
Abstract
Symptoms of posttraumatic stress disorder include hyperarousal, avoidance of trauma-related stimuli, re-experiencing of trauma, and mood changes. This review focuses on the frontal cortical areas that form crucial links in circuitry pertinent to posttraumatic stress disorder symptomatology: (1) the conditioned fear extinction circuit, (2) the salience circuit, and (3) the mood circuit. These frontal areas include the ventromedial prefrontal cortex (conditioned fear extinction), the dorsal anterior cingulate and insular cortices (salience), and the lateral orbitofrontal and subgenual cingulate cortices (mood). Frontal lobe structural abnormalities in posttraumatic stress disorder, including volumetric reductions in the cingulate cortices, impact all three circuits. Functional analyses of frontal cortices in posttraumatic stress disorder show abnormal activation in all three according to task demand and emotional valence. Network analyses reveal altered amygdalo-frontal connectivity and failure to suppress the default mode network during cognitive engagement. Spine shape alterations also have been detected in the medial orbitofrontal cortex in posttraumatic stress disorder postmortem brains, suggesting reduced synaptic plasticity. Importantly, frontal lobe abnormalities in posttraumatic stress disorder extend beyond emotion-related circuits to include the lateral prefrontal cortices that mediate executive functions. In conclusion, widespread frontal lobe dysfunction in posttraumatic stress disorder provides a neurobiologic basis for the core symptomatology of the disorder, as well as for executive function impairment.
Keywords
Posttraumatic stress disorder (PTSD) afflicts approximately 3.5% of the general population of the United States. 1 By definition, PTSD is a disorder that occurs in individuals who have been exposed to a traumatic experience. A wide range of events, e.g., natural disasters, terrorist attacks, and sexual abuse, can trigger the onset of PTSD, and not surprisingly, the disorder is particularly prevalent (∼20%) in combat-exposed veterans.2–5 Approximately one third of people diagnosed with PTSD have a severe form of the disorder 1 and experience debilitating symptoms that disrupt family dynamics, other social interactions, and workplace functioning.6,7 Frequently, PTSD occurs in conjunction with depressive and anxiety symptomatology and/or substance abuse. 1 In addition, increased suicidal ideation and behaviors occur in association with PTSD, 8 and the risk for suicide is significantly higher in military veterans with PTSD relative to the general population. 9
Neurobiologic studies of PTSD have largely focused on the amygdala and hippocampus as reviewed in detail elsewhere.10,11 Among other functions, the amygdala mediates emotion-related processing, including fear conditioning and extinction, while the hippocampus plays an important role in providing contextual memory for emotion-related processes.12–15 Major PTSD-related alterations have been found in these regions, including hyperactivity of the amygdala in response to trauma- and non-trauma-related, emotionally charged stimuli,10,11,16–18 although the findings are not entirely consistent. Smaller hippocampal volume in association with PTSD is perhaps the most replicated pathologic correlate of the disease,19–21 whereas both hippocampal activation and deactivation have been reported in PTSD during exposure to emotion-laden stimuli or during performance of memory tasks.10,22
To examine the role of frontal lobe in PTSD, an internet search was conducted using the terms “PTSD” and “MRI” and encompassing the years 2000–2018. Note that a few seminal articles published earlier are included as these were referenced multiple times in the more recent literature. Studies were included if they reported a structural or functional comparison between individuals diagnosed with PTSD and a comparison group of either trauma-exposed or trauma-naïve participants. Studies of PTSD in which participants were children or adolescents were excluded. In addition, only functional connectivity studies that shed further light on the structural and functional magnetic resonance imaging (MRI) findings in the frontal lobe have been included.
This review spotlights the role of the frontal cortex in PTSD in order to further explore the neurobiologic substrates of PTSD dysfunction. We focus on three frontal lobe circuits that have been identified as important to understand the PTSD symptomatology17,23,24: (1) the conditioned fear extinction circuit, (2) the salience circuit, and (3) the mood circuit—and review the literature on morphologic and functional alterations in the frontal lobe nodes of these circuits. Our goal is to forge a link between the burgeoning literature on neuroimaging findings in PTSD and known functional circuits in the frontal lobe to better understand the neurobiologic underpinnings of the disease.
Fear Conditioned Extinction Circuitry
It has been hypothesized that the circuitry of fear conditioning and fear conditioned extinction are intimately involved in the pathology of PTSD.25,26 Fear conditioning, which is the pairing of a negative, unconditioned stimulus (UC) to a neutral, conditioned stimulus (CS), is critically mediated by the basolateral amygdala. 12 In PTSD, the unconditioned negative stimulus is the traumatic event, and the conditioned stimuli are the sights, sounds, and other sensory experiences that occur concurrently with the event. In normal subjects, physiological responses, such as visceral, cardiac, visual, and auditory sensations, that are associated with fear re-experiencing are gradually “extinguished” by an active re-learning process. In PTSD, a failure to extinguish these fear responses contributes to the persistent physical and cognitive symptoms of re-experiencing the trauma, including increased autonomic arousal and phobic behaviors. 25 Importantly, extinction of conditioned fear is achieved by the formation of new memories, not by erasure of the initial fear conditioned association.27,28 Furthermore, although initial extinction is a function of the basolateral amygdala, 29 memory or recall of extinction is modulated by a circuit involving the ventromedial prefrontal cortex (vmPFC), a cortical region equivalent to the infralimbic cortex in rodents.14,17,28,30 The human vmPFC encompasses the subgenual anterior cingulate cortex (sgACC, BA25), the rostral anterior cingulate cortex (rACC, BA32), the medial prefrontal cortex (mPFC, BA10), and the medial orbitofrontal cortex (mOFC, BA11). In particular, the sgACC has been implicated in human studies of retention of fear extinction. 30
The amygdalar circuitry involved in fear conditioning is located primarily in the basolateral amygdala (Figure 1).12,31,32 Prominently involved regions include the lateral (LA), basal (B), accessory basal (AB), and central (CE) nuclei as well as the intercalated cell masses (ICM).
32
Nocioceptive information reaches LA, AB, and CE through the spinal cord, brain stem, thalamus, and cortex. Neutral sensory information, e.g., from the auditory thalamus and cortex, is conveyed mainly to LA. Contextual information from the hippocampus is transmitted to LA, as well as to B and AB. Thus, the convergence of UC (nocioceptive input) with CS (other sensory information) and integration of UC with contextual information involves LA, B, and AB. During fear conditioning, LA disinhibits the output of CE neurons, although LA projects only indirectly to CE via ICM or B.
33
As the major output nucleus of the basolateral amygdala, CE projections mediate the entire panoply of fear behaviors. CE stimulates hypothalamic and brain stem regions that govern blood pressure control, respiration, freezing behavior, and the release of adrenal stress hormones and projects to monoaminergic nuclei that may alter perception and cognition via thalamic and cortical modulation.12,31
Conditioned fear extinction circuitry. Fear conditioning is mediated in the basolateral amygdala where nocioceptive information is integrated with auditory and visual sensory information, as well as contextual information from the hippocampus. Fear conditioned responses are enacted via efferents to the hypothalamus and brain stem. Extinction of fear conditioning involves new learning in the vmPFC (shaded area), including cingulate ((Brodmann area (BA)25, BA32)) and orbitofrontal cortices (BA10 and BA11), and projections of the vmPFC to the amygdala to curtail the fear conditioned output of the amygdala. Frontal area designations according to Ongur and Price.
38
AB: accessory basal nucleus; B: basal nucleus; CE: central nucleus; ICM: intercalated cell masses; LA: lateral nucleus.
Although the acquisition of the initial stages of conditioned fear extinction occurs within the basolateral amygdala with minimal requirement for additional input, the long-term retention of fear conditioned extinction requires participation of vmPFC-amygdala connections in humans (the infralimbic cortex in rodents).28,34 The vmPFC receives glutaminergic input from the mediodorsal nucleus of the thalamus, the hippocampus, the basolateral amygdala, and the auditory association cortex. It is hypothesized that exposure to the CS without the US activates glutaminergic inputs to the vmPFC, resulting in long-term potentiation at one or more of these synapses. 28 In turn, the vmPFC projects to the amygdala resulting in depressed CE output, perhaps through the activation of ICM inhibitory neurons. 28
Salience Circuitry
Salience circuitry (Figure 2) tags sensory information with emotional valence, directs attention to salient stimuli, and governs motor and visceral responses to such stimuli.35–37 Functional imaging analyses in human subjects have identified the dorsal anterior cingulate cortex (dACC, BA24) and fronto-insular cortex, as designated by Ongur and Price,
38
as important links in the “salience network” as defined by functional analyses.
39
These same areas appear to be involved in visceral self-awareness, such as the perception of heart rate, suggesting that visceral awareness contributes to evaluation of salience.
36
Visceral afferents are relayed through the fronto-insular cortex to the dACC.
38
Hyperactivity of the dACC and its associated salience network in PTSD has been postulated to contribute to hypervigilance and inappropriate reactivity to neutral stimuli in PTSD subjects.
40
Salience circuitry. Emotional tagging of sensory information occurs in part in the basolateral amygdala where sensory inputs from the thalamus and cortex are integrated with visceral afferents from autonomic centers. The amygdala relays information to the dorsolateral anterior cingulate cortex (dACC, BA24, shaded area) directly and indirectly through the thalamus and fronto-insular cortices (BA13, Iam, Iapm, Ial; shaded area); the insular cortex, especially Ia, also receives direct visceral input. Widespread efferents from the dACC then imbue salience to cognitive and motor circuitry through output projections to the striatum, thalamus, and associational cortices. Endocrine and visceral responses are conveyed through reciprocal projections to the amygdala and to subgenual anterior cingulate cortex (sgACC, BA25), an area with projections to the hypothalamus and brain stem. Frontal area designations according to Ongur and Price.
38
AB: accessory basal nucleus; B: basal nucleus; CE: central nucleus; ICM: intercalated cell masses; LA: lateral nucleus.
Classically, the dACC has been considered part of the Papez circuit, linking medial temporal, thalamic, and cortical areas in a network for emotional processing. 41 The dACC has both direct and indirect amygdala afferents, with the latter relayed through the anterior medial nucleus of the thalamus,42,43 establishing the dACC as an important structure in the mediation of emotion-guided behavior. The emotional tagging of inputs likely occurs in the amygdala through the integration of somatic sensory input with visceral input from the autonomic system. 44 Nuclei in the basolateral amygdala, in particular B and AB, relay this viscerally tagged sensory input to the dACC. 42 The dACC, in turn, is uniquely positioned to influence somatic motor, endocrine, and visceral autonomic output through projections to cortical and striatal areas, to the amygdala, and to the periaqueductal gray matter and the sgACC.45–47 The dACC itself may impart emotional salience on higher order cognitive processing via links to the mediodorsal nucleus of the thalamus, as well as connections with the dorsolateral prefrontal cortex (dlPFC) and posterior cingulate and parietal cortices.45,48,49
Mood Circuitry
Altered mood is an important aspect of PTSD that may be mediated in part by orbitofrontal and medial cortical networks (Figure 3). Postmortem and neuroimaging studies of major depressive disorder have identified the OFC, dlPFC, and cingulate cortices as pathologic sites in depression.50,51 Emphasis has been placed on the sgACC as a critical node and a target for deep brain stimulation treatment of the disorder.
51
The lateral OFC is important in integrating multi-sensory inputs related to food intake with emotional tagging from the amygdala.38,52 Thus, decision-making that is guided by emotion and the reward properties of a stimulus is thought to be a core function of orbital areas. In accordance with this functional role, the OFC projects to a ventromedial diagonal swath of the rostral caudate and putamen, notably the same striatal region that receives input from the dACC and a striatal domain that has been associated with reward and motivation.47,53,54 Although the sgACC receives limited direct projections from the dlPFC, OFC, and medial frontal cortices,
55
robust connectivity between the sgACC and dACC, an area that is densely reciprocally connected with the dlPFC and OFC, may facilitate the integration of emotion-driven sensory experiences with higher order cognitive processing. The sgACC is a major hub in the control of autonomic and endocrine function via projections to the brain stem and hypothalamus.45,46 In addition, the sgACC projects into the ventral striatopallidal loop, specifically the shell of the nucleus accumbens, which is important for goal-directed behavior, behavioral sensitization, and altered affective states.
47
Notably, the shell has extensive projections to midbrain dopamine neurons through which the sgACC can indirectly influence multiple cortical and striatal systems via dopaminergic modulation.
54
Mood circuitry. The lateral orbital frontal cortex (OFC, shaded area) receives input from virtually all senses, including those related to food intake such as gustatory and olfactory information. These sensory inputs are integrated with limbic input from the amygdala and hippocampus that impart emotional and contextual importance. The OFC influences cognitive and motor activity through outputs to the striatum, thalamus, and associational cortices as well as reciprocal connections with the amygdala and hippocampal formation. The OFC also relays information via dorsal anterior cingulate cortex (dACC, BA24) to the subgenual anterior cingulate cortex (sgACC, BA25), a major output node of the mood network that influences the autonomic system via the hypothalamus and brain stem and the reward system via the ventral striatum. Frontal area designations according to Ongur and Price.
38
AB: accessory basal nucleus; B: basal nucleus; CE: central nucleus; ICM: intercalated cell masses; LA: lateral nucleus.
Structural and Functional Abnormalities in PTSD
Frontal lobe structural MRI findings in PTSD.

F: female; M: male; NS: not specified; OFC: orbitofrontal cortex; TE: trauma-exposed control; TN: trauma-naïve control; ROI: region of interest; rPTSD: remitted PTSD; SPM: statistical parametric mapping; VBM: voxel-based morphology; PTSD: posttraumatic stress disorder; MRI: magnetic resonance imaging.
Frontal lobe functional connectivity findings in PTSD.

act: activity; amyg: amygdala; ant: anterior; anti-corr: anti-correlation compared to controls; BA: Brodmann area; BLA: basolateral amygdala; CMA: centromedial amygdala; coMDD: comorbid for major depressive disorder; corr: correlation compared to controls; CVA: canonical variates analysis; decr: decreased; F: female; fMRI: functional magnetic resonance imaging; GAD: generalized anxiety disorder; hippo: hippocampus; incr: increased correlation compared to controls; M: male; rACC: rostral anterior cingulate cortex; NAcc: nucleus accumbens; NS: not specified; SCR: skin conductance response; PCC: posterior cingulate cortex; PET: photon emission tomography; TE: trauma-exposed controls; thal: thalamus; vlPFC: ventrolateral prefrontal cortex; vmPFC: ventromedial prefrontal cortex; vs.: versus.
Structural and functional changes in the mOFC have also been reported in PTSD. In women who developed PTSD symptomatology after breast cancer surgery, the BA11 region of the OFC was smaller than that of trauma-naive controls or resilient cancer patients, and the deficits were directly correlated to the greater elevation of PTSD symptoms. 101 Because cancer- and surgery-related PTSD is a less common and sometimes controversial form of PTSD, these findings require replication in another PTSD cohort. Diminished activity in the mOFC in PTSD has been observed during trauma-related and emotionally valenced exposures.40,74,76 The mOFC is also under-activated during the performance of a memory task when emotionally laden stimuli are employed. 84 Moreover, altered functional connectivity of the mOFC with other frontal regions during executive task performance has been associated with PTSD.124,102 Resting state functional connectivity between the mOFC and the basolateral amygdala is decreased by 50% in PTSD, and connectivity is inversely associated with PTSD symptoms. 103 Functional connectivity between the amygdala and the mOFC is also decreased during viewing of emotion-evoking faces. 100 Finally, a postmortem analysis of spine density in the mOFC uncovered a shift in spine populations, resulting in a greater density of stubby spines in brains from individuals with a diagnosis of PTSD subjects compared to those without. 125 These findings suggest that neuroplasticity may be altered in frontal circuitry related to emotion processing in PTSD.
Salience circuitry is also prominently impacted in PTSD. For starters, there is near uniform consensus that dACC volume is diminished in PTSD.57,61,104–109 Meta-analyses have confirmed findings of smaller dACC volume in people diagnosed with PTSD.19,126–128 Reduced volume of the insular cortex has also been observed in PTSD.57,60,62,105,108,110 Functionally, altered activation of the dACC and insular cortex in PTSD occurs during exposure to negative imagery.40,71,75,76,79–82 During executive task performance, participants with PTSD exhibit abnormal activity in the dACC,85,88,89,94,111 as well as in the insula.90,112 In the resting state, increased connectivity of the dACC and the insula with the amygdala has been observed in PTSD,95,96 perhaps indicating an increased relay of emotionally valenced information to frontal nodes of the salience circuit. Likewise, amygdala connectivity with the dACC is enhanced in PTSD participants when listening to trauma scripts. 97 However, during performance of emotion-laden tasks, diminished connectivity among nodes of the salience circuit, i.e., the amygdala, the dACC, and the anterior insula, has been observed.98,113 Thus, the opposite directionality in findings may indicate that, in PTSD, salience circuitry is overactive at rest and underactive during some emotionally charged situations.
With regard to mood circuitry, the sgACC is implicated in PTSD pathology by findings of decreased volume of the sgACC56,62 and altered activation of the sgACC during exposure to negative imagery.68,70,71 During performance of cognitive tasks that include emotionally valenced stimuli, decreased activity has been observed in the sgACC in PTSD participants84,85,89; in contrast, increased sgACC activity has been found during fear conditioning in PTSD. 93 When PTSD participants listen to personal trauma scripts, increased functional connectivity between the amygdala and the sgACC has been described. 97 When fearful faces are viewed, decreased connectivity between the amygdala and the sgACC has been reported. 100 However, in a matching task that utilizes emotional faces, increased amygdala connectivity with the sgACC was found. 98 From a more general perspective, alterations in the structural integrity of the cingulum bundle, the uncinate fasciculus, and other frontal tracts129–135 lend further support for the premise that impoverished connectivity in frontal lobe circuitry may underlie important aspects of behavioral disturbance in PTSD. 136
Although the preponderance of evidence supports a role of the emotion-related circuitry in the frontal lobe in PTSD, some findings suggest that PTSD impacts the frontal regions associated with cognitive and executive functioning. For example, structural neuroimaging findings largely implicate the cingulate cortices and insula (Table 1); however, several studies have found smaller volume or reduced cortical thickness in individuals diagnosed with PTSD in the lateral prefrontal cortices.63,65,67,114,115 Likewise, functional neuroimaging studies have found changes in activity in the dlPFC and vlPFC in response to negative stimuli68,71,72,75,76,78,116; altered activity has even been reported in the motor cortex.126,117 Perhaps not surprisingly, abnormal activity patterns in the dlPFC and vlPFC are most often observed in participants with PTSD when they engage in cognitive tasks, particularly those involving working memory.78,84–86,88–91,118–121 One particularly enlightening study found that participants with PTSD fail to activate portions of the executive network, including the dlPFC, during a task requiring working memory. 88 At the same time, these same participants show a pattern of hyperactivity in the executive network while performing a memory-independent task that normally does not activate the dlPFC. 88 Thus, growing evidence suggests that frontal lobe disturbances in PTSD extend beyond those circuits most directly linked to emotion processing. It is well established that optimal cognitive performance is dependent on emotional status, including low stress levels, appropriate attentional focus, and emotional well-being. Indeed, the interdependence of cognition and emotional processing reflects the degree of connectivity of emotion-processing and cognitive circuits; an example is the central role that the dACC plays in salience and cognitive processing by virtue of dACC connections with the dlPFC and other higher order cortical areas.45,47–49 This relationship is further illustrated by the finding that poor neuropsychological performance in participants with PTSD has been associated with reduced functional connectivity between emotion circuitry (rACC and dmPFC) and the executive network (vlPFC). 137 The implication of this observation is that disturbances in emotion-laden processing will have ripple effects that result in the deterioration of high-level cognitive processing.
Frontal lobe functional MRI findings in PTSD.
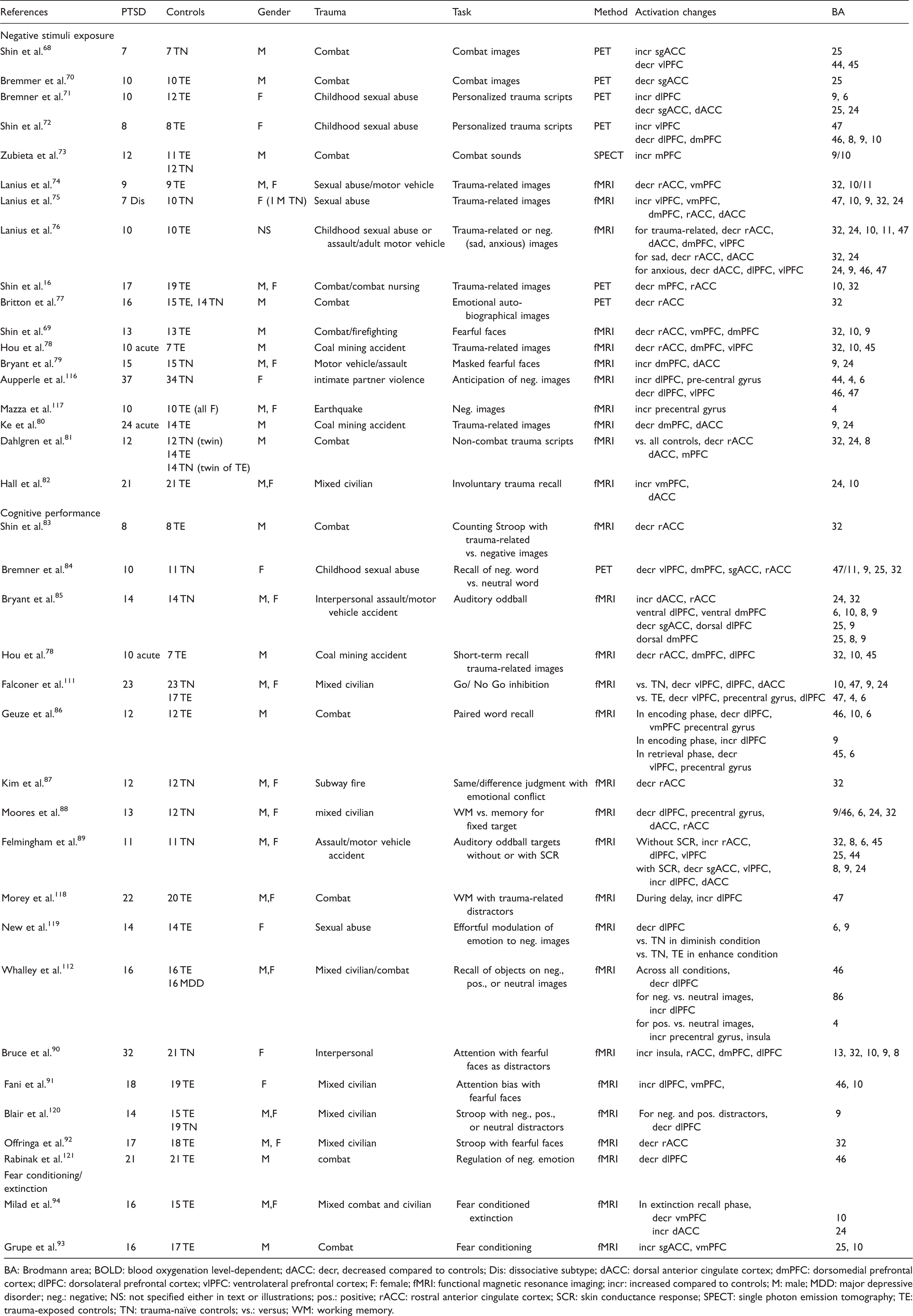
BA: Brodmann area; BOLD: blood oxygenation level-dependent; dACC: decr, decreased compared to controls; Dis: dissociative subtype; dACC: dorsal anterior cingulate cortex; dmPFC: dorsomedial prefrontal cortex; dlPFC: dorsolateral prefrontal cortex; vlPFC: ventrolateral prefrontal cortex; F: female; fMRI: functional magnetic resonance imaging; incr: increased compared to controls; M: male; MDD: major depressive disorder; neg.: negative; NS: not specified either in text or illustrations; pos.: positive; rACC: rostral anterior cingulate cortex; SCR: skin conductance response; SPECT: single photon emission tomography; TE: trauma-exposed controls; TN: trauma-naïve controls; vs.: versus; WM: working memory.
Default Mode Network Connectivity in PTSD
In recent years, the role of large-scale networks in normative cerebral function and in the dysfunction associated with psychiatric illness has come to the forefront.140,141 One such network is the default mode network, which includes prominently the posterior cingulate, parietal, and temporal cortices, and like the salience and central executive networks, the default mode network also encompasses frontal regions (BA10, sgACC, rACC, BA9, and dACC). 142 The default mode network was originally described as a “ground state” of brain oscillations such that connectivity within the default mode network needed to be overridden before other circuits were activated. 142 More recent analyses indicate that the default mode network is engaged in internal mentation, self-referential, and social processes. 143
A “triple network hypothesis” has been advanced to suggest that aberrant functioning of the default mode, salience network, and central executive network may constitute a common pathologic mechanism underlying a wide range of neuropsychological disorders. 140 With regard to PTSD, some have suggested that activity in the default mode and executive networks is suppressed by overactivity in the salience network.144,145 Several studies of functional connectivity indicate that the default mode network may be compromised in PTSD, and in particular anterior frontal portions of the network show a weakening that correlates with increased PTSD symptom severity.122,123,146,147 Furthermore, altered connectivity between the amygdala and nodes of the default mode network (rACC) and the salience network (dACC) has been described in PTSD. 95 Taken together, these studies suggest that processes related to non-task-specific mentation may be abnormal in PTSD.
Limitations
This review provides an overview of structural and functional abnormalities associated with PTSD in the context of frontal lobe circuitry that has been implicated in the disorder. One limitation of the study is that the literature reviewed may not be comprehensive. Although we attempted to include all relevant studies published between the years 2000 and 2018, it is possible that pertinent studies were missed in the literature search. In addition, this review does not assign greater weight to studies that are methodologically stronger, as for instance studies having large numbers of participants. Finally, only the functional connectivity studies most pertinent to the discussion of frontal lobe structural and functional neuro-imaging findings are included in this review. Nonetheless, we have made every effort to include literature that is representative of the field.
Moreover, this review does not address the important question of whether alterations in frontal lobe structure and function are present before the trauma exposure that results in PTSD. If abnormalities appear before trauma exposure, these alterations may predispose individuals to develop PTSD, whereas abnormalities that appear after onset of the disorder reflect pathologic changes associated with the disorder. Additional studies that examine individuals before and after trauma exposure, e.g., soldiers before and after combat deployment, are needed to address this issue.
Clinical Implications
Greater understanding of the frontal circuitry impacted in PTSD could be informative for targeting treatment of the disorder with transcranial magnetic stimulation (TMS). For example, in participants with comorbid PTSD and major depressive disorder, the strength of functional connectivity between specific regions, i.e., sgACC and the default mode network, correlates with the response to TMS stimulation of the dlPFC.148,149 Network analyses have also been useful in predicting effective psychotherapies for PTSD, including cognitive remediation, mindfulness-based stress reduction, and biofeedback. 149 The focus of this review on frontal lobe disturbances in activity and connectivity in PTSD may also facilitate development of new pharmacologic therapies to target disruptions in cortical connectivity, much as recent discovery of the novel antidepressant properties of ketamine have been validated by clinical study of glutamate signaling and prefrontal functional connectivity.150–152
Concluding Remarks
Without question, the neuroimaging of individuals diagnosed with PTSD has provided valuable insight into PTSD-related alterations in frontal lobe structural integrity and functional activity. Volumetric deficits and altered activity patterns have been observed in the emotion-processing circuitry related to fear conditioned extinction, salience, and mood in PTSD, as might be expected given the symptomatology of PTSD. Yet, it is important to note that structural and functional abnormalities extend to frontal regions that mediate cognitive and executive functioning, areas that generally have not been theoretically implicated in PTSD. We hypothesize that the disturbed functioning of emotional circuitry impacts executive networks that mediate critical functions, such as rational thinking, inhibitory control, and working memory. Moving forward, postmortem analyses of PTSD brains have the potential to probe the cellular bases of these abnormalities and deepen our understanding of PTSD pathophysiology and the mechanisms underlying this debilitating condition. The establishment of the National PTSD Brain Bank 153 (https://www.research.va.gov/programs/tissue_banking/ptsd/default.cfm) has been instrumental in providing access to quality PTSD brains for ongoing research efforts in a number of laboratories, including our own. Our group is examining spine densities on neurons in the sgACC, mOFC, and dACC in PTSD and control brains and simultaneously assessing genetic and epigenetic signatures of these selected brain regions. This work and the work by others will lead to a clearer picture of the pathological substrates of PTSD and aid in developing strategies for preventing this disease, as well as novel therapies for treatment.
Footnotes
Acknowledgment
The authors thank Dr. Leisa Glantz for careful editing of this manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Department of Defense and Veterans Affairs (5101CX001245). The funding source was not involved in interpretation of data or writing of the report. This manuscript is the result of work supported with resources and the use of facilities at the VA Connecticut Healthcare System, West Haven, CT, Central Texas Veterans Health Care System, Temple, TX and the Durham VA Medical Center, Durham, NC. The views expressed in this article are those of the authors and do not necessarily reflect the position or policy of the VA or the United States government.