
Abstract
Introduction
Adequate sleep is essential for cognitive and emotion-related functioning, and 9 to 12 hr of sleep is recommended for children ages 6 to 12 years and 8 to 10 hr for children ages 13 to 18 years. However, national survey data indicate that older youth sleep for fewer hours and fall asleep later than younger youth. This shift in sleep duration and timing corresponds with a sharp increase in onset of emotion-related problems (e.g., anxiety, depression) during adolescence. Given that both sleep duration and timing have been linked to emotion-related outcomes, the present study tests the effects of sleep duration and timing, and their interaction, on resting-state functional connectivity (RS-FC) of corticolimbic emotion-related neural circuitry in children and adolescents.
Methods
A total of 63 children and adolescents (6–17 years, 34 females) completed a weekend overnight sleep journal and a 10-min resting-state functional magnetic resonance imaging scan the next day (Sunday). Whole-brain RS-FC of the amygdala was computed, and the effects of sleep duration, timing (i.e., midpoint of sleep), and their interaction were explored using regression analyses.
Results
Overall, we found that older youth tended to sleep later and for fewer hours than younger youth. Controlling for age, shorter sleep duration was associated with lower RS-FC between the amygdala and regions implicated in emotion regulation, including ventral anterior cingulate cortex, precentral gyrus, and superior temporal gyrus. Interestingly, midpoint of sleep was associated with altered connectivity in a distinct set of brain regions involved in interoception and sensory processing, including insula, supramarginal gyrus, and postcentral gyrus. Our data also indicate widespread interactive effects of sleep duration and midpoint on brain regions implicated in emotion regulation, sensory processing, and motor control.
Conclusion
These results suggest that both sleep duration and midpoint of sleep are associated with next-day RS-FC within corticolimbic emotion-related neural circuitry in children and adolescents. The observed interactive effects of sleep duration and timing on RS-FC may reflect how homeostatic and circadian process interact in the brain and explain the complex patterns observed with respect to emotional health when considering sleep duration and timing. Sleep-related changes in corticolimbic circuitry may contribute to the onset of emotion-related problems during adolescence.
Keywords
Introduction
Sleep is an integral part of a healthy lifestyle, as sleep problems (e.g., trouble staying asleep) and sleep deprivation have been linked to emotion dysregulation and psychological problems.1,2 The link between poor sleep and emotional problems may be due to effects on processing of emotional cues or events. For example, sleep-deprived children and adults show enhanced emotional responding to negative stimuli.3–5 Importantly, average sleep duration has been steadily decreasing among Americans since the year 1985. 6 Thus, more individuals are not fulfilling their recommended sleep requirements, which could have implications for emotional health. Indeed, epidemiological studies show a steady increase in prevalence of emotion-related disorders, including anxiety and depression, over the past decade. 7 Taken together, the strong and pervasive link between poor sleep and emotional health problems warrant further investigation into underlying mechanisms.
Emerging neuroimaging studies suggest that alterations in emotion-related neurocircuitry may be a core mechanism by which poor sleep confers heightened vulnerability to emotion-related disorders. Indeed, in a landmark study by Yoo et al., 8 adults who were sleep deprived for 35 hr showed higher amygdala response and lower functional connectivity (FC) between the amygdala and ventral medial prefrontal cortex (vmPFC) during an emotion processing task, relative to a well-rested control group. The amygdala is considered a central region in emotion regulation circuitry and is responsible for filtering and prioritizing emotionally relevant information from the environment.9,10 The vmPFC and adjacent ventral anterior cingulate cortex (vACC) have direct bidirectional connections with the amygdala and are implicated in automatic (or implicit) forms of emotion regulation. 11 Positive amygdala-vmPFC/vACC FC is observed during active emotion regulation, 11 and lower amygdala-vmPFC/vACC FC is thought to reflect loss of “top-down” inhibitory control over emotional responding (i.e., emotion dysregulation)—a finding that is reported across a range of emotion-related disorders. 10 A similar pattern of reduced FC with shorter sleep has been reported in two separate studies in healthy adults, wherein sleep deprivation or shorter sleep was associated with reduced FC between the amygdala and brain regions associated with higher-order emotion regulation, including the vmPFC/vACC and also the dorsal anterior cingulate cortex/dorsomedial prefrontal cortex (dACC/dmPFC), which is implicated in more effortful (or explicit) forms of emotion regulation.1,12 Interestingly, a recent study in 16 young adult males 13 suggests that, for sleep-deprived individuals, next-day recovery of sleep reversed the effects of sleep deprivation (i.e., increased FC) on amygdala-vmPFC/vACC FC. This restoration was associated with improved self-reported mood regulation. 14 Together, these findings suggest that a night of adequate sleep restores functional connections within emotion regulation circuitry. This restoration may allow the individual to respond appropriately to social and emotional events the following day.
Importantly, studies on the effects of sleep on emotion-related neurocircuitry published to date have focused almost exclusively on sleep duration and do not consider the impact of sleep timing. These processes are thought to be distinct and interactive. Sleep duration relates to homeostatic functions that reflect levels of sleep pressure, or the need to sleep, and is based on amount of time awake.14,15 The timing of sleep relates to the circadian clock, which regulates sleep onset, awakening, and the sleep–wake cycle.16,17 The midpoint between sleep onset and offset on work/school-free days is highly correlated with circadian preferences for the timing of sleep referred to as chronotype. 18 Previous studies link a later chronotype—that is, a tendency for evening over morning activities—to emotion dysregulation in both adult and pediatric samples.19–21 To our knowledge, only one study examined the effects of chronotype on amygdala function and FC using an emotional faces task in healthy adults. 22 The authors reported higher amygdala response to fearful faces and lower amygdala-dACC FC among individuals with a later chronotype as compared to earlier. In addition, two studies reported that FC patterns within large-scale neural networks, particularly the default mode network, vary over the course of the day and depend an individual's circadian rhythm (i.e., chronotype).23,24 Together, these findings suggest that FC shows rhythmic fluctuations across the 24-hr day and that timing of sleep can also impact emotion-related brain functioning and may be relevant for the pathophysiology of emotion-related disorders. Interestingly, previous studies also link the combination of later midpoint and excessively long sleep durations to risk of depression and other emotion-related disorders.25–29 These findings suggest that there may be separate and interactive effects of sleep duration and midpoint on emotion-related outcomes; however, no studies have examined the interactive effects of these measures on emotion-related neural circuitry. Of note, it is critical that observational studies evaluate midpoint of sleep on work/school-free days, to limit interference by social (e.g., work and school) schedules. 30 However, studies on work/school-free days may be capturing weekend recovery (or “catch-up”) effects that help to cope with insufficient sleep during weekdays (or “sleep debt”). 31
Prior neuroimaging studies on sleep and emotion-related neural circuitry have focused primarily on adult samples. The link between sleep and the emotional brain is important to study in pediatric samples given (a) the sharp increase in the incidence of psychiatric problems, particularly anxiety, during the transition from childhood and adolescence,32,33 (b) the developmental shift in midpoint of sleep such that older youth naturally fall asleep later than younger youth, 34 and (c) the decline in sleep duration over the past three decades amongst school-aged youth. 35 Recommendations from the American Academy of Sleep Medicine suggest that children ages 6 to 12 years and adolescents ages 13 to 18 years require 9 to 12 hr and 8 to 10 hr, respectively, to promote optimal health. 36 Thus, it is apparent that many youth are not meeting daily sleep requirements. Additionally, a gradual decline in adolescent sleep duration has been reported over the past 20 years.35,37 Research on the impact of sleep on emotion-related neural circuitry in children and adolescents may help identify mechanisms leading to the development of anxiety.
The transition between childhood and adolescence also represents a critical period for emotional neurodevelopment. Previous studies show dramatic changes in functional activation and FC of corticolimbic brain regions involved in emotion processing and regulation. For example, relative to adults, children show higher amygdala response to negative emotional faces,38–41 which may suggest age-related decreases in emotion reactivity and/or later-emerging frontal regulatory connections. Two separate studies have documented an age-related shift in amygdala-vmPFC/vACC FC during emotional face processing tasks such that correlations were positive and then shifted to negative with age from childhood to young adulthood.38,42 This shift to negative connectivity in early adolescence (∼10–12 years) is thought to underlie the development of neural connections that are critical for emotion regulation, for example, top-down inhibitory control of vmPFC/vACC over amygdala reactivity.42,43 Immaturity and/or abnormalities in emotion-related neural circuitry may contribute to the development of emotional psychopathology.44–46
To our knowledge, only two neuroimaging studies have examined links between sleep and emotion-related neural circuitry in children or adolescents. One study in a sample of 15 male children (ages 7–11) linked shorter sleep duration to higher activation in regions involved in emotion generation, including the amygdala, insula, and temporal pole while viewing negative emotion faces. 47 Shorter sleep duration was also associated with lower amygdala-vmPFC/vACC FC during processing of emotional faces, which may reflect poorer top-down control over amygdala reactivity. 47 Given the role of vmPFC/vACC in attentional control and assessing value of stimuli or actions,48,49 effects of sleep duration on amygdala-vmPFC/vACC FC may reflect a shift of attention and/or relevant behavior away from emotionally laden stimuli. Another study found that the effects of sleep duration on neural response and FC during the processing of negative emotion faces differed in healthy (n = 19) versus anxious (n = 11) adolescents. 50 In particular, among anxious adolescents, there was a positive correlation between sleep duration and neural response to emotional faces in the dACC and hippocampus, a limbic region that is critical for emotion-related memory formation and has strong reciprocal connections with the amygdala.51,52 In healthy adolescents, in contrast, this correlation was negative. Together, these data suggest that sleep duration impacts FC within emotion-related neural circuitry in youth. However, the relatively limited sample size of these studies (n's 14–19) requires replication. These studies also focused on the effects of sleep duration on emotion-related neural circuitry and did not explore main effects or interactions with sleep timing (i.e., midpoint). One recent study examined links between chronotype on amygdala FC to emotional faces in adults 23 ; however, it is unclear if similar patterns would be observed in children and adolescents. Third, these studies examined emotion-related neural circuitry in the context of an emotion face processing task. It is unclear whether sleep affects intrinsic connectivity of this circuitry outside of an emotion processing task. For instance, resting-state FC (RS-FC) reflects the baseline organization of networks irrespective of present task demands, and may contribute to risk of later-emerging emotional problems.53,54
The present study examines the effects of sleep duration, midpoint, and their interaction on RS-FC within emotion regulation circuitry in youth. Following previous studies,8,47 we focused on RS-FC of the amygdala, given the central role of this region in emotion reactivity. We also examined sleep over the weekend, to limit interference of midpoint of sleep calculations by social (e.g., work and school) schedules. 31 However, one possible limitation of examining work/school-free days is that we may be capturing weekend recovery (i.e., catch-up) effects from insufficient sleep during weekdays (i.e., sleep debt). 32 Based on prior literature, we predicted that shorter sleep duration would be associated with lower RS-FC between the amygdala and frontal regions such as the vmPFC/vACC and orbitofrontal cortex, and parietal regions such as inferior parietal cortex. In addition, we predicted that a later midpoint of sleep would be associated with lower RS-FC between the amygdala and regions of the brain implicated in regulating the circadian clock such as the posterior cingulate cortex, vmPFC/vACC, and angular gyrus. Given that previous studies have also found that the combination of later midpoint and excessively long sleep durations predict risk of depression and other emotion-related disorders,24–28 we predicted interactive effects of sleep duration and midpoint on amygdala RS-FC, for example, in areas of overlap (e.g., vmPFC/vACC). Further, given that shorter sleep duration and later sleep timing have both been linked to poor emotional outcomes, we expected to observe similar findings in our sample and that these changes would be mediated by RS-FC. Importantly, we tested these links in a racially and economically diverse sample of youth, consistent with calls for greater generalizability in neuroimaging studies 55 and for more research that can address racial, ethnic, and socioeconomic disparities. 56 Our sample included youth from lower income households and who endorsed exposure to threat-related adversity (e.g., violence, medical-related adversity), which are known risk factors for emotional psychopathology. 57 Thus, we tested for potential links among duration and midpoint of sleep, and anxiety and depressive symptom severity.
Materials and Methods
Participants
Demographic and emotional health data (N = 63).

Note: KBIT-2: Kaufman Brief Intelligence Test, Second Edition; CDI-S: Children's Depression Inventory, Short Form; SCARED: Screen for Child Anxiety-Related Emotional Disorders.
Ten children reported more than one type of adversity exposure.
Participants were recruited through local advertisements and healthcare providers. All children spoke English as a first language. Children with history of brain injury, neurological or movement disorders, or magnetic resonance imaging (MRI) contraindications were excluded from the study. Consistent with the study goals of (a) identifying neural correlates of sleep-related emotional disturbance and (b) achieving a more representative sample of our study setting (Metro Detroit, Michigan), we did not screen out children for current or past diagnosis of anxiety or mood disorder. Rather, we selected our sample to include a subset of youth at high risk of emotion-related disorders, and indeed, a subset of youth endorsed high levels of anxiety and/or depressive symptomology (see later). Participants and parents provided full written consent or assent as appropriate, and all study methods were approved by the institutional review board of Wayne State University (WSU).
Measures of Sleep Duration and Emotional Psychopathology
Sleep
This study was performed over the weekend (Saturday and Sunday) to more accurately capture individual variation in sleep timing (i.e., midpoint) with minimal interference from extrinsic factors, such as school start times. In addition, previous studies indicate shorter sleep duration in youth during the school year in comparison to vacation days.58,59 However, to compensate for sleep debt accumulated during the week, youth may oversleep on weekends, which may bias measures of sleep duration on weekend days (see Discussion section). At the end of Day 1 (Saturday), children and their parents/guardians were given a sleep packet to be completed together at home. Sleep duration and midpoint of sleep the night before their scan (on Day 2; Sunday) were estimated from child/parent reports of sleep and wake-up time. Participants were instructed to complete the sleep packet immediately when they woke up the morning of their scan (Sunday). Five children had times missing for bed time and/or wake-up time, and children with versus without missing data did not differ in age, sex, race/ethnicity, IQ, depression, anxiety, income, or threat-related adversity exposure (ps > 0.08). Thus, to maximize sample size, missing values were imputed using the sample mean. Results were consistent with those obtained when excluding participants with missing values and for additionally controlling for participants with missing data.
Anxiety and Depression
On Day 1, children completed standardized age-appropriate measures of depression (Children's Depression Inventory, Short Form (CDI-S))49,60 and anxiety (Screen for Child Anxiety-Related Emotional Disorders (SCARED)). 50 Of note, 48% of participants exceeded thresholds suggested for detecting pathological anxiety (SCARED > 22) 61 and 37% for depression (CDI-S ≥ 3). 62 Overall, 60% of participants exceeded thresholds for anxiety and/or depression (see Table 1). Therefore, although diagnostic testing was not performed here, these standardized measures suggest a significant number of youth at risk for emotional psychopathology. Of note, the SCARED is comprised of five anxiety subtypes: generalized anxiety, separation anxiety, panic symptoms, social anxiety, and school avoidance. 63 We examined links between sleep measures and depression and total anxiety scores and considered anxiety subtypes in exploratory analyses.
MRI Acquisition Parameters and Analysis
On Day 2 (Sunday), participants underwent a 10-min eyes-closed resting-state functional MRI (fMRI) scan. A multiecho multiband sequence was used for fMRI data collection, and an anatomical image was also collected during each scan session, for anatomical-functional coregistration. Of note, the experimenters checked in with the participants after the resting-state scan and noted that four children fell asleep during the scan. To ensure that these participants did not bias our results, we re-ran FC analyses controlling for whether the participant fell asleep during the scan. Results of these additional analyses are given in the Supplemental Material and are consistent with our reported results. The four participants who fell asleep during the scan did not differ on prescan measures of sleep duration or midpoint, or in demographic factors (i.e., age, sex, race/ethnicity, ps > 0.1). FMRI preprocessing and denoising was conducted using multiecho independent components analysis (ME-ICA) software (v3, beta 1; https://bitbucket.org/prantikk/me-ica) and included, in brief, skull-stripping, coregistration of first echo time series and anatomical-functional coregistration, de-obliquing, and removal of the first 15 s to account for signal equilibration. ME-ICA was used to remove likely non-BOLD artifact from the data, and remaining participant motion (as captured using DVARS) 64 was controlled for, when appropriate, in group-level analyses. RS-FC of left and right amygdala was computed for each participant. Group-level analyses included three separate whole-brain regression analyses, each controlling for age: (a) sleep duration as the regressor of interest, (b) midpoint of sleep, and (c) the interaction between sleep duration and midpoint of sleep. Results were considered significant using a whole-brain voxel and cluster level corrected threshold. See Appendix A in Supplemental Material for further details.
Results
Effects of Demographic or Clinical Measures on Sleep Duration
Reported sleep duration ranged from 5.5 to 12.7 hr (M = 9.81, SD = 1.39). Of note, a majority (72%) of participants met recommendations for sleep duration outlined by the American Academy of Sleep Medicine
37
; 10% of the sample slept longer than recommended, and 17% slept shorter. Specifically, in children ages 6 to 12 years, 15% of participants slept shorter, and 10% slept longer than recommended. For children between the ages of 13 to 18 years, 30% slept shorter, and 10% slept longer. There was no association between sleep duration and midpoint of sleep (p = 0.24). As shown in Figure 1 and consistent with previous reports in youth,35,65–67 older youth tended to sleep for shorter durations, r(63) = −0.404, p = 0.001, and had a later midpoint of sleep, r(57) = 0.357, p = 0.006. Total anxiety scores, depression, adversity exposure, and income were not related to sleep duration or sleep midpoint, ps > 0.05. However, when examining anxiety subtypes, longer sleep duration was associated with greater separation anxiety symptoms, r(41) = 0.35, p = 0.026. Although there were no sex differences in midpoint of sleep, there were sex differences in sleep duration such that females reported sleeping longer than males, t(61) = 2.2, p = 0.032. These sex differences are consistent with previous studies in youth samples.68–70 Nonetheless to ensure that observed results are not biased by sex or adversity exposure, we performed additional fMRI analyses controlling for these variables. These results are consistent with our main analyses and are reported in Supplemental Tables S15 to S26. In addition, to ensure that results are driven by effects of sleep rather than anxiety or depressive symptoms, we performed two additional whole-brain regression analyses entering anxiety and depressive symptoms as regressors of interest in separate models (controlling for age). Observed effects were independent of those observed for sleep parameters and are presented in the Supplemental Material (see Tables S13 and S14).
Older youth tended to sleep for shorter durations (a) and had a later midpoint of sleep (b). Shaded areas indicate recommended sleep duration based on age, according to the American Academy of Sleep Medicine.
37

Effects of Sleep Duration on Amygdala RS-FC
Figure 2 shows areas of increasing (red) and decreasing (blue) RS-FC with longer sleep duration for left (a) and right (b) amygdala. Longer sleep duration is associated with higher amygdala RS-FC with several regions that span the cerbellum, as well as frontal, temporal, parietal, and occipital lobes (see Table S1 for complete list). Of note, regions showing higher amygdala RS-FC with longer sleep durations included superior temporal gyrus, precentral gyrus, and vACC. In contrast, longer sleep duration was associated with lower amygdala RS-FC, with a variety of regions, including cuneus, inferior frontal gyrus, insula, and thalamus (see Table S2).
Effects of sleep duration on resting-state functional connectivity of the right and left amygdala. Longer sleep is associated with higher amygdala connectivity with the regions shown in red and lower amygdala connectivity with the regions shown in blue. Results significant at a whole-brain corrected threshold (p < 0.001, 9 voxels) and shown at p < 0.005, 20 voxels for display purposes. Regions circled for left amygdala include middle cingulate cortex, dorsal anterior cingulate cortex, and superior temporal gyrus. Regions circled for right amygdala include dorsal anterior cingulate cortex, inferior frontal gyrus, thalamus, and midbrain.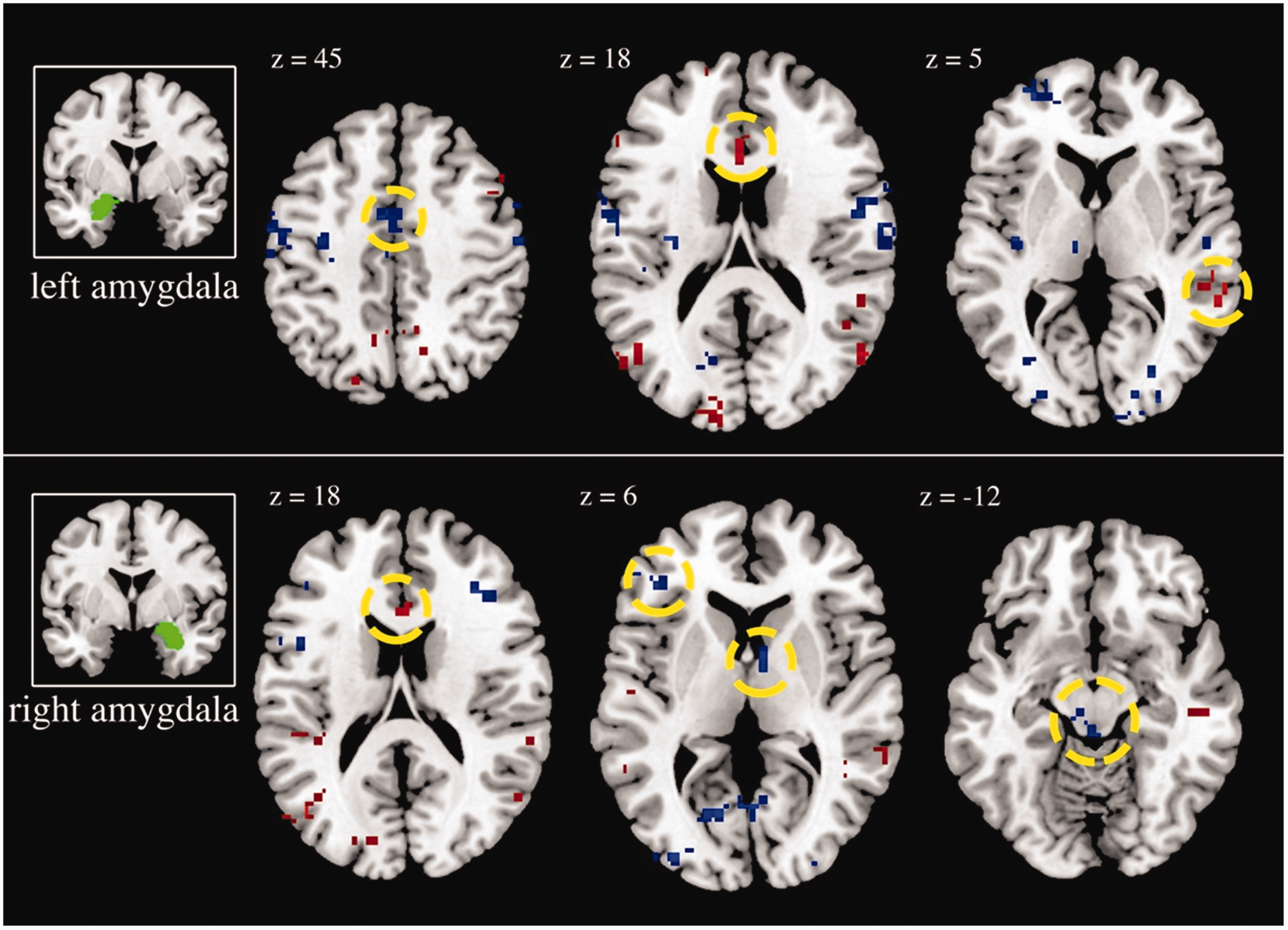
Effects of Midpoint of Sleep on Amygdala RS-FC
Figure 3 shows areas of higher (red) and lower (blue) RS-FC with a later midpoint of sleep for left (a) and right (b) amygdala. Later midpoint was associated with higher amygdala RS-FC with several regions (see Table S3), including the insula, dmPFC/dACC (Brodmann Area (BA) 9), cerebellum, supramarginal gyrus, postcentral gyrus, and superior frontal gyrus. Lower amygdala RS-FC was observed for later midpoint in the postcentral gyrus, middle temporal gyrus, and the cerebellum (Table S4).
Effects of midpoint of sleep on resting-state functional connectivity of the right and left amygdala. Later midpoint of sleep is associated with higher amygdala connectivity with the regions shown in red and lower amygdala connectivity with the regions shown in blue. Results significant at a whole-brain corrected threshold (p < 0.001, 9 voxels) and shown at p < 0.005, 20 voxels for display purposes. Regions circled for left amygdala include inferior parietal lobe, postcentral gyrus, and insula. Regions circled for right amygdala include inferior parietal lobe, middle cingulate cortex, and cerebellum.
Interactive Effects of Sleep Duration and Midpoint of Sleep on Amygdala RS-FC
Figure 4 shows areas of increasing (red) and decreasing (blue) RS-FC with a longer sleep duration and later midpoint of sleep for left (a) and right (b) amygdala. The combination of longer sleep duration and later midpoint is associated with altered amygdala RS-FC in a strikingly large number of regions (see Table S5). Of note, higher RS-FC was observed in vACC as well as dmPFC/dACC, superior temporal gyrus, precuneus, cerebellum, and insula. Lower amygdala RS-FC was observed in a similar set of brain areas, including supplemental motor area, vACC, dmPFC/dACC, precuneus, superior temporal gyrus, cerebellum, and insula (Table S6).
Interactive effects of duration and midpoint of sleep on resting-state functional connectivity of the right and left amygdala. The combination of longer sleep duration and later midpoint of sleep is associated with higher amygdala connectivity with the regions shown in red and lower amygdala connectivity with the regions shown in blue. Results significant at a whole-brain corrected threshold (p < 0.001, 9 voxels) and shown at p < 0.005, 20 voxels for display purposes. Regions circled for left amygdala include superior frontal gyrus, medial frontal gyrus, and inferior frontal gyrus. Regions circled for right amygdala include superior frontal gyrus, medial frontal gyrus, and inferior frontal gyrus.
Discussion
To our knowledge, this is the first study to examine the effects of sleep duration, midpoint of sleep, and their interaction on emotion-related neural circuitry in a pediatric sample. In line with previous studies,35,65–67 we found that older youth tended to sleep later and for fewer hours than younger youth. Also consistent with previous studies in adult and pediatric samples,13,47,71,72 we found that longer sleep duration was associated with higher RS-FC within emotion-related neural circuitry, including amygdala RS-FC with vACC, precentral gyrus, and superior temporal gyrus. Interestingly, midpoint of sleep was associated with altered connectivity in a distinct set of regions involved in interoception and sensory processing, including insula, supramarginal gyrus, and postcentral gyrus. Our data also indicate widespread interactive effects of sleep duration and midpoint on brain regions involved in emotion regulation, sensory processing, and motor control. These patterns may reflect how homeostatic and circadian process interact in the brain and explain the complex patterns observed with respect to emotional health when considering sleep duration and timing. 73 Of note, these patterns were observed outside the context of an emotional task and thus add to previous task-based studies by showing that sleep duration and timing affect integrity of emotion-related neural circuitry both inside and outside of specific emotion processing tasks (e.g., emotional faces). Effects of sleep on intrinsic FC may therefore influence a range of neurobehavioral processes (e.g., attention, emotion regulation). Taken together, these data may provide new insights on the well-established link between poor sleep and the emergence of emotion-related problems (e.g., anxiety, depression) during the transition from childhood to adolescence.
Our whole-brain analyses on sleep duration are consistent with previous RS-FC studies in healthy adults. Previous literature shows that, compared to sleep-deprived individuals, well-rested individuals show higher amygdala RS-FC with various regions, including vACC, superior temporal gyrus, middle temporal gyrus, and inferior frontal gyrus. Higher coupling between the amygdala and these regions has been observed during active up- and downregulation of emotion 74 and may suggest that sleep increases the ability to voluntarily up- and downregulate emotional responses. Midpoint of sleep correlated with amygdala RS-FC in a separate set of regions, suggesting distinct effects of sleep duration and timing on emotion-related neurocircuitry. Later midpoint of sleep was associated with higher amygdala RS-FC with the insula, supramarginal gyrus, and postcentral gyrus. Interestingly, previous studies have linked higher amygdala RS-FC with the insula75,76 and supramarginal gyrus 77 among adolescents or adults with major depressive disorder. Given the observed links between midpoint of sleep and depression,78,79 the effects of later midpoint of sleep on amygdala connectivity with brain regions involved in interoceptive awareness and emotion regulation (e.g., insula) may reflect risk of a depressive phenotype. Of note, we observed higher amygdala-dmPFC/dACC FC response in children with a later midpoint of sleep. Interestingly, this pattern differs from a previous study in adults showing lower amygdala-dACC FC during fearful face processing in adults with a later chronotype. 23 Lower amygdala-dACC FC has been observed in depressed adults and never-depressed at-risk populations (e.g., family history of depression), 77 fitting with the role of this region in cognitive regulation of limbic regions and downregulation of negative emotional responses.80,81 It is possible that the observed discrepancies are due to effects of task versus resting state or that the effects of sleep on amygdala-dmPFC/dACC circuitry differ in adults than in children and adolescents, whose frontolimbic circuitry and cognitive control over emotion responding may be immature.82,83 Alternatively, more rostral regions of the dmPFC/dACC have been implicated in the conscious, negative appraisal of threatening situations, 84 and higher amygdala-dmPFC/dACC FC has been observed during fear expression.81,85 Thus, higher amygdala-dmPFC/dACC FC observed in youth with later midpoints of sleep may reflect a bias toward negative appraisals of situations and greater coordination within fear neural circuitry. We additionally observed higher amygdala-dmPFC/dACC RS-FC for youth who both slept longer and later (i.e., sleep duration × midpoint interaction). Thus, amygdala-dmPFC/dACC circuitry, at least in childhood and adolescence, may relate to the interaction between homeostatic and circadian processes.
Although we are the first to link sleep timing (i.e., midpoint of sleep) to emotion-related neural circuitry in a pediatric sample, previous literature has explored the associations between chronotype and behavioral and self-report measures of emotion regulation. Previous studies in adolescents link later chronotypes (or “eveningness”) to poorer self-reported emotion regulation, 86 greater self-reported emotional and behavioral problems, 87 and lower mood. 88 These data suggest that chronotype has an effect on emotional regulation and mood, and, as a result, we would expect to observe altered RS-FC within emotion-related neural circuitry. Our results support this hypothesis because youth with a later midpoint of sleep showed decreased amygdala RS-FC with the postcentral gyrus and middle temporal gyrus. RS-FC between the amygdala and postcentral gyrus has been implicated in predicting emotion regulation skills in school-aged children and adults. 89 Further, in an emotional face processing task, healthy adults who had increased coupling of the amygdala with the middle temporal gyrus showed appropriate upregulation of emotion. 74 We also found that later midpoint was associated with higher RS-FC between the amygdala and the insula, a region that is critical for emotional awareness and interoception. The insula also has strong reciprocal connectivity with hypothalamic and brainstem regions involved in the sleep–wake cycle.90,91 Connections between brain regions involved in the sleep–wake cycle and emotion regulation may reflect changes in circadian patterns observed in those with emotion-related disorders. Midpoint-related changes in emotion-related neural circuitry during childhood may contribute to the development of psychopathology.
Our analyses show a strikingly larger number of regions associated with the interaction between sleep duration and midpoint than for either sleep parameter alone. These findings suggest that the interactive effects of sleep length and timing may impact emotion-related neurocircuitry and subsequent outcomes and that a more nuanced view of the relationship between sleep and emotion regulation is required. Indeed, among adults, the combination of longer and later sleep is associated with increased risk of depression and other psychiatric disorders.27,92,93 In this study, we found lower amygdala RS-FC connectivity with brain regions implicated in emotion regulation—including the superior temporal gyrus,94,95 vACC,92,96 and vmPFC97,98—among youth who slept longer and later. Moreover, the observed interactive effects of sleep duration and midpoint is relevant for a conceptual framework proposed by Gruber and Cassoff 73 that explores the intersection of sleep and emotion regulation. In that review, the authors suggest that unsuccessful emotional regulation is caused by inappropriate emotional reactivity, which is influenced by sleep duration and altered connectivity between the amygdala and prefrontal regions. Frontoamygdala circuitry is governed by homeostatic process that relates to sleep length. The authors also suggest that misalignment of circadian processes (e.g., later chronotype) can increase the likelihood of emotional reactivity via mood dysregulation. Therefore, youth in our sample may be experiencing the combined effects of poor mood regulation, due to later chronotype, and altered RS-FC between the amygdala and emotionally relevant brain areas due to shorter sleep duration. Furthermore, it may be beneficial to expand this framework to include emotion regulation brain regions outside of prefrontal areas.
Interestingly, alterations in amygdala RS-FC with the cerebellum were consistently observed across sleep duration, midpoint, and interaction results. For example, children who slept for longer periods of time and had a later midpoint of sleep showed positive and negative effects on amygdala RS-FC with the cerebellum. The cerebellum, commonly known for its role in motor control and coordination,99,100 is also implicated in emotion processing and regulation. 101 For example, a study by Turner et al. 102 found that lesions in the cerebellum were associated with altered emotion-related responding to frightening stimuli. The authors suggested that the cerebellum, which coordinates closely with prefrontal and cingulate regions, is a key brain region that determines how an individual processes and regulates emotionally laden stimuli. 102 Importantly, both higher and lower RS-FC between the amygdala and cerebellum have been reported in previous studies of adolescents with generalized anxiety disorder.103,104 Thus, disruptions in amygdala-cerebellar circuitry may contribute to the development of adolescent anxiety.
Surprisingly, sleep measures were not associated with anxiety or depression symptom severity in our sample. Previous studies have reported higher anxiety and depression with shorter sleep duration.105,106 It is possible that effects of sleep on emotion-related health may not become apparent until later in adolescence and may be driven by an array of hormonal, social, and neurobiological changes that take place during this time.107,108 Our exploratory analyses in anxiety subtypes show that longer sleep duration was associated with increased separation anxiety symptoms. Separation anxiety is a common subtype of anxiety in childhood and has been shown to mediate the link between childhood adversity and subsequent onset of common psychiatric disorders in adulthood. 109 Separation anxiety is also prevalent among low-income samples, 110 which is consistent with our sample (see Table 1).
While this study expands upon prior literature in a number of ways, including a larger sample size, inclusion of sleep timing measures, and a socioeconomically diverse sample, there are some important limitations. First, our estimates of sleep were based on one night of sleep, prior to their MRI scan. To more closely capture an individual's sleep behavior, future studies should measure sleep duration and patterns over a longer period of time to characterize habitual sleep duration and timing which would account for the variability in these parameters and the sleep debt accrued prior to brain imaging. Prospectively collected sleep data including sleep diaries and wrist actigraphy over a longer period would help elucidate these effects and address this limitation. In addition, it would be important to rule out sleep pathology (e.g., sleep-related breathing disorders and insomnia) which affect sleep duration, timing of sleep, and sleep quality. Second, self- and parent-reported sleep measures do not always correlate with more objective measures of sleep such as polysomnography, actigraphy, 111 cortisol, body temperature, or melatonin, 23 and the inclusion of these variables would help to clarify the mechanisms associated with the aforementioned associations between sleep duration and timing. Third, the present study focused on overall duration of sleep. Future studies may explore the effects of the variability in the quality and quantity of sleep architecture associated with slow-wave sleep and rapid eye movement sleep on emotion-related neurocircuitry and health outcomes. Finally, because we measured sleep and FC on weekend days, we are unable to examine the potential impact of sleep debt on our results. That is, longer sleep durations may reflect restorative sleep due to the accumulation of sleep debt throughout the week, due to a misalignment between the child's chronotype and school timings (“social jetlag”). 31
Conclusions
The present study demonstrates distinct and interactive effects of sleep duration and timing (i.e., midpoint) on intrinsic FC of emotion-related neural circuitry in youth. Alterations in emotion-related neural circuitry may help to explain the strong and pervasive link between poor sleep and the emergence of emotion-related disorders, many of which emerge in the first two decades of life. Research on mechanisms underlying the link between poor sleep and emotional health should have implications for school timing and mental health. For example, the documented detrimental effects of social jetlag112,113 may be reduced by the implementation of school times that are aligned with the sleep rhythms of children and adolescents.
Supplemental Material
Supplemental material for Effects of Duration and Midpoint of Sleep on Corticolimbic Circuitry in Youth
Supplemental Material for Effects of Duration and Midpoint of Sleep on Corticolimbic Circuitry in Youth by Aneesh Hehr, Hilary A. Marusak, Edward D. Huntley and Christine A. Rabinak in Chronic Stress
Footnotes
Acknowledgments
The authors would like to thank Craig Peters, Allesandra Iadipaolo, Farrah Elrahal, Limi Sharif, Shelley Paulisin, Pavan Jella, Kelsey Sala-Hamrick, Laura Crespo, Xhenis Brahimi, Klaramari Gellci, and Suzanne Brown of WSU for assistance in participant recruitment and data collection. The authors would like to thank Dr. Moriah Thomason for sharing some of the included data. Thanks also to the children, adolescents, and their families who generously shared their time to participate in this study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research reported in this publication was supported by the WSU Department of Pharmacy Practice, a Karmanos Cancer Institute and American Cancer Society #14-238-04-IRG (CAR), and the WSU Undergraduate Opportunities Research Program. Dr Marusak is supported by American Cancer Society award 129368-PF-16-057-01-PCSM. Dr Rabinak is supported by National Institute of Mental Health grants K01 MH101123 and R61 MH111935.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.