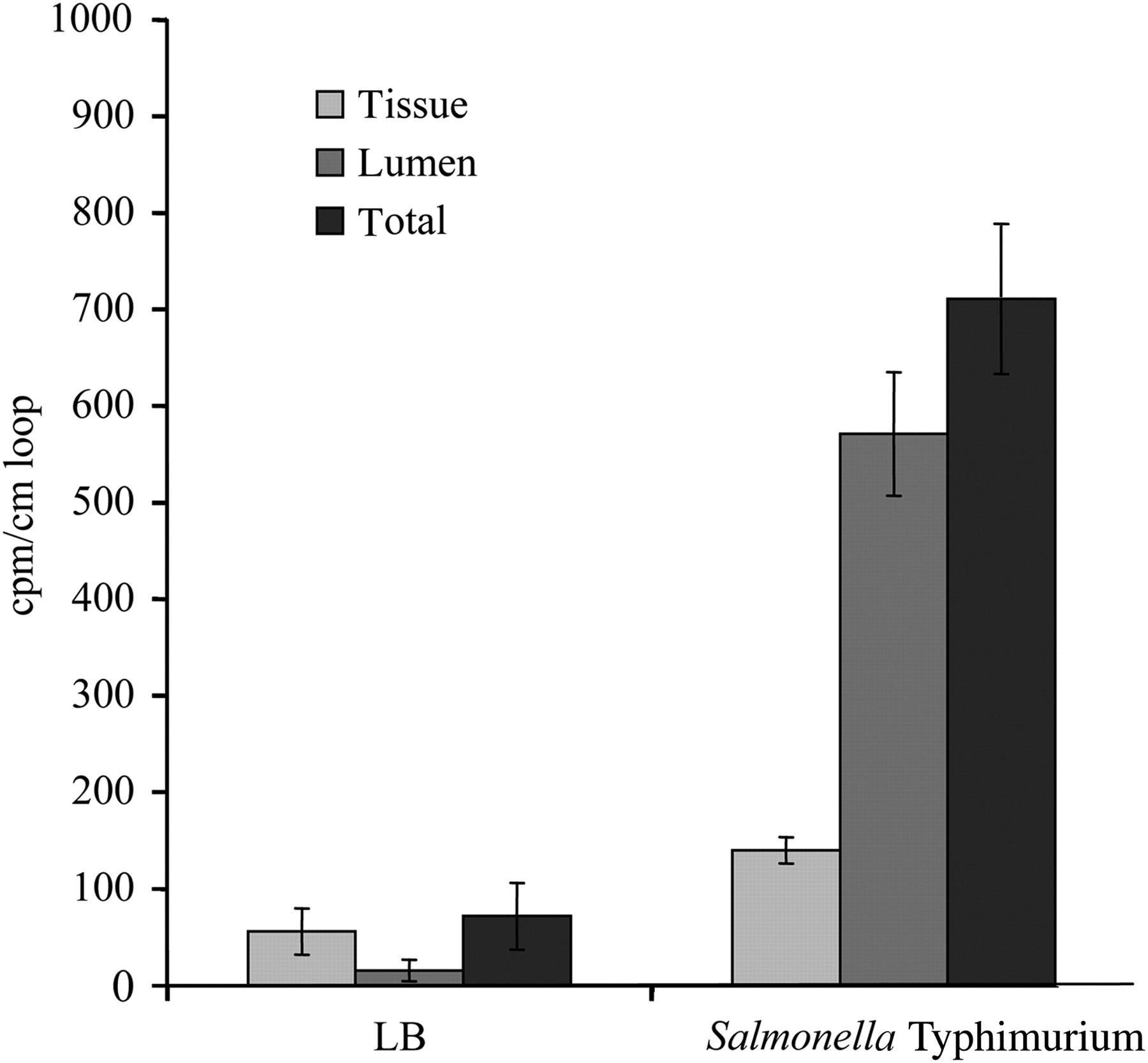
The results obtained in the intestinal loop model are shown in Figure 2. S. Typhimurium induced massive attraction of PMN in both the lamina propria and the lumen of the gut. The loops contained a rather small amount of slimy, pus-like substance. The number of PMN was significantly (P < 0.05) higher in the loops inoculated with Salmonella compared with the control loops. The intra- and inter-experimental variation between loops was relatively low.
Radioactive γ counts per minute (cpm) per cm loop as a measure for the early polymorphonuclear cell influx elicited by Salmonella Typhimurium in porcine intestinal loops. Approximately 1.5 × 109 colony forming units (cfu) of S. Typhimurium were injected into the loops and left for 12 h before analysis. Luria-Bertani broth (LB) served as negative control. The neutrophil influx in the gut tissue (tissue), in the contents of the gut lumen (lumen) and the total influx in both compartments (total) is presented. Each mean ± standard error of mean is calculated from three loops in two piglets
In the bacterial killing assay with PMN, 60 min after the addition of S. Typhimurium, 45% [±11% standard deviation (SD)] of the bacteria was killed by the PMN.
Course of a Salmonella Typhimurim infection in pigs
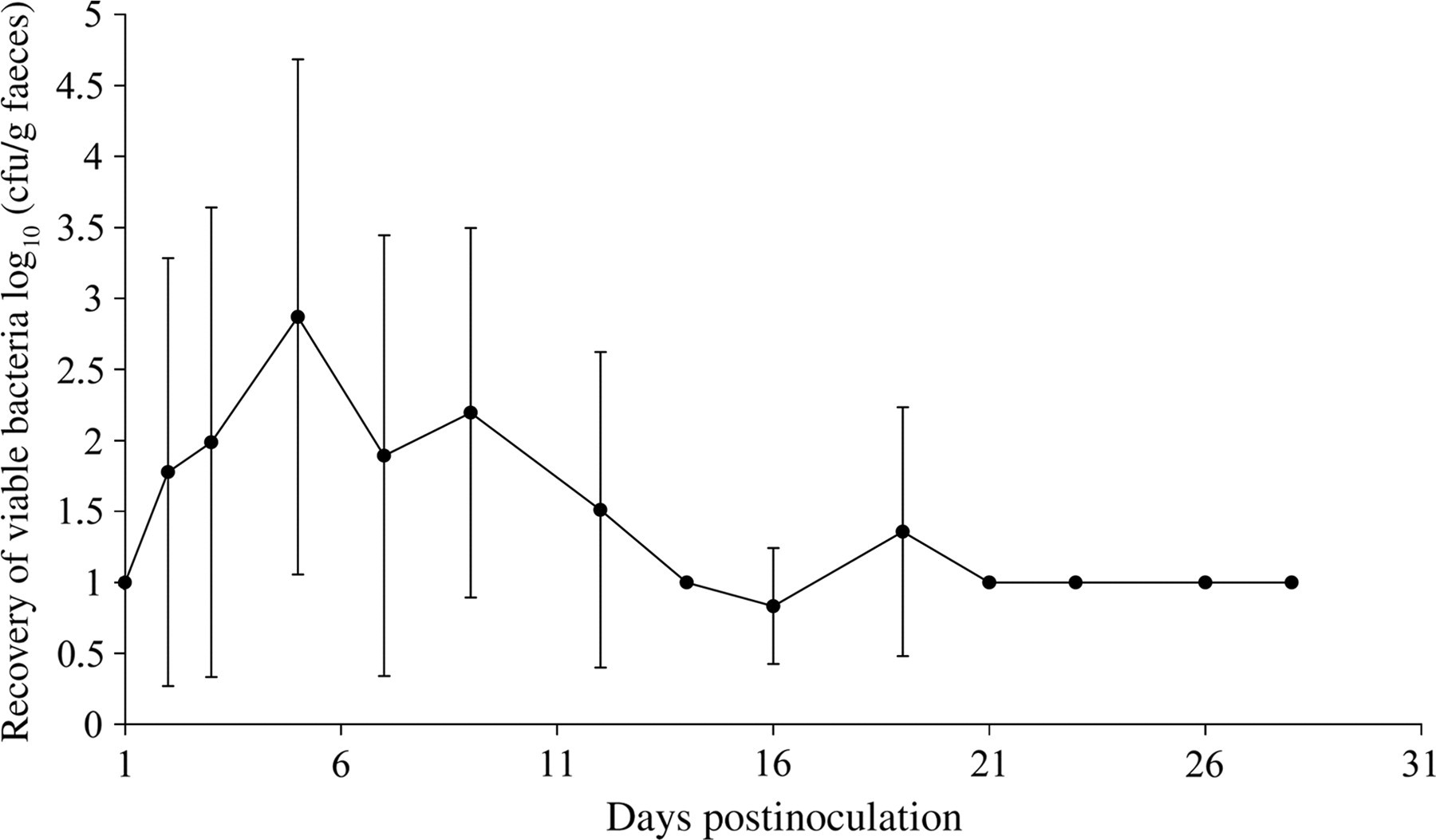
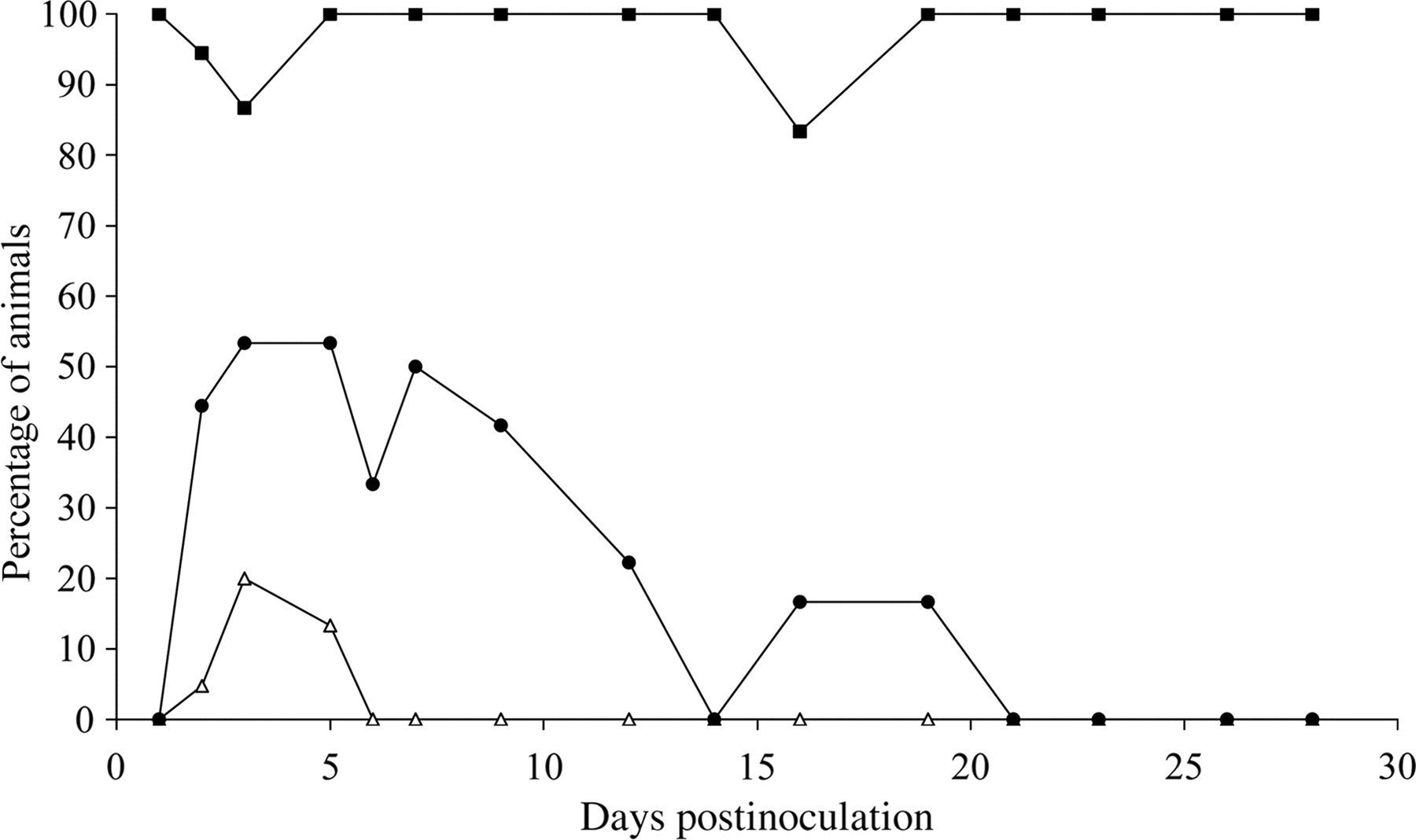
In the Salmonella-inoculated groups, all the pigs were colonized. Mean bacterial counts in the faeces were the highest at day 5 pi and the pigs were shedding S. Typhimurium intermittently until day 28 pi (Figure 3). The dynamics of the percentage of piglets that shed S. Typhimurium at least at enrichment level and of diarrhoea and fever (>40°C) are shown in Figure 4. Mean bacterial counts (log10 ± SD) in the various tissues of the infected pigs are shown in Table 1. Mean bacterial counts in the internal organs were highest at day 5 pi and remained stable at an enrichment level from day 21 pi on. Mean bacterial counts of the intestinal contents in 16 of the 21 cases were lower than the bacterial counts of the intestinal wall.
Faecal shedding of pigs inoculated with 107 colony forming units (cfu) of Salmonella Typhimurium, expressed as the average number of viable bacteria (log10) ± standard deviation of S. Typhimurium per gram faeces
Percentages of pigs that excrete Salmonella Typhimurium in their faeces (-▪-), show diarrhoea (-•-) or fever (-Δ-) after inoculation with 107 colony forming units (cfu) of S. Typhimurium
Colonization of Salmonella Typhimurium in different samples at days 1, 2, 5, 9, 14, 21 and 28 after experimental inoculation with 107 colony forming units (cfu) of S. Typhimurium, expressed as the average number of viable bacteria (log10) ± standard deviation (SD) of S. Typhimurium per gram organ. The frequency (F) shows the fraction of positive samples in relation to the total number of tissue samples
Days postinoculation
1
2
5
9
14
21
28
Samples
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
Mand. ln.
2/3
0.67 ± 0.58
2/3
0.67 ± 0.58
3/3
1 ± 0
3/3
1 ± 0
3/3
1 ± 0
3/3
1 ± 0
3/3
1 ± 0
Tonsil
2/3
1.83 ± 1.60
3/3
2.00 ± 1.73
3/3
2.60 ± 1.38
3/3
2.17 ± 2.02
3/3
2.17 ± 2.03
3/3
1.87 ± 1.51
3/3
1 ± 0
Lung
0/3
0 ± 0
2/3
0.67 ± 0.58
3/3
1 ± 0
2/3
0.67 ± 0.58
2/3
0.67 ± 0.58
2/3
0.67 ± 0.58
0/3
0 ± 0
Heart
0/3
0 ± 0
0/3
0 ± 0
1/3
0.33 ± 0.58
1/3
0.33 ± 0.58
1/3
0.33 ± 0.58
1/3
0.33 ± 0.58
2/3
0.67 ± 0.58
Liver
2/3
0.67 ± 0.58
1/3
0.33 ± 0.58
3/3
1 ± 0
3/3
1 ± 0
2/3
0.67 ± 0.58
1/3
0.33 ± 0.58
0/3
0 ± 0
Spleen
1/3
0.33 ± 0.58
1/3
0.67 ± 0.58
1/3
0.33 ± 0.58
1/3
0.33 ± 0.58
0/3
0 ± 0
0/3
0 ± 0
0/3
0 ± 0
Kidney
0/3
0 ± 0
1/3
0.33 ± 0.58
0/3
0 ± 0
2/3
0.67 ± 0.58
0/3
0 ± 0
0/3
0 ± 0
1/3
0.33 ± 0.58
Mesent. ln.
1/3
0.33 ± 0.58
3/3
1.90 ± 1.55
3/3
1.55 ± 0.94
3/3
1.56 ± 0.96
2/3
0.67 ± 0.58
2/3
0.67 ± 0.58
3/3
1 ± 0
Jejunum
1/3
1.64 ± 2.84
2/3
3.32 ± 2.88
3/3
1.75 ± 1.31
1/3
1.26 ± 2.18
1/3
0.33 ± 0.58
2/3
0.67 ± 0.58
3/3
1 ± 0
Ileum
1/3
0.84 ± 1.46
3/3
1 ± 0
3/3
1.94 ± 1.63
3/3
1.41 ± 0.71
2/3
1.21 ± 1.32
0/3
0 ± 0
3/3
1 ± 0
Caecum
2/3
1.62 ± 1.99
3/3
2.43 ± 1.25
3/3
3.87 ± 1.09
3/3
3.60 ± 0.56
3/3
1 ± 0
2/3
0.67 ± 0.58
3/3
1 ± 0
Contents jejunum
1/3
1.63 ± 2.83
2/3
2.23 ± 2.15
2/3
1.33 ± 1.15
2/3
1.33 ± 1.15
1/3
0.96 ± 1.66
0/3
0 ± 0
2/3
1.33 ± 1.15
Contents ileum
1/3
0.33 ± 0.58
2/3
0.67 ± 0.58
3/3
1 ± 0
2/3
0.67 ± 0.58
1/3
0.33 ± 0.58
3/3
1 ± 0
2/3
0.67 ± 0.58
Contents caecum
1/3
0.80 ± 1.38
2/3
1.50 ± 1.81
3/3
1.95 ± 1.66
3/3
1.59 ± 1.02
2/3
0.67 ± 0.58
3/3
1 ± 0
3/3
1 ± 0
In the negative control group, S. Typhimurium was not isolated from faeces or tissues and none of the piglets developed either diarrhoea or fever.
Salmonella Typhimurim infection is dose-dependent
In the Salmonella-inoculated groups, all the pigs were colonized. Mean bacterial counts (log10 ± SD) in the various tissues of these pigs are shown in Table 2. Results of the clinical scores (diarrhoea, anorexia, fever) and the mean body weight are summarized in Table 3. Significantly (P < 0.05) more pigs of group 3 developed diarrhoea than the pigs of groups 1 and 2. The loss of weight in group 3 was significantly (P < 0.05) higher than in groups 1 and 2 and the control group.
Colonization of Salmonella Typhimurium in different tissue samples at day 5 after experimental inoculation with either 105, 107 or 109 colony forming units (cfu) of S. Typhimurium, expressed as the average number of viable bacteria (log10) ± standard deviation (SD) of S. Typhimurium per gram organ. The frequency (F) shows the fraction of positive samples in relation to the total number of tissues
Infection dose
105cfu
107 cfu
109 cfu
Samples
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
F
Log10 cfu/g ± SD
Mand. ln.
2/3
0.92 ± 0.88
3/4
0.92 ± 0.7
4/4
1.41 ± 0.67
Tonsil
4/4
1.48 ± 0.96
4/4
1.44 ± 0.88
4/4
2.66 ± 1.16
Bronch. ln
1/4
0.25 ± 0.50
0/4
0 ± 0
1/4
0.25 ± 0.50
Lung
1/4
0.25 ± 0.50
3/4
0.75 ± 0.50
4/4
1 ± 0
Heart
0/4
0 ± 0
1/4
0.25 ± 0.50
3/4
0.75 ± 0.50
Liver
3/4
1.06 ± 0.78
4/4
1 ± 0
4/4
1 ± 0
Spleen
4/4
1 ± 0
3/4
0.75 ± 0.50
3/4
0.88 ± 0.64
Kidney
4/4
1 ± 0
3/4
0.75 ± 0.50
2/4
0.71 ± 0.89
Mesent. ln.
1/4
0.48 ± 0.96
3/4
1.21 ± 1.07
4/4
2.40 ± 0.62
Jejunum
2/4
1.06 ± 1.53
4/4
3.26 ± 1.57
3/3
3.51 ± 1.29
Ileum
3/4
1.08 ± 0.94
4/4
1.21 ± 0.29
3/3
3.27 ± 0.40
Caecum
4/4
2.76 ± 1.18
4/4
3.25 ± 0.96
3/3
4.46 ± 0.32
Contents jejunum
2/4
0.73 ± 0.92
3/4
2.35 ± 2.18
3/3
5.12 ± 1.67
Contents ileum
3/4
2.75 ± 0.50
4/4
2.33 ± 1.62
3/3
4.66 ± 0.58
Contents caecum
4/4
1 ± 0
4/4
1.49 ± 0.98
3/3
5.37 ± 0.51
Results of the clinical scores (diarrhoea, fever, anorexia) during five days postinoculation (pi) with either 105 (group 1), 107 (group 2) or 109 (group 3) colony forming units (cfu) of Salmonella Typhimurium and the mean gain in body weight at five days pi
Average fraction of animals showing
Groups
Diarrhoea
Fever
Anorexia
Mean weight gain (kg)
1
0.05
0
0
−0.14
2
0.1
0.05
0
−0.13
3
0.45
0.30
0.15
−0.49
Negative control
0
0
0
+0.15
Clinical scores are presented as average fractions of animals showing the respective clinical signs over the five-day period
The bacterial load in the ileum and caecum and in the contents of the ileum and caecum of pigs from groups 1 and 2 were significantly (P < 0.05) lower than the bacterial load of the pigs from group 3.
In the negative control group, S. Typhimurium was not isolated from the faeces or tissues at any time point.
Discussion
In this study, we evaluated several in vitro models to assess the virulence of S. Typhimurium strains.
An important virulence property of Salmonella is its ability to invade non-phagocytic cells. The invasion of intestinal epithelial cells is generally accepted as a major event in the pathogenesis of Salmonella infections.21 In this study, invasion was markedly increased in a polarized cell line compared with a non-polarized cell line. As polarized cell lines more closely approach the in vivo situation and since some Salmonella virulence features are lost using non-polarized cells,22 we suggest using polarized cell lines as much as possible.
One of the important characteristic features of Salmonella-induced diarrhoea is the massive influx of neutrophils.23 In no longer than 12 h, the porcine intestinal loop model was able to mimic this trait in a reproducible manner. Based on the fact that one piglet can provide approximately 30 individual loops, one can easily understand that this model has tremendous advantages compared with the models using individual pigs. Indeed, in several species including pigs, the intestinal loop model has been used to explore the intestinal inflammation phase during Salmonella infections.24–27 In contrast with the situation in calves,26 we did not find a large volume of watery fluid in the intestines, but rather a small amount of slime consisting mainly of neutrophils, so the measurement of luminal fluids may not be as useful in porcine loops as it is in bovine loops. However, some degree of fluid response can be induced, using very young piglets.28 Although a variant form of the intestinal loop model has been used to quantify invasion in the intestinal epithelial cells,29,30 we recommend using a polarized intestinal epithelial cell line, as it requires no animals to be used and it proves to be cheaper and less time-consuming. As interleukin-8 (IL-8) is a major cytokine in the development of Salmonella-induced intestinal inflammation,23 one could try to assess the ability of a Salmonella strain to induce an intestinal inflammation response by measuring the IL-8 production using either macrophages or epithelial cells.12,23 However, as other cytokines also play a role in the attraction of neutrophils to the lumen of the gut,23 and also because enteritis is the result of an extensive interplay of different receptors on different cell types,23 the intestinal loop model is a much more physiological and reliable model.
Intracellular survival and proliferation inside macrophages have been shown to contribute to persistent infections of Salmonella in mice.31,32 In spite of the fact that the systemic phase of a S. Typhimurium infection is hardly as important in pigs as in mice, the role of mononuclear cells, for example in lymphoid tissues, in the pathogenesis of S. Typhimurium infections in pigs must not be neglected. As in mice, the source of isolation of the mononuclear cells determined the degree of growth or survival of S. Typhimurium.31,33 In our case, the tissue macrophages allowed intracellular growth, whereas the monocytes were more restrictive. This finding may reflect the loss of important microbicidal functions of resident macrophages compared with blood monocytes.34 Therefore, it may be opportune to include both the cell types in in vitro assays.
Although the work with cell lines or single cell suspensions has its advantages, it also holds limitations. Simple steps of the pathogenesis, like the invasion in intestinal epithelial cells may easily be reproduced using a cell line, but the complex interactions of Salmonella with different cell types and the interplay of different host cell types among themselves resulting in, for example, the carrier state cannot be replaced by one single in vitro assay.35 This may mean that sometimes the use of an in vivo infection model is inevitable. The oral infection model may be considered to be the most physiological model. Oral infection of pigs resulted in the colonization of and persistence in both the tonsils and the gut, using low, moderate or high inoculation doses. However, it took an inoculation dose of at least 107 cfu to render all the animals positive for Salmonella in all the gut samples at day 5 pi. This uniformity may be important for standardization purposes in comparative assays. Inoculation with 109 cfu also guarantees uniform colonization, but this extremely high inoculation dose differs greatly from the natural infection, which is often established with low or moderate numbers of Salmonella.36 Although all the animals were colonized with 107 cfu, only a maximum 50% of the animals displayed signs of diarrhoea and only a maximum 20% had fever, both on days 3 and 5 pi. These results suggest that the oral inoculation model can be used to investigate colonization (at day 5 pi) and even persistence (at day 28 pi), but is not suited as an enteritis model or a systemic disease model, because of low and variable numbers of animals showing clinical signs.
In conclusion, the in vitro models and the intestinal loop model evaluated in the present study can be used to screen candidate knockout vaccine strains for attenuation. A promising selection of strains may then be used for further in vivo characterization. Data obtained from the porcine experimental infection model are helpful to determine optimal inoculation dose, time points of sampling and numbers of animals needed in future experimental infection studies.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the Federal Public Service Public Health, Food Chain Safety and Environment, DG 4, Brussels, Belgium and the Institute for the Promotion of Innovation by Science and Technology in Flanders (IWT Vlaanderen). The IPEC-J2 cell line was a kind gift of Dr Schierack, Institut für Mikrobiologie und Tierseuchen, Berlin, Germany. The technical assistance of G Massaer is gratefully appreciated. The authors wish to thank Dr L Herman, Dr M Heyndrickx and Lic N Botteldoorn for kindly providing the S. Typhimurium strain.
References
1.
BerendsBR, UrlingsHA, SnijdersJM, Identification and quantification of risk factors in animal management and transport regarding Salmonella spp. in pigs. Int J Food Microbiol1996;30:37–53
2.
KirkpatrickBD, McKenzieR, O'NeillJP, Evaluation of Salmonella enterica serovar Typhi (Ty2 aroC-ssaV-) M01ZH09, with a defined mutation in the Salmonella pathogenicity island 2, as a live, oral typhoid vaccine in human volunteers. Vaccine2006;4:116–23
3.
MethnerU, BarrowPA, GregorovaD, RychlikI. Intestinal colonisation-inhibition and virulence of Salmonella phoP, rpoS and ompC deletion mutants in chickens. Vet Microbiol2004;98:37–43
4.
MohlerVL, HeithoffDM, MahanMJ, Cross-protective immunity in calves conferred by a DNA adenine methylase deficient Salmonella enterica serovar Typhimurium vaccine. Vaccine2006;24:1339–45
5.
WoodRL, RoseR, CoeNE, Experimental establishment of persistent infection in swine with a zoonotic strain of Salmonella Newport. Am J Vet Res1991;52:813–19
6.
WoodRL, RoseR. Populations of Salmonella Typhimurium in internal organs of experimentally infected carrier swine. Am J Vet Res1992;53:653–8
7.
IsaacsonRE, KinselM. Adhesion of Salmonella Typhimurium to porcine intestinal epithelial surfaces: identification and characterization of two phenotypes. Infect Immun1992;60:3193–200
8.
Fedorka-CrayPJ, KelleyLC, StabelTJ, Alternate routes of invasion may affect pathogenesis of Salmonella Typhimurium in swine. Infect Immun1995;63:2658–64
9.
AlthouseC, PattersonS, Fedorka-CrayP, Type 1 fimbriae of Salmonella enterica serovar Typhimurium bind to enterocytes and contribute to colonization of swine in vivo.Infect Immun2003;71:6446–52
10.
SchwartzKJ. Salmonellosis. In: StrawE, D'AllaireS, MengelingWE, TaylorDJ, eds. Diseases of Swine. Ames: Iowa State Universtity Press, 1999:535–51
11.
DonnéE, PasmansF, BoyenF, Survival of Salmonella serovar Typhimurium inside porcine monocytes is associated with complement binding and suppression of the production of reactive oxygen species. Vet Microbiol2005;107:205–14
12.
BoyenF, PasmansF, DonneE, Role of SPI-1 in the interactions of Salmonella Typhimurium with porcine macrophages. Vet Microbiol2006;113:35–44
13.
SchierackP, NordhoffM, PollmannM, Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochem Cell Biol2006;125:293–305
14.
RussellWMS, BurchRL. The Principles of Humane Experimental Techniques.Special Edition. Potters Bar: Universities Federation for Animal Welfare, 1992
15.
BoyenF, PasmansF, Van ImmerseelF, Salmonella Typhimurium SPI-1 genes promote intestinal but not tonsillar colonization in pigs. Microbes Infect2006;8:2899–907
16.
KaefferB, BottreauE, VelgeP, Epithelioid and fibroblastic cell lines derived from the ileum of an adult histocompatible miniature boar (d/d haplotype) and immortalized by SV40 plasmid. Eur J Cell Biol1993;62:152–62
17.
RhoadsJM, ChenW, ChuP, l-glutamine and l-asparagine stimulate Na+ −H+ exchange in porcine jejunal enterocytes. Am J Physiol1994;266:G828–38
18.
DomP, HaesebrouckF, De-BaetselierP. Stimulation and suppression of the oxygenation activity of porcine pulmonary alveolar macrophages by Actinobacillus pleuropneumoniae and its metabolites. Am J Vet Res1992;53:1113–18
19.
DomP, HaesebrouckF, KampEM, Influence of Actinobacillus pleuropneumoniae serotype 2 and its cytolysins on porcine neutrophil chemiluminescence. Infect Immun1992;60:4328–34
20.
BarrioB, VangroenewegheF, DosogneH, Decreased neutrophil bactericidal activity during phagocytosis of a slime-producing Staphylococcus aureus strain. Vet Res2000;31:603–9
21.
SchlumbergerMC, HardtWD. Triggered phagocytosis by Salmonella: bacterial molecular mimicry of RhoGTPase activation/deactivation. Curr Topics Microbiol Immunol2005;291:29–42
22.
RaffatelluM, WilsonRP, ChessaD, SipA, SopA, SopB, SopD and SopE2 contribute to Salmonella enterica serotype Typhimurium invasion of epithelial cells. Infect Immun2005;73:146–54
23.
TükelC, RaffatelluM, ChessaD, Neutrophil influx during non-typhoidal salmonellosis: who is in the driver's seat?FEMS Immunol Med Microbiol2006;46:320–9
24.
WallisTS, HawkerRJ, CandyDC, Quantification of the leucocyte influx into rabbit ileal loops induced by strains of Salmonella Typhimurium of different virulence. J Med Microbiol1989;30:149–56
25.
MehtaA, SinghS, GangulyNK. Role of reactive oxygen species in Salmonella Typhimurium-induced enterocyte damage. Scand J Gastroenterol1998;33:406–14
26.
BoltonAJ, OsborneMP, WallisTS, Interaction of Salmonella Choleraesuis, Salmonella Dublin and Salmonella Typhimurium with procine and bovine terminal ileum in vivo.Microbiology1999;145:2431–41
27.
BoyenF, PasmansF, DonneE, The fibronectin binding protein ShdA is not a prerequisite for long-term faecal shedding of Salmonella Typhimurium in pigs. Vet Microbiol2006;115:284–90
28.
ClarkeRC, GylesCL. Virulence of wild and mutant strains of Salmonella Typhimurium in ligated intestinal segments of calves, pigs, and rabbits. Am J Vet Res1987;48:504–10
29.
UzzauS, LeoriGS, PetruzziV, Salmonella enterica serovar-host specificity does not correlate with the magnitude of intestinal invasion in sheep. Infect Immun2001;69:3092–9
30.
MeyerholzDK, StabelTJ. Comparison of early ileal invasion by Salmonella enterica serovars Choleraesuis and Typhimurium. Vet Pathol2003;40:371–5
31.
BuchmeierNA, HeffronF. Intracellular survival of wild-type Salmonella Typhimurium and macrophage-sensitive mutants in diverse populations of macrophages. Infect Immun1989;57:1–7
32.
LeungKY, FinlayBB. Intracellular replication is essential for the virulence of Salmonella Typhimurium. Proc Natl Acad Sci USA1991;88:11470–4
33.
HenselM, SheaJE, WatermanSR, Genes encoding putative effector proteins of the type III secretion system of Salmonella pathogenicity island 2 are required for bacterial virulence and proliferation in macrophages. Mol Microbiol1998;30:163–74
34.
Chitko-McKownCG, ChapesSK, BrownRE, Porcine alveolar and pulmonary intravascular macrophages: comparison of immune functions. J Leukoc Biol1991;50:364–72
35.
NiewoldTA, VeldhuizenEJ, van der MeulenJ, The early transcriptional response of pig small intestinal mucosa to invasion by Salmonella enterica serovar Typhimurium DT104. Mol Immunol2007;44:1316–22
36.
LoynachanAT, HarrisDL. Dose determination for acute Salmonella infection in pigs. Appl Environ Microbiol2005;71:2753–5