
Abstract
Following the fast pace of the growing field of stem cell research, retinal cell replacement is finally emerging as a feasible mean to be explored for clinical application. Although neuroprotective treatments are able to slow the progression of retinal degeneration caused by diseases such as age-related macular degeneration and glaucoma, they are insufficient to fully halt disease progression and unable to recover previously lost vision. Comprehensive, technological and intellectual advances over the past years, including the in vitro differentiation of retinal cells at manufacturing scale from embryonic stem (ES) cell and induced pluripotent stem (iPS) cell cultures, progress within the area of retinal disease modeling, and the first clinical trials have started to shape the way towards addressing this treatment gap and translating retinal cell replacement to the clinic. Here, summarize the most recent advances within retinal cell replacement from both a scientific and clinical perspective, and discuss the remaining challenges towards the delivery of the first retinal cell products.
Introduction
Every 20 milliseconds eyes - our windows to the world - provide us with information about our environment, allowing us to distinguish movements, objects, and thousands of color shades. Responsible for the perception and integration of sensory information is the retina, a layer of nervous tissue that in humans contains about 110 million rod and 6 million cone photoreceptors. 1 Located underneath the photoreceptor layer of the retina, there are two additional neuronal layers, one containing horizontal cells (HCs), bipolar cells (BCs) and amacrine cells (ACs) and another containing displaced ACs and retinal ganglion cells (RGCs) (Figure 1). In-between those layers, two plexiform layers, formed by the axonal and dendritic processes of the adjacent neurons, constitute a unique environment within which horizontal and vertical synaptic connections are established across the tissue. With little extracellular matrix, comparable to other parts of the central nervous system, retinal cytoarchitecture relies on Mueller glial cells (MGs). Amidst structural support, 2 due to the extension of elaborate cellular processes from the inner to the outer limiting membrane (ILM/OLM) and across both plexiform layers, MGs are essential for balancing neurotransmitters, trophic factors, and metabolites. 3

Overview of the healthy and neurodegenerative retina.
Given the immense structural complexity, retinal development is evolutionary conserved and highly regulated. After the induction of the eye field, two distinct domains are established, the retinal pigment epithelium (RPE) and the neural retina. Within the latter, the proliferation, differentiation, and interkinetic nuclear migration of retinal progenitor cells (RPCs)4,5 shape the distinct layering of the retina in an inside-out progression. 6 During this process, all subclasses of retinal neurons and MGs are born from one common RPC population, whereas astrocytes and microglia found in the adult retina do not originate from RPCs. During neurogenesis, RGCs are born first, then ACs and HCs, shortly followed by cone photoreceptors. Rod photoreceptors and BCs, alongside MG cells, are generated last and in some species, mature only after birth. 7 Noticeably, while neurogenesis lasts only a few hours in lower vertebrates, it can be spaced out over days in rodents or even weeks, in pigs, primates, and humans. 8 In higher vertebrates, the termination of retinogenesis coincides with the complete depletion of RPCs. 9 On the contrary, in lower vertebrates, RPCs become restricted to the ciliary margin zone. 10
From this point on, intrinsic regeneration upon damage/injury to retinal neurons, glia, or RPE only occurs in lower vertebrates through one of three mechanisms: (1) Cell cycle reentry of Mueller glial cells;11,12 (2) Reactivation of proliferation within RPC in the CMZ 10 or (3) De-differentiation of RPEs into RPCs.13–15 In higher vertebrates, the regenerative potential of MGs appears diminished, and even though recent findings demonstrate that MG-specific overexpression of ASCL1, together with histone deacetylase inhibition, can reinstate their regenerative potential in young mice, 16 the retina of higher vertebrates remains intrinsically vulnerable to disease and injury. Aside from slow, age-related loss of neurons over years, additional stress or insult results in neurodegenerative disorders, including conditions like glaucoma, and age-related macular degeneration (AMD) (Figure 1).
As projected by the National Eye Institute, the number of patients affected by neurodegenerative conditions is going to double by 2050, increasing the need for appropriate care. AMD alone, with a global prevalence of 8.7% (age group 45 to 85 years), is estimated to affect 196 million people by 2020. 17 Likewise, glaucoma is projected to impact 76 million people worldwide.18,19 While certain symptoms of both conditions can currently be managed for several years or even decades, the underlying neurodegeneration, causing the progressive loss of vision, cannot be counteracted.
In recent years, driven by advances in stem cell biology, cell/tissue replacement has become an obvious strategy to halt disease progression and recover previously lost vision. Ongoing collaborations between basic researchers and practicing ophthalmologists have led to the first clinical trials in the field of cell therapy (Table 1) and the promise for more trials in the near future. In this review, we will summarize the progress and achievements of cell replacement therapies in the retina, excluding cell-based therapies primarily oriented towards neuroprotection, and discuss remaining challenges.
Past and current clinical trials of cell replacement in the retina.
Summary of past and current clinical trials focused on assessing safety and efficacy of cell products for cell replacement within the retina. To date, trials are mainly focused on RPE replacement in AMD and Stargardt disease using ES-derived RPE. As a leader in the field, Astellas has performed multiple trials, the references to which have been limited to the three most recent examples. While subretinal suspension delivery is most common, more recent trials are assessing sheet delivery on scaffolds. For photoreceptor replacement, ReNeuron and Jcyte are the first to attempt human retinal progenitor transplantation within patients affected by retinitis pigmentosa. Trials including the transplantation of central nervous system or bone marrow derived stem cells as well as trials aimed at neuroprotection, rather than cell replacement, have been omitted. None of the cell products shown has been trialed further than phase II.

AMD, age-related macular degeneration; ES, embryonic stem cell; hESC, human embryonic stem cells; hRPC, human retinal progenitor cells; iPSC, induced pluripotent stem cell; RPE, retinal pigment epithelium.
The RPE, a story of translational success
In the healthy retina, the apical processes of the RPE closely envelope the outer segments of rod and cone photoreceptors, mediating diurnal outer segment recycling. Outer segments filled with stacks of membrane discs containing visual pigment have high metabolic demand and are renewed daily through shedding and reforming by 30%. Drusen deposits (AMD), accumulation of lipofuscin (Stargardt disease), or ischemic insult (choroideremia) can lead to disruption of normal RPE function, ultimately slowing photoreceptor metabolism and causing cellular damage. Although gene therapy was recently proven efficient in restoring RPE function in Leber congenital amaurosis (LCA),20,21 RPE replacement remains the only strategy for multifactorial diseases.
First reports of human RPE cultures date back to 1980, 22 and with the rise of embryonic stem (ES) cell culture, RPE was one of the first tissues to be differentiated in vitro, opening an unlimited array of possibilities for translational development. For the first trials in RPE replacement, there were attempts of transplanting freshly isolated adult and fetal RPE, immortalized adult RPE, and even translocation (rotation) of the RPE layer within the eye. While these attempts did not result in vision recovery, they aided the establishment of a strong technological platform for other modern therapies. At present, there are several ongoing phase I, I/IIa, and II clinical trials for pluripotent stem cell-derived RPE replacement (Table 1).
The reasons for the success of RPE replacement are multifaceted: (1) RPE cell biology and phenotypes are well described and conserved among species; (2) RPE differentiation from ES cells follows default pathways and results in easily identifiable, pigmented cells, streamlining readouts for protocol modifications; (3) Animal models of RPE dysfunction are readily available (Royal College of Surgeons rats, MerTK mice, etc.); (4) RPE cells mature in vitro with proper cell contacts and full functional phenotype (phagocytosis, polar growth factor production, visual pigment recycling); (5) The RPE quantity required for functional rescue is relatively small compared with photoreceptors; (6) RPE layer visualization within the retina is established using optical coherence tomography (OCT), adaptive optics scanning laser ophthalmoscopy (AO-SLO) and fundus imaging.
While for surgical delivery, RPE suspension injection into the subretinal space minimizes surgery time and damage to the adjacent tissues, animal studies have undoubtedly demonstrated an advantage of sheet transplantation over single-cell suspension. ES/induced pluripotent stem cells (iPSC)-derived RPE sheets can be supported by naturally produced Bruch’s membrane23–25 or polymer, 26 allowing to deliver RPE as a ‘tissue’, thereby avoiding epithelial to mesenchymal transition and preserving the extracellular matrix (ECM), cell contacts, cell polarity and hemidesmosomes, connecting RPE to the basal lamina. Although RPE, delivered as a suspension, survive and mature in the subretinal space, in vitro studies suggest that reacquisition of proper morphology and function is delayed by weeks, hence, further delaying the halt of ongoing neurodegeneration. Overall, the progress within the RPE replacement field, demonstrating transplant survival, integration, and the positive metabolic effect of transplants stands as the first demonstration of the advancement of the ES/iPS technology to the level of clinical relevance and applicability.27,28
Making neural retina and RPE: From 2D culture to 3D mini retinas
For cell replacement to become a clinically applicable therapy, the generation of purified, functionally competent retinal cells in therapeutically relevant quantities is essential. To achieve this goal, several distinct sources for retinal cells have been explored, including primary cells, in vitro differentiated cells from both ES and iPS cells as well as ex vivo and in vivo transdifferentiation from MGs29,30 or RPE. 31 Overall, high efficiency of differentiation, functional integration after delivery, safety profile, scalability, and cost-efficiency of the cell manufacture are prerequisites towards therapeutic application, as defined by the 2016 International Society for Stem Cell Research Guidelines for Stem Cell Research and Clinical Translation. While for RPE, as discussed above, most of these concerns have been addressed; the proper strategy for photoreceptor and RGC culture remains debated.
Animal studies of photoreceptor transplantation have shown that RPCs, postmitotic precursors, and mature photoreceptors all hold potential for cell replacement, challenging the use of a single default strategy as employed for the generation of RPE. While isolating photoreceptor precursors from developing tissue abolishes risks of transplanting nondifferentiated pluripotent cells, multiple donors are needed to retrieve the quantities required for a successful transplant, making this approach nonrelevant for clinical translation. Further in vitro expansion of RPCs32,33 to date does not provide the level of functional rescue, seen in primary cell transplants, 34 eliminating it as a potential solution.
With the dawn of stem cell research, the constraints posed by the reliance on primary tissue were lifted by the possibility for maintenance of retinal neurons in vitro, 35 closely followed by the de novo differentiation of retinal neurons from either ES or iPS cell lines, the strategy for which turned out to be functionally conserved across species, including humans.36,37 Initially relying on two-dimensional adherent neural rosette structures or simple monolayers [Figure 2(c) and Table 2],38–42 retinal differentiation protocols quickly diversified to suspension cultures, utilizing the formation of simple embryoid bodies (spheroids) or complex three-dimensional retinal organoids [Figure 2(a) and (b) and Table 2].37,43–47 Aside from those physical aspects, critical for scale-up and manufacture of clinical grade products, culture strategies vary with respect to their underlying molecular approach to differentiation.

Comparative overview of standard 2D and 3D retinal cell culture protocols.
Comparison of 2D and 3D cell culture systems.
Comparison of advantages and disadvantages of 3D versus 2D retinal cell culture, focusing on general aspects including cell purity and yield as well as high throughout adaptability. While 2D culture remains competitive with respect to cell yield and purity, novel 3D approaches are largely free of chemical additives driving differentiation, thereby recapitulating in vivo development more closely. In addition, due to a more complex tissue architecture, 3D cultures are paving the way towards more complex disease modeling in vitro.
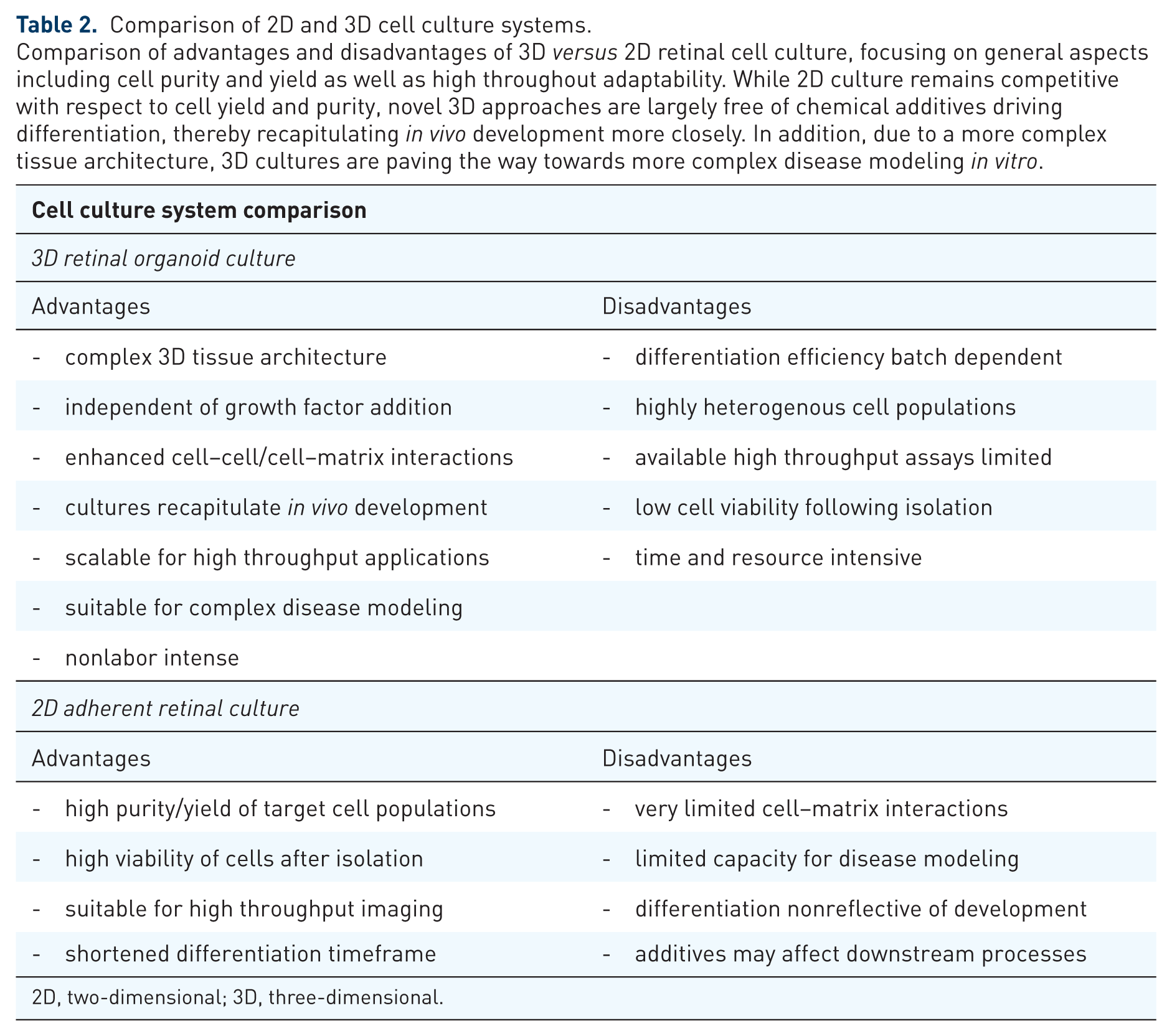
2D, two-dimensional; 3D, three-dimensional.
While Sasai and colleagues46,47 rely on default developmental pathways, and thus do not depend on artificial guidance for cell specification and fate commitment, other protocols direct cell fate determination by either the expression of transcription factors or modulation of growth factors 40 and cellular signaling pathways, including Wnt and Notch.44,48,49 Therefore, most directed approaches increase the relative abundance of specific cell types, but whether this level of artificial guidance might prove counterproductive for deriving bona fide retinal cells remains to be debated. Overall, despite collective efforts to optimize and standardize retinal differentiation within three-dimensional retinal organoids, the reproducibility of retinal induction remains variable, both with respect to cellular maturation, retinal structure, and layering. 50 External sheer stress, limitations in oxygen diffusion, and general batch dependencies with respect to cellular potency are the main culprits, rendering retinal organoids suitable for only a handful of high throughput screening applications. To address the sensitivity of retinal organoids to sheer stress, the addition of scaffolds is currently explored as an option to enhance structural stability and to improve handling in long-term cultures. Proposed scaffold matrices include: Matrigel, 51 RGD-alginate,52,53 laminin motif hydrogels, 54 methyl cellulose and gelatin.
Furthermore, since three-dimensional organoids recapitulate in vivo development, the duration of differentiation matches retinogenesis time in utero. Hence, in human-derived cultures, the time required for functional maturation of retinal organoids ranges in order of months, making development of a potential therapy time- and resource-consuming (Table 2). Nevertheless, the call to shorten culture times should be viewed with caution as modifications shortcutting differentiation time might alter cell identity.
Following differentiation, the next step for generating a clinically relevant cell product is the isolation of desired cell populations (rod/cone photoreceptors or RGCs). Contrary to RPE differentiation, yielding purity levels of 99.5%, cultures of neural retina, differentiated from pluripotent cells, contain RPC, multiple retinal and brain neuron and glia types, as well as single pluripotent cells. 55 Hence, targeted selection is essential for multiple reasons, with the depletion of proliferative cells being a top priority. Protocols developed for cell extraction are based on adhesive properties or expression of general surface markers, such as SSEA1 in pluripotent cells, 56 CD73 (ecto-5’-nucleotidase) in rod precursors and Thy1 in RGCs (Figure 3).39,57 To improve isolation specificity, the use of marker panels 58 instead of single markers has been suggested as well as the use of novel, more specific surface markers for the isolation of rod versus cone photoreceptors, including Kcnv2, Cacna2d4, and Cnga1.59,60 As for the actual technology used for pulldown, fluorescence activated cell sorting, immunopanning or magnetic activated cell sorting (MACS) are options, all of which can be performed under current good manufacturing practice (CGMP) conditions.37,61,62

Delivering a cell product: from stem cells to cell therapy.
In summary, the progress within the field of ES/iPS retinal culture to date has overcome the translational limitations of primary cells. Although still restricted to a certain extent, it allows for the generation of large quantities of retinal neurons, enabling translational studies.
Cell replacement therapy: a tale of single cells and sheets
Given the success of RPE sheet transplants, demonstrating the technical feasibility of the replacement surgery, and the complicated cytoarchitecture of the retina itself, retinal patches remain an intriguing idea for treating late stage neurodegenerative diseases. Unfortunately, following the first full thickness transplants, it became clear that amidst limited axonal outgrowth from mature neurons and the lack of proper tissue conservation protocols, full retinal patch replacement was unable to succeed.63,64 While animal studies using fetal and adult retinal sheets demonstrated significant survival, synaptogenesis between donor and recipient tissue was absent.65–67 Conceptually, causing further disruption within an already neurodegenerative retina to integrate a full thickness patch appears questionable. Nevertheless, this approach is still currently explored, 68 though focused towards the use of bioprinting and scaffolds69,70 including iPSC-derived cells to engineer ‘sandwiches’, comprised of choroid, RPE, and neural retina.
Technically less challenging, and surgically less disruptive, is the targeted replacement of individual retinal cell types (i.e. photoreceptors and RGCs), delivered as single layer sheets or cell suspension. While single layer sheets allow the control of cell density, alignment, and polarity, the formation of neural rosettes can impede outer segment alignment and thereby abolish transplant function. 24 On the other hand, single-cell delivery relies on migratory capacity of donor neurons to ‘fill in’ empty spots within the degenerating host retina.71,72 With respect to synaptogenesis, the presence of neighboring host cells can be assumed to enhance donor cell integration, addressing the pitfall of retinal patch transplants. It is notable that though devoid of regenerative potential, the vertebrate retina itself can remodel synaptic contacts as shown in ablation studies73–75 and during early neurodegeneration,76,77 as well as AMD, 78 arguing for the potential to integrate new neurons to existing circuits.
Also towards therapy development, the use of single cells is favorable as it allows for better isolation, characterization, cryopreservation, and quality control of the final product. However, cell delivery within biodegradable matrices, including hydrogels, poly(lactide-co-glycolide) (PLGA) scaffolds or ethylene-vinyl acetate (EVA)-copolymer holds the potential to temporarily protect donor cells, minimize rapid wash-off away from the site of injection, and might even provide structural guidance to enhance axonal outgrowth.70,79,80
Bringing back the light: photoreceptor replacement
High acuity vision is mediated by only 0.5% of the retinal area due to the distinct spatial distribution of rod and cone photoreceptors throughout the retina. While both photoreceptor types are aligned within tightly packed layers below the OLM, the 750μm center of the macula is exclusively formed of cone photoreceptors. 81 Hence, the foveal region, comprising only 5 out of 1000 mm2 of the retinal area, is the main region of interest for photoreceptor replacement. Distinct with respect to their outer segment morphology and photo pigment expression, rod and cone photoreceptors are also functionally different. While cones are essential for color vision and high visual acuity, rods are required for night vision and the perception of luminance. 82 To mediate the metabolic demands of outer segment turnover, as mentioned earlier, close interaction between RPE and outer segments is essential. Any disruption of RPE/photoreceptor proximity leads to progressive degeneration of photoreceptors, starting from the rods in the periphery, and eventually extending towards the cone-dense macula. Furthermore, various genetic defects affecting photoreceptor development can cause loss of photoreceptors and vision reduction in patients. Neuroprotective treatments and gene therapy can prevent such single-factor diseases or compensate for initial stages of degeneration, but remain inefficient in treating late stage degeneration. For cases of severe vision loss, retinal prosthesis devices were approved by the United States Food and Drug Administration (US FDA).83,84 However, those devices rely on external sensors to detect light, and only provide subthreshold resolution below the level required for functional vision.85,86
Targeted replacement of photoreceptors can overcome those challenges,57,87–89 and similar to RPE replacement, has been aided by the availability of a plethora of differentiation protocols involving the use of both ES and iPSC-derived cultures.46,90–93 Inspired by the early trials of RPE replacement, photoreceptor replacement was initially attempted using full thickness retinal sheets or patches.94–97 Later on, direct subretinal transplantation was attempted using undifferentiated RPCs,98–100 photoreceptor precursors56,57,72 and forebrain-derived neuronal progenitors (NPCs),34,101–103 which were expected to differentiate into photoreceptors due to the subretinal space microenvironment. While the integration of donor NPC progeny indeed influences gene expression within the subretinal space itself, 101 many research groups challenge the ability of NPCs to differentiate towards retinal lineages.104–106 Nevertheless, subretinal NPC transplantation is explored in the clinical setting for its neuroprotective mode of action.
For cell manufacture, the production of both NPCs and RPCs is scalable, with RPCs posing only minimal safety concerns since they are already pre-committed towards the retinal lineage. Though currently explored in clinical trials, the ability of RPCs to repopulate the retina and mature into functional photoreceptors remains questionable, partially because reliable functional readouts within the xenotransplant setting remain a challenge.23,107 When further addressing the concern of functional maturation, recent studies conclude that both primary and ES/iPSC-derived postmitotic photoreceptor precursors achieve structural and functional integration almost indistinguishable from host neurons.56,72,108–111. Initially focused on rods, more recent experiments also demonstrated feasibility of cone transplantation,87,88 often employing three-dimensional organoid cultures as a source of photoreceptors.
Despite the apparent progress in mouse and higher vertebrate models, the observation of cell fusion and cytoplasmic transfer between donor and host photoreceptors has called many results into question.112–116 While to achieve neuroprotection, the fusion of donor and host cells might proof functional to ‘rejuvenate’ damaged neurons, this outcome must be regarded as clearly distinct from actual cell replacement. First, since it requires the presence of remaining host photoreceptors, and secondly, because it does not address the actual challenge of functional integration, as it relies on previously established synaptic connections. Experimental options to distinctly address this issue are available, in the shape of cellular ploidy analysis, Y-chromosome tracing, and the assessment of human nuclear antigens when performing xenotransplants, but so far these have not been employed exhaustively. Interestingly, the observed cytoplasmic exchange has resulted in the proposal of targeted cell fusion as a treatment strategy involving, for example, MGs, 117 potentially changing the overall conceptual basis of photoreceptor replacement.
A bridge to the brain: retinal ganglion cell replacement
Within the retina, RGCs comprise the most diverse class of neurons, with about 30 subtypes described to date. 118 Aside from morphological features like their brain projection pattern, 119 dendritic field, and plexiform layer stratification, 120 RGC subclasses can be distinguished by their distinct transcription profile and physiological properties.121,122 Notably, distinct RGC subtypes are also different with respect to their intrinsic susceptibility to damage and regenerative potential.123,124 Compared with the other retinal neurons, RGCs are unique as their axons exit the eye through the optic nerve to connect all the way to the brain, where they are mapped in retinotopic fashion onto the visual cortex. Consequently, when considering the underlying challenges of cell therapy with respect to RGC replacement, it quickly becomes apparent that physical cell replacement in situ will not necessarily result in functional integration. While newly integrated photoreceptors only require establishing short range connections with the underlying horizontal and bipolar cells, RGCs must extend long axons and reestablish functional connections to their specific targets in the brain. Therefore, RGC replacement only became a theoretical option in the treatment of glaucoma and other optic neuropathies, following recent advances within the field of axon regeneration.125,126
Comparable with photoreceptor replacement, the first RGC transplantation attempts used either undifferentiated RPCs 127 or trans-differentiated glial cells,29,30 demonstrating some functional recovery in retinas of N-Methyl-D-aspartate (NMDA)- induced RGC damage/death. However, the observed functional rescue can be explained by neuroprotective effects. To date, the only successful transplants of RGCs, resulting in the integration of electrophysiologically functional neurons have been described for primary RGCs, with an overall success rate of only 10%.128,129 Additionally, only 1% of primary donor cells survived after transplantation, furthermore illustrating the low efficiency of RGC replacement achieved so far. For ES/iPSC-derived RGCs, no successful transplantations have been reported, probably due to the remaining challenges in the generation and isolation of large quantities of differentiated neurons from retinal organoid cultures.
As alluded to earlier, the initial integration of donor cells is only the first hurdle to functionally relevant RGC replacement. Therefore, advances within the field of axon regeneration must be considered as closely tied to RGC therapy success or failure. In recent years, factors like osteopontin, PTEN inhibitors and a variety of trophic factors, like pigment epithelium-derived factor (PEDF) 130 and ciliary neurotrophic factor (CNTF), 131 have been described to aid RGC survival, axon elongation, and regeneration after optic nerve injury/crush. Aside from those mainly cell autonomously regulated factors, a recent study proposed an effect of mobile zinc dysregulation within neighboring interneurons as primary factor in the death of RGCs after injury, demonstrating that zinc chelators had a beneficial effect towards neuroprotection and axon regeneration. 132 While those studies are encouraging with respect to the physical aspect of bridging the distance between the eye and the brain, they do not address the question whether in vitro-derived RGC axons will be able to form target-specific synapses once they reach beyond the eye. Interestingly, in vitro human induced pluripotent stem cells (hiPSC)-derived RGCs displayed some target-specific responses when cocultured with different brain regions, 133 arguing intrinsic target selectivity, though the overall evidence remains limited. Furthermore, all studies addressing the rewiring of the retinal network are heavily focused towards the reestablishment of connectivity between the newly integrated RGCs and the brain, leaving aside the fact that also within the retina, the donor RGCs are required to establish selective synaptic connections to enable functional integration. While the spatial proximity of synaptic targets clearly suggests for this step to be more likely to occur spontaneous than the rewiring of axonal connections to a far out target like the brain, the assumption of simplicity might be ill-guided.
To address this gap, it might be advised to refocus research attention towards the development of the retina and the initial establishment of synaptic circuits within. Following the logic of retinal circuit assembly occurring over temporally distinct phases in development, the microenvironment instructing RGC and amacrine cell or photoreceptor and bipolar cell wiring can be assumed to be vastly different. Since in contrast with lower vertebrates in which neurogenesis might only last a few hours, human neurogenesis extends over several weeks, it appears imminent to assume for synaptogenesis to be cell type specific and to assess factors involved in each of its phases. Furthermore, the development of a methodology for single-cell functional studies in vivo will be required to improve functional assessment of transplantation outcome. This is crucial due to the immense diversity among RGC subtypes, rendering it difficult to assess which subtypes are essential to which aspects of vision, especially when trying to compare across different species where subtypes might be distinct from one another. Taken together with the recognition of their subtype-specific susceptibility to damage, RGC replacement poses the biggest challenges yet in the field of retinal cell replacement, far surpassing the obstacles previously addressed when replacing homogenous, single-cell type tissues like the RPE.
To replace and rewire: the promise of combinatorial therapy
Arising from the success in photoreceptor/RGC neuroprotection studies and discussed advances in axon regeneration, the use of various combinatorial treatments has become a new focus within cell replacement studies. The nature of tested cotreatments is manifold, as are their aims, including the improvement of donor cell survival, integration and axon outgrowth by modification of the host microenvironment or the enhancement of donor cell resilience to handling pre-transplantation.
To improve donor cell integration, enzymes and chemical modulators of glial response have been tested successfully. Matrix metalloprotease 2 (MMP2), which causes the selective digestion of the ILM/OLM, helps to eliminate the physical border to cell entry to the RGC or outer nuclear layer.134–136 Similarly, low doses of aminoadipic acid (AAA) enhance donor cell integration by causing glial end foot retraction from the ILM/OLM, thereby weakening its barrier integrity.137,138 Aside from its glial toxic effect, AAA also reduces gliosis, leading to a reduction in donor cell clearance from the tissue upon transplantation, consequently improving overall survival. 139
Cytokines and trophic factor supplementation, including factors like brain-derived neurotrophic factor (BDNF) 140 , glial cell-derived neurotrophic factor (GDNF) 141 , platelet-derived growth factor (PDGF) 142 , CNTF131,143 and interleukin (IL)-6 144 have shown to be broadly successful in prolonging/enhancing neuron survival in various animal models of photoreceptor and RGC death, including retinal detachment, optic nerve crush, and laser-induced ocular hypertension. 145
Currently, routes of small or large compound delivery to the retina pre-, mid- or post-transplant are being explored, including: adeno-associated virus (AAV)-mediated delivery, 143 light-activated or thermosensitive liposome transfection146,147 and other sustained release systems.148,149 Trans-plantation of cells overexpressing growth factors is another option; CNTF150,151, GDNF 152 , insulin-like growth factor (IGF)-1 153 and nerve growth factor (NGF) 154 have been delivered by ARPE19, neural stem cells (NSC) and mesenchymal stem cells (MSC) cells respectively. Furthermore, a vast spectrum of neuroprotective small molecules is being tested, including anti-apoptotic drugs, histone deacetylase inhibitors, RIP kinase inhibitors and antioxidants.155–157 Some of them are also being used as cotreatments for cell delivery, such as N-acetyl-L-cysteine. 158 Taken together, cotreatments allow to optimize distinct facets of the process from cell delivery, to cell integration, and network rewiring, and should be expected to improve the odds towards the generation of an effective neuronal cell replacement therapy for the retina within the next decade.
Current challenges
Understanding the role of the (disease) microenvironment
Although retinal structure and metabolism is highly conserved among mammals, and the eye has an active immune privilege mechanism, the functional integration of xenotransplants into rodent retinas is not comparable to syngeneic and allogeneic transplants in patients. Furthermore, retinal immune privilege should not be overstated when considering transplantation into a neurodegenerative environment, within which ongoing gliosis, 159 inflammation and immune infiltration can change the microenvironment significantly from what is being observed within healthy hosts. 160
To overcome the limitations of xenotransplants, two approaches have been developed: the use of analogous cell models, and xenotransplantation into humanized models. While the first approach requires adaptation of cell culture and differentiation protocols for primate or pig cells, it provides the advantage of a similar surgical setting and allows to test additional parameters, including delivery strategies and tools, volume, concentration, and total dosage of cells, as well as the requirement for immune suppression. Humanized mouse models have been very effective in understanding the immunology of transplantation, 161 and Rag2 knockouts have been effective hosts for human embryonic stem cells (hESC)-derived photoreceptor precursors. Nevertheless, while the latter allows testing of the exact cell product to be used in the clinic, functional integration studies are limited. Additionally, addressing the immune component of the therapy isolated from the disease environment might be shortsighted.
Studies tracking survival of transplanted retinal sheets showed them being targeted by the immune system and ultimately becoming fragmented and apoptotic. Other studies using RPCs as donor cells for photoreceptor replacement do not report any immune response nor improvement of transplant integrity after transient immune suppressive treatment in allotransplants. 162 Noticeably, the hosts used in this study, experienced no prior insult to the retinal microenvironment, proposing intact immune privilege prior to transplant. In contrast, within hosts subject to ongoing neurodegeneration, infiltration of immune cells can be assumed as apparent even prior to transplantation, increasing the likelihood of donor cells being targeted. One way to circumvent immune targeting and further improve allotransplantation altogether is the use of major histocompatibility complex (MHC)-matched iPSCs for the generation of donor cells, as demonstrated within the field of RPE replacement. 163 Likewise, to the changes observed within diseased retinas and associated differences in transplantation success depending on the used disease model, 108 the retinal microenvironment also changes upon aging, affecting transplant outcome. In mice, transplantation into younger animals was more successful than in adults, underlining this assumption. 164
Another independent factor to consider are extracellular matrix components, not only as a hindrance towards cellular integration in the shape of the ILM/OLM, but also as essential, instructive cues required for cell polarity and spacing, as well as dendritic arborization and axonal guidance. 165 Hence, while disruption of those membranes appears conducive to enhancing donor cell integration, it might have to be considered that with ongoing neurodegeneration, causing host neurons to die, remaining extracellular matrix might contain the only remaining cues for newly integrating cells. Consequently, when assessing animal models of retinal damage, the ability of donor cell integration should not only be correlated to remaining cell numbers within the host.
Developing and delivering clinical grade products from iPSC/ES
Although cell therapy is a century behind standard pharmacological drug development, RPE and limbal stem cell transplantation studies have demonstrated the requirements for the design of cell products. While standard drug development criteria, like safety and efficacy, are applicable, the living nature of cell products slightly shifts priorities.
Safety is a top criterion for cell therapy design and development, partially due to the novelty of the approach and limited availability of historical data. Major concerns for cell manufacture are microbiological contamination, the presence of nondifferentiated cells, acquired mutations (primarily tumorigenic) and potential genetic transfer. Quality assurance criteria and tools are very sensitive and specific for detecting viruses, bacteria, and fungi. However, the threshold for pluripotent and partially differentiated cells present in cell drug products is not yet established. Importantly, these criteria require tight control of manufacture conditions since they cannot be dealt with later. Similarly, treatment efficacy criteria need to be addressed early on, but so far have often not been the primary focus during the development process. 166 Consequently, though it is possible to probe donor cell function individually, the actual contribution of the transplant to vision is difficult to assess, and visual function rescue in rodent disease models often can be attributed to trophic factor release. To disentangle the contribution of neuroprotective effects and actual cell replacement in animal models, donor cells are traced by the introduction of fluorescent probes or iron micro particles, allowing further insights into the mechanism of action.
Within clinical trials, the tracing of donor cells has yet to be resolved since there are no US FDA-approved technologies to do so to date. Hence, the mode of action and clinical decisions about post-transplantation therapy adjustments, including the need for repeated treatment in the same or adjacent areas, immune suppression or other co-treatments, are inherently difficult to derive. While transplanted RPE can be seen on OCT due to the hyperreflectivity of the pigment,166,167 other retinal neurons are transparent and therefore indistinguishable. Consequently, though donor cell placement at the initial side of transplantation can be established in case of subretinal transplants due to an immediate local retinal thickness increase in OCT, it is impossible to determine what happens post-transplantation. While ideally, donor cells integrate into the transplanted area over time, they might be phagocytosed or have migrated away from the transplant area.
Overall, the clarity and accessibility of the retina for structural and functional imaging is remarkable, with high resolution fundus imaging, OCT and spectral domain optic coherence tomography (SD OCT), heidelberg retinal tomography (HRT) and adaptive optics scanning laser ophthalmoscopy (AO-SLO) combined providing unprecedented resolution up to single-cell resolution in all dimensions. Therefore, complications including inflammation, ischemia, or neovascularization can be monitored efficiently, aiding the assessment of therapy safety. With respect to the delivery/surgical procedure, potential complications overall are expected to be comparable with other ocular surgeries. The Limited space of the eye and high metabolic demand of neural tissue renders spatial interactions between neurons, glia, RPE and vasculature very important, thus disruptions by retinal detachment or tear, partial vein occlusion, or glial scar formation can lead to irreversible alterations and damage. One of the initial concerns for subretinal delivery, the persistent detachment of the retina from the RPE in the transplant area, has been resolved, as for hundreds of patients treated by now with cells or gene vectors, the initial detachment was observed to spontaneously resolve within 48 hours.
Further potential complications in the procedure are associated with the cell material itself or its carrier medium (delivery solution or hydrogel), which may be prone to cause immune responses, especially once certain structural barriers, like vessels, are compromised either due to age-related/disease-associated degeneration or by the procedure. In addition, parts of the delivered cell product can result in the formation of clusters and clots; to address the latter, cells are formulated right before the injection in buffers without bivalent ions. Overall, the procedure itself, aside from the expertise needed to perform vitreoretinal surgery, also presents the additional challenge of on-time preparation of the cell product. A potential solution is the use of ‘thaw-and-inject’ formulations that do not require synchronized cell manufacture at a good manufacturing practice facility, delivery scheduling and formulation of the cell product for injection on site.
Since the ongoing clinical studies are focused on safety (Table 1), it is premature to draw conclusions on the efficiency of cell therapy. Nevertheless, it should be noted that readouts of visual and retinal function are diverse, but not universal: they usually do not cover the whole range of sensitivity. Thus, the proper selection of several methods for clinical trial design is crucial for future success. Objective methods to quantify low vision are only starting to develop with the dawn of gene and cell therapy, and currently are primarily focused on rod rescue (RPE transplants) and replacement (RPC transplants). While within those trials, readouts for contrast sensitivity, maze tests, perimetry, and microperimetry are informative measures to detect changes, ultimately longitudinal imaging of both structure and function with proper mapping of the region of interest should be employed to track the tissue structure and assess repopulation and functional restoration or delay in disease progression.
All the factors mentioned above will influence the length of future clinical trials. While for RPE/photoreceptor replacement, 6 months might be sufficient to detect changes in vision, it will undoubtedly take years to complete similar studies for the replacement of RGCs. 168
Concluding remarks
Leading to the clinic, in recent years many fundamental obstacles in cell manufacturing and retinal disease modeling have been addressed. At present, the methodological development and critical evaluation of functional outcome measures, as well as the integration of combinational therapies, will be essential to drive the current momentum of the field towards clinical translation. To tackle the most difficult of the underlying challenges, including synaptogenesis and retinal rewiring, reflecting on developmental concepts might be advised.
While clinical translation of retinal cell replacement is currently shaped by the experience gained from RPE, the overall critical advances, moving the replacement of retinal neurons from a laboratory model to a clinically relevant treatment, are still lacking. Largely fueled by excitement arisen from the outcome of cell transplantation into animal models of retinal disease, the next steps towards clinical translation must be approached with care. Considering the discovery of artifacts like cytoplasmic transfer and the ongoing quest to distinguish neuroprotective effects from functional improvements attributed to actual cell replacement, planning and execution of clinical trials needs to be based on a maximum inclusion of objective metrics for safety, efficacy, and mechanism of action.