
Abstract
Post-traumatic stress disorder (PTSD) is a disabling psychiatric condition that can develop following a physical, psychological, or sexual trauma. Despite the growing body of literature examining the psychological and biological factors involved in PTSD psychopathology, specific biomarkers that may improve diagnosis and treatment of PTSD have yet to be identified and validated. This challenge may be attributed to the diverse array of symptoms that individuals with the disorder manifest. Examining the interrelated stress and fear systems allows for a more comprehensive study of these symptoms, and through this approach, which aligns with the research domain criteria (RDoC) framework, neural and psychophysiological measures of PTSD have emerged. In this review, we discuss PTSD neurobiology and treatment within the context of fear and stress network interactions and elucidate the advantages of using an RDoC approach to better understand PTSD with fear conditioning and extinction paradigms.
Keywords
Introduction
Post-traumatic stress disorder (PTSD) is an incapacitating psychiatric condition that some individuals develop after experiencing stressful or traumatic life events. According to the Diagnostic and Statistical Manual of Mental Disorders, 5th Edition (DSM-5), PTSD is diagnosed based on clusters of symptoms, such as the re-experiencing/having intrusive memories of the trauma, avoidance, negative cognition and mood, and hyperarousal. 1 PTSD afflicts 7% to 8% of the adult population in the United States, 2 and despite the many efforts to understand the pathophysiology of the disease, improved diagnosis and treatment options are still needed. The heterogeneous and complex nature of PTSD may contribute to the difficulty in achieving these improvements, and one approach to addressing this challenge is through the research domain criteria (RDoC) framework.3,4 Through this perspective, we are able to envision PTSD in a way that is more comprehensive, not limited by subjectively reported symptoms or DSM-5 criteria, and inclusive of neurobiological and physiological measures.
One underlying transdiagnostic feature of PTSD and other anxiety disorders is fear dysregulation. Accumulating evidence has demonstrated abnormalities in the brain circuits that mediate fear responses, and these abnormalities are associated with PTSD.5–8 Both preclinical and clinical data implicate the role of stress and fear circuit interactions in the development, maintenance, and treatment of PTSD. In this review, we examine the neurobiology of PTSD and its intersection with stress and fear response systems. Additionally, we discuss the advantages of using an RDoC approach (supplied by fear conditioning and extinction paradigms) in combination with the DSM-5 to improve our understanding and treatment of PTSD.
Neurobiology of Fear and PTSD
Individuals with PTSD have great difficulty regulating their fear of trauma-associated stimuli, making the study of fear integral to fully understanding the disorder.5,7,9–11 The behavioral and neurobiological responses to fear are critical to this understanding and have been investigated extensively using a paradigm dependent on a Pavlovian learning process called fear conditioning. During this type of learning, a subject is presented with a neutral stimulus (conditioned stimulus), which is typically a light, tone, image, or context. This neutral stimulus is paired with an unconditioned stimulus, which is usually a shock. As the association between the two stimuli is learned, the subject expresses a response that is called the conditioned response. In rodents, the conditioned response when measuring fear is usually freezing behavior or the fear-potentiated startle reflex. In humans, it is typically a psychophysiological index of fear including measures such as fear potentiation of the acoustic startle reflex,12,13 skin conductance response (SCR),5,14–16 or heart rate.17,18 Healthy individuals learn to reduce their fear response to the conditioned stimulus in the absence of a threat. The failure to reduce fear in this way often leads to the development of PTSD psychopathology.10,19–21
Psychophysiological measures have been valuable to the study of fear processing and the neurobiology and symptoms of PTSD.5,22–25 In one example of this, combat veterans, who were subthreshold for a full PTSD diagnosis but did express PTSD symptoms, showed abnormal psychophysiological responses to fear (i.e., heart rate, SCR, startle, and respiratory rate) that were similar to those with a full PTSD diagnosis. 17 This suggests that despite not having a PTSD diagnosis, many individuals may still need treatment for the symptoms of PTSD and benefit from additional psychophysiological measures. Using fear paradigms in this way provides a venue in which specific attributes of PTSD, such as impaired safety learning, amplified fear responses to trauma- or stress-related stimuli, and fear generalization can be distinctly investigated. Moreover, these fear paradigm-centered findings support the idea that shifting focus towards the symptoms rather than relying solely on DSM diagnoses may foster more insight and effective treatments for PTSD. Considering this and the broad range of applications for studying fear, examining the neural correlates of fear in PTSD (or PTSD symptomatic but subthreshold) individuals may provide insight into the neurobiological mechanisms underlying the disorder.
Much of what we know about human fear neurocircuitry has correlates in rodents, enabling many preclinical findings to be translatable to human studies. The fear network has been extensively studied in rodents, with the prelimbic (PL) and infralimbic (IL) areas of the medial prefrontal cortex (mPFC), specific nuclei of the amygdala, and the hippocampus identified as important mediators of fear-related processes.26–32 One study delineating the roles of these regions in fear acquisition, extinction, and memory retrieval found that inactivation of the PL before extinction training reduced fear expression during early extinction training (where one would see evidence of fear memory) but had no effect on extinction memory. 30 IL inactivation prior to extinction training left the fear memory intact, while extinction learning and memory were impaired. 30 Moreover, stimulation of the PL during tone presentations increased fear expression and impaired extinction learning, whereas IL stimulation reduced freezing to the conditioned tone during extinction. 33 Together, these findings suggest that PL activity is necessary to drive the expression of conditioned fear, and IL activity is important for its inhibition.
In addition to identifying these critical brain regions, numerous rodent studies have also revealed the role of specific cell types and synchrony of activity or connections between the regions. While the amygdala’s contributions to fear processes have long been recognized, further investigations have delineated specific microcircuits within its nuclei. For example, interactions between the lateral and central divisions of the amygdala have been shown to increase and decrease fear output based on the centrolateral amygdala’s inhibitory control of the centromedial amygdala. 34 The connections between the amygdala and the prefrontal cortex have also been found to be critical in regulating fear responses.28,35–38 For example, rats exhibiting high anxiety-related behaviors showed impaired extinction with abnormal activations within the prefrontal-amygdala circuit (reduced prefrontal activity and increased centromedial amygdala activity). 39 During safety learning, theta oscillation synchrony within this circuit also seems to modulate fear responding.38,40 Theta oscillations are neural signals that oscillate at a frequency range of 4 to 12 Hz, and synchrony of these oscillatory patterns between brain areas is important for learning and indicates communication between the regions during specific learned behaviors and/or stimuli presentations. 40 Increased synchrony of theta oscillations between the mPFC and the basolateral amygdala was found to modulate discrimination between safety and fear learning, suggesting that basolateral amygdala responses (and their timing) to inputs from the mPFC determine increases and reductions in fear behaviors. 38
Fear conditioning and extinction studies in humans support the translatability of these preclinical findings, identifying critical nodes of the fear network, including the ventromedial prefrontal cortex (vmPFC), amygdala, dorsal anterior cingulate cortex (dACC), insula, and hippocampus (Figure 1).5,41–43 Brain activations within the dACC (homologous with the rodent PL) appear critical during fear acquisition, whereas vmPFC (homologous with the rodent IL) activation plays an important role in fear extinction.5,44 Additional brain regions, including parts of the cerebellum, insula, thalamus, and striatum, have also been found to be significantly activated during fear acquisition.43,45 As these regions are not only involved within the threat detection and fear processing circuitry, these findings suggest other networks should be investigated as well.
Summary of brain activations commonly considered the fear network regions. In healthy subjects, the dACC, insula, and amygdala exhibit robust activations during fear acquisition. During fear extinction, the AMY and vmPFC show increased activations. The vmPFC and hippocampus are activated during extinction recall.
There is accumulating evidence indicating that individuals with PTSD have difficulty regulating their fear and learning safety signals.6,7,10,19–21,46–52 Studies have shown extinction learning and memory recall to be significantly impaired in patients with PTSD compared to trauma-exposed non-PTSD control participants.5,52,53 Milad et al. 5 found that the impairment was restricted to the recall of the extinction memory, whereas conditioned fear acquisition and extinction learning were not disrupted in the PTSD group. While studies such as these report deficits in only the memory of extinction or extinction retention, other studies have also reported deficits in the learning of fear inhibition.8,48,54,55 For example, PTSD subjects exhibited greater fear-potentiated startle responses to the extinguished cue during extinction, showing an impairment in learning to extinguish their fear, in addition to heightened startle during fear conditioning. 48
PTSD and anxiety literature also describe an inverse relationship between activations within frontal regions and the amygdala, i.e., vmPFC hypoactivation and amygdala hyperactivation; this suggests a loss of prefrontal control of the amygdala, which would normally regulate fear responses.7,56–58 Milad et al. 5 reported that the extinction memory impairment in PTSD patients was associated with significant deactivation in the vmPFC and heightened activation within the dACC, suggesting less prefrontal inhibition, but more excitation, of the amygdala. A meta-analysis reported that reduced activation of the vmPFC was correlated with increased activation of the amygdala in PTSD patients, 59 indicating the consistency of this brain activation pattern in the fear network in PTSD. Thus, alterations in neural connectivity or communication between these brain regions due to disproportionate activation may underlie some PTSD symptoms.57,60,61 In fact, amygdala hyperreactivity and differential cortical activity (top-down processing regions) in response to fearful versus neutral face stimuli was found to be predictive of the perseverance of PTSD symptoms. 61 This was also observed for symptom severity in individuals below threshold for a PTSD diagnosis.61,62 Higher Clinician-Administered PTSD Scale scores appeared to be associated with reduced amygdala-vmPFC connectivity, suggesting that communication between these two structures may be compromised with more severe cases of PTSD. 63 Moreover, structural differences in these areas have been reported in PTSD; reduced cortical thickness in the frontal lobes and hippocampal volume of soldiers with PTSD compared to those without PTSD were observed.64,65 Given that daily life stressors and other triggering stimuli are often unavoidable, the altered neurocircuitry in PTSD creates an ongoing inability to control fear. This is also true in instances when exposure to stress is prolonged and becomes chronic stress, which has been shown to exacerbate PTSD symptoms. 66
Chronic Stress, Fear, and PTSD
Fear is an adaptive threat response, and as such, it is important for survival. A certain level of stress can be similarly beneficial, promoting alertness and energy in situations when a quick reaction is necessary. 67 When a stress response is activated in these situations, it triggers components mediating the “fight-or-flight” response. These include the locus coeruleus/norepinephrine (NE) system, the corticotropin-releasing factor/hypothalamic-pituitary-adrenal (HPA) axis, as well as other neurobiological systems.68,69 Traumatic or chronic stress, which is similar to an exaggerated fear response that cannot be turned off, can alter functioning of these systems (often increasing responsivity to future stressors). This may contribute to PTSD symptoms and have serious consequences for mental and physical health.70–74 Chronic stress refers to not only normally occurring, daily life stressors and any subsequent trauma experienced after a PTSD-inducing event but also the repeated stress caused by re-experiencing trauma through recurring, intrusive thoughts, and/or nightmares. Prior traumatic stress exposure is a prerequisite for PTSD diagnosis according to DSM-5 criteria, but persistent alterations in the neurobiological mechanisms mediating the stress response also appear to underlie many PTSD symptoms,70,75,76 suggesting that consequent stressors following the diagnosis are also critical contributors to the disease. As the type of initial trauma can affect PTSD symptomatology, it may also be important to account for the type or source of chronic stress exposure in PTSD research.
The literature on PTSD consistently reports increased NE and dysregulation of the HPA axis, a neurobiological system that modulates stress hormones and reactions to trauma, in individuals with PTSD.77–80 Compared to healthy controls, the pharmacological stressor yohimbine produced increased levels of plasma NE metabolite, as well as PTSD symptoms such as intrusive memories of trauma, emotional numbing, and grief. 81 Geracioti et al. 82 reported that combat-related film footage also elicited elevations in NE levels in the cerebrospinal fluid of combat-related PTSD patients compared to neutral film footage. In contrast, cerebrospinal fluid concentrations of corticotropin-releasing hormone, also known as corticotropin-releasing factor, were shown to be reduced in response to the traumatic film compared to the neutral film. Yehuda et al.80,83–86 have reported numerous findings on the relationship between low cortisol levels and the risk for PTSD. Urinary cortisol samples revealed that combat veteran PTSD patients had low levels of cortisol excretion compared to non-PTSD individuals, and these low levels were associated with PTSD symptom presentation. 80 Subjects administered a low-dose dexamethasone test exhibited greater cortisol suppression compared to healthy participants, suggesting greater negative feedback sensitivity of the HPA axis and subsequently lower baseline cortisol levels in PTSD. 86 Competing findings on baseline levels of cortisol remain, with some studies describing increased levels in PTSD, whereas others report blunted baseline levels as mentioned above.78,84,87 These mixed findings suggest another measure may be necessary to understand the role of stress hormones in PTSD.
Stress hormones also influence brain structure and function as many preclinical studies have indicated. The brain regions involved in fear and PTSD circuitry, such as the amygdala, prefrontal cortex, cingulate cortex, and hippocampus, are significantly affected by stress.79,88–90 Evidence in the animal literature describes dendritic remodeling in regions that are necessary for fear processing following stress exposure. In rodents, neuronal hypertrophy within the amygdala and dendritic atrophy in the hippocampus were observed following chronic restraint stress or prolonged periods of glucocorticoid exposure.91–95 Chronic restraint stress exposure for 21 days also reduced dendritic branching within the mPFC in rats, 96 suggesting less synaptic plasticity and function within the mPFC and less control over the amygdala. Furthermore, in vivo single-unit recordings in rats that were chronically stressed revealed hyperexcitability in neurons within the lateral amygdala. 97 These preclinical findings may contribute to the understanding of the hyper-responsivity of the amygdala that is observed in PTSD patients in response to fearful stimuli.98,99
The overlap in stress and fear neurocircuitry facilitates the use of fear behavioral paradigms to uncover neural and psychophysiological signatures of PTSD, which present symptoms related to both systems. In fact, one might consider chronic stress part of the PTSD symptomatology, as it is a typical byproduct of the nightmares, flashbacks, and recurring intrusive thoughts or reminders of the trauma common with the disorder. Investigating the role of stress in PTSD has revealed that stress exposure can enhance fear learning and impair fear extinction, with associated structural and functional changes within the amygdala and other nodes of the fear network.100,101 In rodents, chronic injections of stress hormone corticosterone enhanced fear memory and increased memory-related activity within the amygdala. 102 Rats that were exposed to chronic restraint stress also exhibited improved fear acquisition, but poor extinction memory retrieval, which seemed to be associated with a stress-induced reduction in IL firing. 103 Three exposures to an uncontrollable stressor before fear conditioning not only induced dendritic retraction within the IL but they also impaired extinction. 104 The specificity of this effect of stress to impact only the IL, and not the PL, morphology is interesting to note given its critical role in extinction. Chronic stress also appeared to increase generalization of fear across contexts, disrupting fear extinction and increasing neuronal activity within the amygdala and hippocampus. 105 These preclinical findings are consistent with the impaired extinction recall and decreased vmPFC (and increased amygdala) activation observed in people with PTSD.5,6,57
There are relatively few human studies examining the effects of stress exposure on extinction learning and memory, and within these, there appear to be inconsistencies. Some of these inconsistencies may be attributed to the timing of exposure to stress and sex differences.69,106,107 Studies have reported an impairment in extinction memory retrieval as measured by SCR following stress exposure.108,109 However, some studies also report stress-induced impairments in fear memory retrieval, which may actually enhance extinction processes. 110 In one study using the cold pressor test, there was no effect of stress exposure on fear extinction learning; however, fear memory retrieval was found to be impaired in healthy men but not in women. 111 Aside from the potential sex differences, this result may be attributed in part to the timing of the stressor exposure; HPA activity and stress hormone levels vary based on the time of day and as a function of time since the onset of the stressor. 112 Thus, the distinction between acute and chronic stress effects necessary, as studies could yield opposing results on fear extinction learning and memory simply due to differences in this variable. Moreover, there is strong evidence that fear memory is enhanced when the timing of NE and cortisol release is synchronized but not when the timing is off. 69 The interaction between the NE and cortisol systems has been shown to modify brain activations within the fear network during encoding of neutral and emotional stimuli, with deactivation within prefrontal areas specifically noted. 113
The mixed findings of stress effects on fear extinction may also be due to sex differences and/or the potential influence of gonadal hormones in stress and fear extinction.114–119 Indeed, there are reported interactions between the stress and gonadal hormone systems that can influence fear behaviors.120–124 Antov and Stockhorst
114
examined the interaction between menstrual phase, stress, and fear in the effect of psychosocial stressor exposure on fear conditioning and extinction. No differences in the effects of stress on fear acquisition were observed, but women stressed in the high estrogen phase of their menstrual cycles exhibited enhanced extinction recall. Women stressed in their low estrogen phase, however, exhibited impaired extinction memory. Therefore, sex and gonadal hormones should be taken into consideration when using neural and/or psychophysiological fear responses as biomarkers of fear dysregulation in PTSD as indicated in Glover et al.
115
and Zeidan et al.
119
These and other factors potentially influencing the effects of stress on the brain and fear memory have been summarized in Figure 2.
The effects of stress and influencing factors on fear memory. Stress exposure (acute or chronic) induces stress responses that include changes in cortisol, norepinephrine, and epigenetic mechanisms. These responses to stress can affect fear memory by either increasing or decreasing it during retrieval, and the direction of the effect is dependent on various influencing factors such as individual differences (accounting for personal experiences, coping styles, etc.), sex (or gonadal hormones), stress hormone levels at the time of the stressor/learning/memory recall, brain responses to the stressor, type of stressor, and the presence of PTSD/anxiety symptoms and severity. Disrupting retrieval of the fear memory may enhance extinction processes, suggesting that stress (or stress hormone) administration may improve response to extinction-based treatments such as prolonged exposure therapy.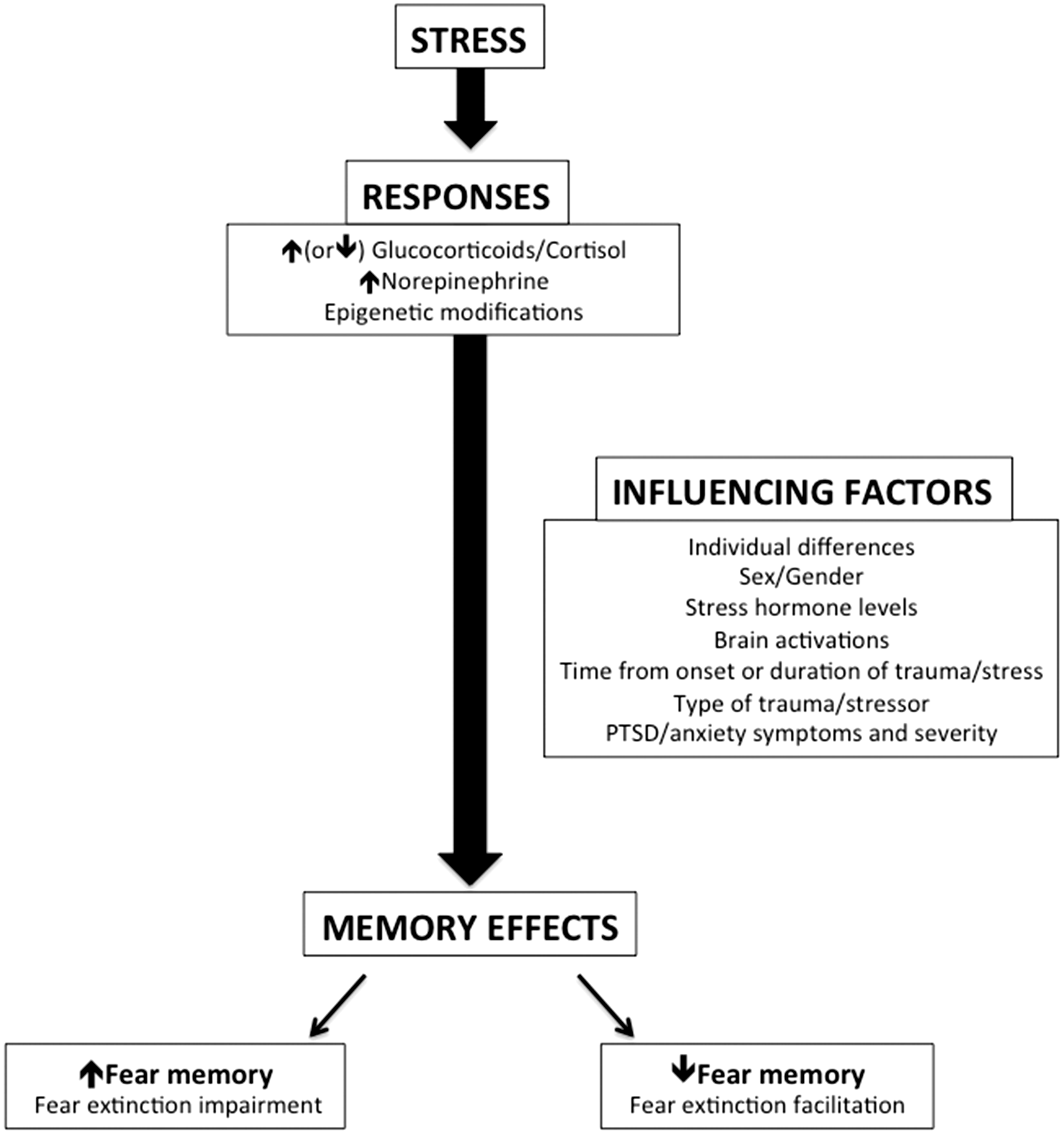
RDoC and PTSD
When diagnosing medical problems, physicians run through a checklist of symptoms, but conditions can still be misdiagnosed if the proper follow-up questions are not asked. For example, chest pain is a shared symptom of multiple medical problems, including angina, aortic dissection, pneumonia, gastroesophageal reflux disorder, and costochondritis, the inflammation of the cartilage joining the upper rib with the breastbone. If a patient arrives at a hospital complaining of this symptom, the efficacy of their treatment hinges upon the medical professional’s ability to properly identify its root cause. Luckily, we have a strong understanding of how the heart functions, and through physical exams and history taking, physiological symptoms and genetics are taken into account to arrive at a more specific and effective treatment. 125 Medical professionals treating mental illness follow a similar checklist of symptoms to help identify disorders. However, with the nature of the DSM-5 yielding 636,120 ways to have a PTSD diagnosis, as reported by Galatzer-Levy and Bryant, 126 and the diagnosis further dependent on subjective reports of symptoms, effective diagnosis (and subsequent treatment) can be a challenge. The RDoC framework may be a valuable tool in this regard, supplementing the DSM-5 and enabling us to obtain more comprehensive and quantifiable diagnostic biomarkers that span across mental disorders.
Diagnoses determined via the DSM alone can often lead to the exclusion of individuals who experience symptoms of PTSD but are subsyndromal or subthreshold for the criteria for PTSD nosology. These people may be suffering from symptoms of mental illness but will not receive proper treatment simply because they do not present their condition in a way that aligns with the DSM-5. On the other hand, it may also be possible for an individual to present biomarkers (i.e., an aberrant functional activation of the amygdala) for a PTSD diagnosis, despite not having the symptoms/disorder. Identifying associations between RDoC’s specific units of analysis and research domains (biological targets and functional constructs) will provide a more complete diagnosis/treatment profile to improve detection and treatment.3,127,128 Moreover, as more data are collected, these constructs and categorizations will become more refined, reliable, and transdiagnostic.
Moving away from the restrictive nature of the DSM-5’s distinct and somewhat limited classifications, we can provide more accurate and targeted treatments for individuals with various mental illnesses and comorbidities. 127 In fact, a recent study investigating the psychophysiological and neurobiological patterns during fear conditioning and extinction across anxiety disorders found no differences in SCR between the healthy and anxiety groups but did find differences within the vmPFC, rostral anterior cingulate cortex, and insula activations. 129 In line with the RDoC initiative, Marin et al. 129 also reported a negative correlation between trait anxiety levels and vmPFC activation within the anxiety group. This suggests a link between anxiety symptoms across different disorder diagnoses and a common neural correlate of fear.
In a recent assessment of the domains and constructs of the RDoC, fear conditioning paradigms were the recommended behavioral tasks to address the acute threat construct within the negative valence system domain. 130 A recommended behavioral task has not yet been defined for the sustained threat construct (i.e., chronic stress or distress construct) due to potential ethical violations associated with prolonged stress exposure. Few human studies have examined stress exposure effects on fear extinction learning and memory. However, utilization of a fear conditioning and extinction task in combination with measures of chronic stress using functional magnetic resonance imaging, psychometric, and psychophysiological methods can allow for multidimensional diagnoses. Findings from these studies provide valuable neural and behavioral data to aid in the development of a new classification system enabling improved diagnosis and treatment for individuals who share stress and anxiety-related symptomatology. 9
As chronic stress is a major contributor to the development and maintenance of symptoms related to PTSD and other psychiatric disorders such as anxiety, examining potential biomarkers of chronic stress (or “sustained threat”) may be critical to this paradigm. With the known biology of stress, we can measure stress hormones such as corticosterone in rodents and cortisol (i.e., chronic stress levels from hair samples) in humans; these measures have been extensively studied in PTSD research80,84,86,131 and may be valuable in improving diagnoses/treatment. Moreover, chronic stress not only affects cortisol and catecholamine release as reviewed above, but it can also influence cytokine signaling, alter levels of inflammation markers, and induce epigenetic modifications such as DNA methylation; these alterations have also been reported in PTSD.132,133 One study indicated that a specific glucocorticoid receptor gene polymorphism, the BcII GG genotype, was associated with lower levels of cortisol, more persistent traumatic memories, and increased PTSD symptoms, 134 providing additional evidence of stress hormone effects in PTSD at the genetic level. Subjective measures of chronic stress, including chronic stress scales and questionnaires such as the perceived stress questionnaire, can elucidate the mechanisms that underlie the psychological impacts of stress, subsequently contribute to mental illness, and affect the neurobiology.135–137 The relationships and correlations between these psychological and biological markers of stress and the psychological, physiological, and neural substrates of fear can further aid classification of the symptoms and profiles of individuals suffering from stress- and anxiety-related disorders.
Psychiatric disorders such as PTSD and anxiety disorders are often accompanied by disturbances in learning and memory processes and altered function within the brain regions modulating these processes. These brain regions are involved in fear circuitry and are affected by chronic stress as well.138,139 Fear conditioning and extinction protocols already contain a learning and memory component that allows for evaluating the cognitive effects of fear responding associated with PTSD and other psychiatric disorders. Further evaluating cognitive performance by combining these protocols with an additional and more general learning and/or memory task (i.e., working memory task) that has been extensively studied and has well-known neurocircuitry may provide more useful or comprehensive insight that is not limited to the fear network.45,140
PTSD Treatment
Current therapies for PTSD, such as exposure therapy, are based on extinction processes, and as a result, having a tool to evaluate how well an individual can learn safety signals (or learn not to fear once the threat is no longer present) is integral to the success of these treatments.50,141–143 For individuals with PTSD, trauma is continually re-experienced; living in a chronically stressed condition is the nature of the illness. Interestingly, it is commonly also the nature of their treatment in prolonged exposure therapy with repeated sessions of re-experiencing the trauma. If stress can modulate critical extinction processes, understanding how, when, and where this modulation occurs might offer insight into why treatments for PTSD work for some people and not for others. Among studies on the effect of stress and cortisol on fear extinction that report impairments, many findings also describe disrupted fear memory retrieval, which may enhance extinction memory. 144 More research is necessary to determine how these effects of stress and stress hormones can modify fear extinction and the relevant neurocircuitry. Interestingly, some PTSD studies have used the fear memory-impairing effects of stress hormones by administering NE or cortisol during PTSD treatment to facilitate extinction by impairing retrieval of the trauma memory.145–149 One study examining military veterans with PTSD, who received hydrocortisone or placebo, revealed a reduction in PTSD symptoms with hydrocortisone treatment. 149 This effect was particularly pronounced in those with glucocorticoid sensitivity, supporting the rationale for augmenting prolonged exposure therapy with glucocorticoids for combat-related PTSD. 149 As mentioned previously, sex and gonadal hormones can also affect extinction processes critical for treatment of PTSD and its symptoms and should therefore be considered as factors in classification and treatment. An example of this can be seen in cortisol treatment after reactivation, which demonstrates differential effects on reconsolidation in healthy men and women, enhancing the reactivated fear memory in men and having no effect in women.150,151 This is important as these differences in the effect of cortisol on fear could subsequently produce differential PTSD treatment responses between the sexes.
Fear conditioning and extinction protocols also facilitate the ability to track neural changes and physiological responses with treatment progress in parallel. 152 Helpman et al. 152 examined a cohort of subjects with PTSD and trauma-exposed individuals without PTSD who underwent a two-day fear conditioning and extinction protocol before and after a 10-week long prolonged exposure treatment. It was found that from pre- to post-treatment, brain activations differed within specific brain regions, i.e., rostral anterior cingulate cortex and subgenual anterior cingulate cortex; reduced subgenual anterior cingulate cortex and parahippocampal activations were associated with reductions in PTSD symptom severity.
Similar to these post-treatment brain changes and improvements in PTSD symptoms in humans, there is also evidence in rodents that demonstrates rescued function of stress-affected brain areas, such as the mPFC and hippocampus, following rest after chronic stress.153,154 Moreover, Fucich et al. 155 report therapeutic effects of extinction training in rats that were fear conditioned and then exposed to chronic unpredictable stress; extinction training improved performance on cognitive flexibility and coping behavior tasks, an improvement that was associated with molecular mechanisms that involved protein synthesis within the mPFC. These findings maybe translatable to humans as they indicate that allowing time for recovery may mitigate some of the negative effects of stress, such as exaggerated fear response in the brain. These changes may again be detected and assessed by examining fear learning and extinction during functional magnetic resonance imaging. These clinical and pre-clinical results support the idea that examining the processes that underlie extinction learning, and memory will enable us to understand how treatments can modify brain circuits and neurobiological mechanisms to improve mental health.
Conclusion
Identifying more specific diagnostic biomarkers of PTSD and other stress-related disorders is critical to the improvement of current diagnostic tools and treatment options. It is evident that there is no single biological marker for PTSD vulnerability, and there is a strong rationale for moving away from DSM diagnosis-centric research to focus more on biological underpinnings of the symptoms individuals present.78,128 Stress, whether in the acute or chronic sense, is a persistent catalyst for the development and exacerbation of not only PTSD symptoms but also depression and anxiety.107,156–159 As such, measurements of stress (i.e., subjective reports/ratings, stress hormone levels, genetics, epigenetics) should be included in the study of symptoms across fear-related disorders. Given that the inherent interactions between the fear and stress systems expand the ability to study both networks and their influences in PTSD symptomatology, the fear conditioning and extinction paradigms discussed here provide a transdiagnostic tool to examine both mechanisms in psychiatric disorders. This examination is not limited to PTSD research but rather intended to obtain measures of critical biological markers that correlate with stress- and anxiety-related symptoms for broad applications. In some studies, it has been useful to integrate DSM criteria and RDoC analyses (across several domains/constructs) to gain multidimensional perspectives in identifying biomarkers. Schmidt and Vermetten 160 assessed the value of RDoC in PTSD research and found it useful (i.e., identification of amygdalar hyperactivity across domains) but still lacking in interpretation of findings and linking cellular and molecular mechanisms with symptomatology. As reviewed here, incorporating measures of chronic stress at both the psychological and biological levels within fear conditioning and extinction paradigms may help improve diagnostic efficacy and push forward the development of increasingly targeted treatments for stress- and fear-based disorders.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIMH grants 1R01MH097880-01 and 1R01MH097964-01 to Mohammed R Milad.