
Abstract
The survival, growth, self-renewal, and differentiation of human pluripotent stem cells (hPSCs) are influenced by their microenvironment, or so-called “niche,” consisting of particular chemical and physical cues. Previous studies on mesenchymal stem cells and other stem cells have collectively uncovered the importance of physical cues and have begun to shed light on how stem cells sense and process such cues. In an attempt to support similar progress in mechanobiology of hPSCs, we review mechanosensory machinery, which plays an important role in cell–extracellular matrix interactions, cell-cell interactions, and subsequent intracellular responses. In addition, we review recent studies on the mechanobiology of hPSCs, in which engineered micromechanical environments were used to investigate effects of specific physical cues. Identifying key physical cues and understanding their mechanism will ultimately help in harnessing the full potential of hPSCs for clinical applications.
Introduction
For every cell type, its microenvironment, consisting of chemical cues and physical cues, is essential for regulating and maintaining cellular functions and fate.1–4 Although early studies suggest chemical cues, such as growth factors, cytokines, hormones, and extracellular matrix (ECM) ligands, are the major factors that regulate cellular activity,5–8 recent studies on a wide range of cell types collectively indicate the importance of physical cues, such as intercellular forces, matrix mechanical properties, and surface topography, to further influence cellular regulation.9–11 These factors are most evident particularly during embryogenesis and organogenesis, where chemical cues deliver a direct signal to stem cells and progenitors via paracrine, autocrine, or endocrine signaling for an intracellular command, and physical cues set the stage for morphogenesis and guide the process during development.12–14 Observation and identification of such factors and their spatiotemporal changes led to a number of effective protocols for maintenance and differentiation of stem cells in vitro.15,16 Human pluripotent stem cells (hPSCs), including embryonic stem cells (hESCs) 17 and induced pluripotent stem cells (hiPSCs),18,19 are no exception, although these artificially established cell lines do not exist naturally in an adult body.
Over the past 15 years, culture of hPSC has advanced significantly thanks to identification of essential growth factors and specific substrate coatings that allow for cellular growth, survival, and maintenance of the undifferentiated state.20–22 For instance, adherent substrate coatings, such as Matrigel, purified ECM proteins, and even fully synthetic polymers, in combination with a chemically defined growth medium, such as mTeSR, can now sustain long-term maintenance of hPSCs under a serum- and feeder-free condition.20,23–25 Such chemically defined culture systems further enabled discovery of a vast array of imperative regulations and signaling cues of stem cells. For example, specific growth factors in the transforming growth factor–β (TGF-b) superfamily, basic fibroblast growth factor, and insulin-like growth factors are essential for hESC maintenance, survival, and proliferation (reviewed extensively by Avery et al. 26 and Oshimori and Fuchs 27 ). In addition, control of stem cell fate by specific inhibition of certain kinases such as the glycogen synthase kinase (GSK), mitogen-activated protein kinase (MAPK), and Rho-associated protein kinase (ROCK) has been demonstrated recently.28,29 On the other hand, understanding physical cues influencing hPSC regulations has been left behind until very recently. In one notable example, when subjected to cyclic biaxial mechanical strain, the TGF-β/Activin/Nodal pathways were activated and effectively inhibited differentiation of hESCs even without cytokines TGF-β1 and Activin A, suggesting that mechanical cues are capable of regulating certain pathways.30,31 Clearly, detailed knowledge of the chemical and physical factors, as well as their synergistic effects, is necessary to emulate the complexity of the in vivo niche that controls cell transformation during the embryological process.32,33
Recent advances in micro- and nano-fabrication technology and materials science has made engineered tools and platforms available for investigating effects of micromechanical cues on stem cell regulations. For example, specialized biomaterials have been used in hPSC cultures to mimic ECM properties of live tissue models.34–36 Furthermore, effects of physical cues such as matrix stiffness, surface texture, and surface topography on hPSCs and adult stem cells have been explored.37–41 These studies have successfully observed how stem cells respond to these cues and started shedding light on pathways that dictate their fate for survival, self-renewal, and lineage commitment.33,42 It is now evident that stem cells are capable of detecting various physical aspects of their microenvironment and translate such cues into intracellular biochemical signals that modulate cellular activity.43,44 Nevertheless, the complete mechanisms governing so-called mechanotransduction of hPSCs by physical cues are far from being understood. In this review, to continue this progress, we will first summarize how stem cells in general can sense different physical cues in their environment by referring to previous mechanobiology studies. Interpreting results obtained from different cell types, particularly human mesenchymal stem cells (hMSCs), can segue into understanding hPSC regulations in engineered micromechanical environments. We will then survey recent studies that have begun to show the effects of engineered microenvironments in determining the fate of hPSCs for survival, self-renewal, or differentiation. Finally, we will conclude by offering what we envision in the future progress of this emerging field.
Mechanosensory Machinery and Force Transmitter
The distinction between chemical cues and physical cues is how stem cells detect them and process the information subsequently.44,45 Extrinsic chemical cues are detected by binding to and activating specific membrane protein receptors, which then transmit internal signals to activate (or inactivate) protein activators and co-activators for gene regulation. On the other hand, specific mechanisms by which stem cells sense physical cues, particularly mechanical stimuli, are not fully understood. However, accumulated evidence suggests that the peripheral transmembrane proteins, such as integrins and cadherins, and nonmuscle myosin II (NMMII) play an active role in detecting and responding to the physical changes of the microenvironment. 46 Integrins and cadherins function as the mechanosensory machinery, and NMMII is a complex motor protein that binds to cytosol’s actin filaments to form the actomyosin cytoskeleton ( Fig. 1 ). As a result, cellular activity for motility, morphogenesis, and gene transcription could be regulated in the process of adapting to their physical micromechanical environment. An excellent review of mechanotransduction machinery and transcriptional regulators of both stem cells and non–stem cells was recently published by Mammoto et al. 47 In our current article, because of limited space, we focus on the major players including integrins, cadherins, and NMMII.
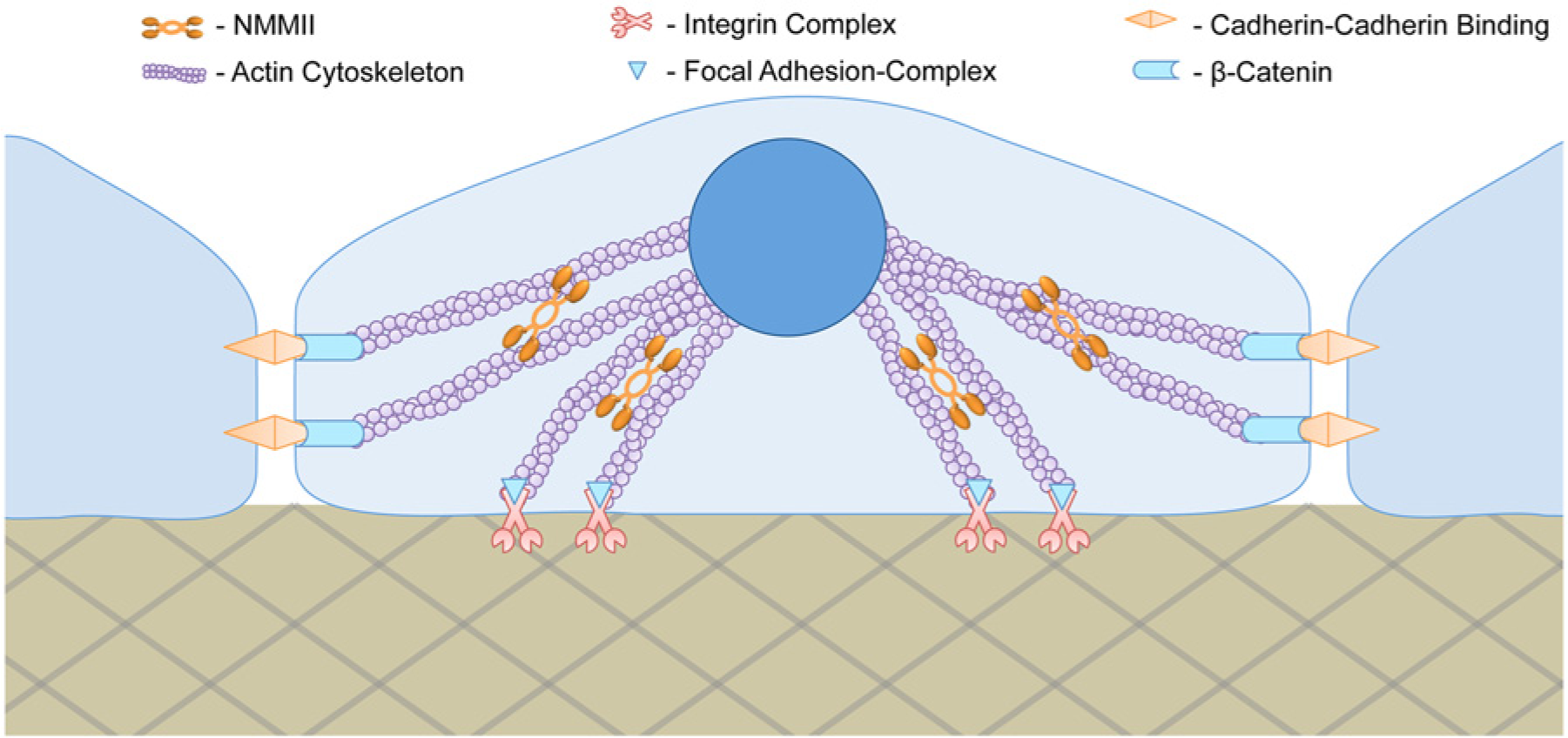
Cell mechanical regulations simplified. Cellular adhesion to extracellular matrix and adjacent cells are regulated by integrins at the focal adhesion and by cadherins at the adherens junction, respectively. Inside the cell, both integrin and cadherin are integrated with the actin cytoskeleton mediated by other proteins such as focal adhesion complex for integrins and β-catenin for cadherins. The actin cytoskeleton supports the cell structure and shape throughout the cytoplasm. When intracellular activity signals to alter cell polarity, nonmuscle myosin II (NMMII) mediates reorganization of actin cytoskeletal tension and induces changes in cell morphology.
Integrin: Cell-ECM Mechanosensor
For every adherent cell, anchorage to the ECM is made by cell membrane–bound integrin-complex proteins at the focal adhesion site. 48 On the internal side of the membrane, the integrin proteins are also integrated with the actomyosin cytoskeleton ( Fig. 1 ). 49 Therefore, the link between cell membrane and the ECM goes far beyond the attachment site and is actually the start point of a communication network between the cell and microenvironment.50–52 Cell-ECM communication is vital for hPSC function and activity as it provides essential signals for maintenance of pluripotency, self-renewal, and survival. 53 However, little is known about mechanical cues induced by the ECM that control stem cell regulation and fate.54,55 One certain thing is that the properties of the ECM and underlying substrates such as elasticity, porosity, and spatiotemporal geometry and topography are among the physical stimuli that a cell, in essence, can “feel.”56,57 When the integrin complex binds to the ECM, the cell tends to conform to its microenvironment by modulating its intracellular mechanics. For instance, when murine or human MSCs were attached to a rigid substrate, they generated more traction force and consequently increased their cytoskeletal tension, emulating the substrate’s rigidity. 38 Conversely, when murine ESCs were attached to a substrate whose stiffness was similar to their normal cytoplasmic value, the cells sustained survival, self-renewal, and the undifferentiated state.58,59 It is speculated that integrin adjustment and clustering is responsible for the increase in traction force and subsequent cytoskeletal tension.60–63 Nevertheless, many studies point to NMMII activity and the Rho/ROCK pathway as the major players that generate cytoskeletal tension. The Rho/ROCK pathway is a molecular feedback mechanism known to be associated with the mechanosensory machinery that regulates NMMII activity (see “NMMII: Mechanical Motor Modulating Cellular Function and Cytoskeleton”). Importantly, formation of focal adhesion up-regulates Rho/ROCK and consequently induces NMMII activity to increase traction force as well as cytoskeletal tension.
Integrin-mediated adhesion pathways are vital for stem cell function and activity. For instance, researchers observed that an elevated activity of the Rho/ROCK pathway was consistent with hyperactivation of NMMII, followed by an instability and destruction of the cytoskeletal structure.64,65 This is now thought to be the main cause of dissociation-induced apoptosis of hESCs. As such, inhibition of the Rho/ROCK pathway results in a higher percentage of hESC survival upon enzymatic dissociation into single cells. 66 Similarly, the phosphoinositide 3-kinase/Akt (PI3K/Akt) pathway and MAPK/extracellular-signal regulated kinase (MAPK/ERK) pathway are the other integrin-mediated adhesion pathways critical for cell survival and maintenance of pluripotency.67–71 Particularly, studies suggest that Akt negates apoptotic signals for sustaining continuous survival or delays the onset of cell death when single cell dissociation occurs.72,73
However, identifying an exact molecular mechanism through which extrinsic forces command stem cell regulations has proven to be difficult due to cross-talk among many defined and undefined factors in the microenvironment. In an attempt to decouple mechanical factors, Huebsch et al. 74 engineered a substrate with defined mechanical properties and studied their effects on MSCs’ lineage commitment. By using a three-dimensional (3D) hydrogel-based, synthetic ECM conjugated with integrin-binding RGD (Arg-Gly-Asp)–modified ligands, they observed that differentiation correlated with traction-dependent reorganization of integrins and the adhesive ligands in response to matrix elasticity and 3D architecture. 74 In addition, some have suggested that mechanical cues may cause tension-induced conformational changes of membrane proteins to expose sites that favor ligand interactions. For instance, Friedland et al. 75 showed that integrin-complex receptors can alter their affinity strength in response to cytoskeleton tension and stress. The same physical force may also present dormant growth factors in an active form when stress is involved. 76 In addition, there are numerous examples of ion channels, membrane receptor proteins, and intracellular proteins being constantly regulated when mechanical forces are applied.77–80 It is, however, still debatable how exactly mechanical stimuli influence changes in transcriptional regulation. Nevertheless, integrin-ECM binding provides crucial inputs for chemical and physical signal transduction. 81
Cadherin: Cell-Cell Mechanosensor
Similar to the integrins complex on the focal adhesion site, transmembrane glycoprotein cadherins in the adherens junction have also been shown to function as a sensor for many cellular activities. 82 Besides mediating cell-cell interactions via a cadherin-cadherin bond as a form of communication among neighboring cells, the adherens junction is capable of detecting external forces in its peripheral surroundings, independent of the ECM. 83 Figuratively, the integrin complex acts as a foot to feel its platform, whereas the cadherin acts as a hand to sense its lateral microenvironment ( Fig. 1 ). This mechanism is essential for hESCs because organogenesis depends on cell-cell communication during the embryological process. 84 For instance, E-cadherin is highly expressed in undifferentiated hESCs, whereas an increase in expression of N-cadherin and VE-cadherin is required for neural formation and angiogenesis, respectively.85,86
The adherens junction is also linked to the cytoskeletal architecture of the cell via cadherin binding to β-catenin in the cytoplasm ( Fig. 1 ). Therefore, when cadherin-cadherin binding forms between two cells, the cytoskeletal backbone is also connected together. Consequently, each cell can affect cytoskeletal tension of the other in the presence or absence of an external force. For example, Liu et al. 87 observed that the adherens junctions between two endothelial cells changed in size and strength in response to cell-cell contact force under mechanical loading. Using a micromechanical force sensor, they measured traction force generated by the cells and quantified local contact force at the cell-cell adhesion site. This entails that the cadherin feedback mechanism is similar to that of integrins, where force-induced cellular functions are mediated by myosin activity. 82 It is important to note that even though Liu’s work was based on endothelial cells, integrin-based adhesion and cadherin-based adhesion are vital anchorage mechanisms that link to the actomyosin cytoskeleton in hPSCs. Consequently, both adhesion complexes may have the ability to mediate force-dependent regulation for survival, growth, morphology, and function of stem cells. Recent studies demonstrated that an excessive perturbation of cell-cell contact in hESCs resulted in collapse of the intracellular architecture, leading to cell blebbing and eventually dissociation-induced apoptosis.64,65 This process was likely mediated by the Rho/ROCK pathway and NMMII hyperactivation.
Even though both the focal adhesion and adherens junction function as mechanosensory machinery, it is said their signaling mechanism can have different effects on hPSC fate and activity. 71 For instance, some reports suggest that the focal adhesion mechanism is necessary for cell migration and differentiation of hPSCs, 88 whereas cadherin binding is necessary for colony formation, pluripotency maintenance, and long-term survival. 89 The cadherin-mediated regulation was evidenced in a study by Li et al. 89 on short-term and long-term inhibition of NMMII using blebbistatin, a NMMII ATPase inhibitor. Temporal inhibition of NMMII using blebbistatin increased single cell survival of hESCs after enzymatic dissociation; however, prolonging inhibition attenuated cell-cell contact and gradually increased apoptosis.89,90 Importantly, they demonstrated that an overexpression of E-cadherin in NMMII-depleted hESCs was able to recover cell-cell contact and overall colony formation as well as to reverse the down-regulation of pluripotency markers. 89 This suggests that cell-cell contact and colony formation are beneficial to long-term survival and maintenance of the pluripotent state of hESCs.
NMMII: Mechanical Motor Modulating Cellular Function and Cytoskeleton
NMMII is known as the critical mechanical motor of a cell because of its integral role in remodeling the cytoskeletal architecture for cellular adhesion, motility, morphology, and overall modulation of polarity. 46 The actomyosin cytoskeleton is intricately connected throughout the cytoplasm, from the membrane to the nucleus ( Fig. 1 ). 91 Consequently, when physical cues from the ECM or adjacent cells are sensed by the mechanosensory machinery, pathways such as Rho/ROCK are up-regulated to activate NMMII, which then helps reorganize the cytoskeleton architecture ( Fig. 2 ).92,93 It is hypothesized that altering the cytoskeleton architecture affects organelle localization and nuclear conformation, thus mediating transcription regulations vital for the fate of stem cells.47,94
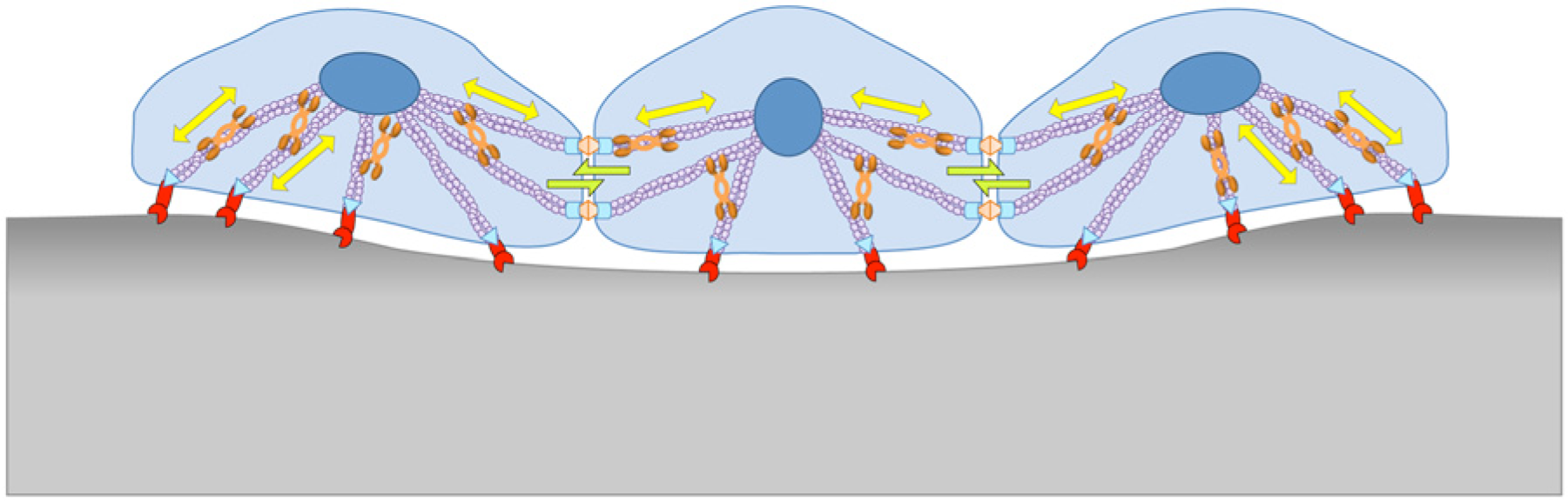
Mechanics of cell–extracellular matrix (ECM) and cell-cell interactions. Mediated by nonmuscle myosin II (NMMII), forces exerted by the ECM and neighboring cells can affect contraction or elongation. In turn, local deformation of the cell and the ECM can be dictated by their mechanical properties and number of cell-ECM and cell-cell force interactions.
Earlier studies that evidenced critical involvement of NMMII and pioneered the field of stem cell mechanobiology were conducted using hMSCs. McBeath et al. 37 examined how directed cell shapes contributed to hMSCs’ commitment to osteoblast or adipocyte. Using a surface micropatterning technique to control cell shape and the degree of cytoskeletal tension, the group observed that cytoskeletal tension was the driving force of lineage commitment. 37 Furthermore, cell shape and tension were highly regulated by Rho/ROCK activity on the actomyosin cytoskeleton; in fact, it was the NMMII-mediated spreading and contracting mechanism that controlled differentiation pathways regardless of soluble differentiation factors. 39 Engler et al. 38 also showed that NMMII activity directed naive hMSC lineage commitment, but in this case, it was the matrix elasticity that dictated cell morphology by modulating NMMII activity and consequently cytoskeletal structure. This cytoskeletal remodeling coincided with dramatic change in transcriptional regulation, resulting in lineage commitment toward a cell type whose elastic modulus was similar to that of the underlying substrate. 38
Micromechanical Control of hPSC Fate
Given the multifaceted factors influencing stem cell regulations, numerous strategies have been explored to efficiently control cellular function for proliferation and differentiation.95–98 Certainly, the lack of efficiency has always been a major issue for stem cell culture. Accordingly, researchers have been and are still developing methods to efficiently expand and/or differentiate stem cells with high purity of a desired phenotype.99–104 At an early stage, hESC culture relied on feeder cells and serum-supplemented media for continuous self-renewal. Since then, many chemically defined culture systems have been developed, eliminating exogenous undefined factors and animal products that are potentially problematic for clinical applications.98,105–108 For instance, Saha et al. 108 engineered ultraviolet/ozone-modified polystyrene surface patterns, which in combination with human serum or recombinant vitronectin coating improved long-term growth of hPSCs. The group, by controlling colony size with the developed technology, was able to achieve at least a threefold increase in cell number compared with a conventional culture employing feeder cells. 108 Although the physically controlled colony size likely modulated exposure to soluble factors and cell-cell interactions, detailed mechanisms explaining the observed effects were not fully understood.
Unlike murine ESCs, which reportedly favor a soft substrate whose stiffness is close to the intrinsic cytoplasmic value (~0.6 kPa) for sustaining self-renewal,58,59 hPSCs seem to favor stiff ones. Musah et al. 109 sought to determine an optimal mechanical property of engineered substrates for supporting pluripotency and robust proliferation of hESCs. By using a stiff polyacrylamide hydrogel (10 kPa) conjugated with glycosaminoglycans (GAG)-binding peptides, the group saw more robust attachment, growth, and maintenance of pluripotency compared with hydrogels of identical stiffness (10 kPa) conjugated with integrin-binding RGD peptides or a softer hydrogel substrate (0.7 kPa) conjugated with the GAG-binding peptides. In addition, they speculated that activation of YAP/TAZ in the nucleus was essential for pluripotency; as such, they saw a higher YAP/TAZ activation on the stiffer hydrogel than on the softer hydrogel. 109 Interestingly, when plated on hydrogels with an intermediate stiffness (3 kPa), the cells attached and formed colonies in a similar manner as on the stiffest hydrogel (10 kPa) but began to detach from the substrate in the middle region of the colony in subsequent days. This suggests that although the intermediate stiffness can facilitate cell anchoring and colony formation, it is not supportive of cell spreading and continuous self-renewal. 109 Lee et al. 110 also demonstrated that substrate properties could significantly affect hESC culture through a survey of multiple permeable membranes whose surface hardness was dictated by their material and pore density. They reported that a polyethylene terephthalate membrane with pore density of 4 × 106 pores/cm2 and hardness of 0.3 GPa was able to support cell attachment and growth of undifferentiated hESCs. 110 However, because the mechanical property of the membrane was coupled with the pore density and thus its permeability, one cannot conclude that hardness alone was the cause of the observed hESCs’ response. In an attempt to decouple mechanical properties of the substrate, Sun et al. 111 employed a polydimethylsiloxane micropost array, which allowed for tuning effective Young’s moduli by changing the post height without altering contact surface area, matrix porosity, or surface chemistry. In this study, they showed that rigid substrates (>15 kPa) promoted growth, colony formation, and maintenance of undifferentiated hESCs ( Fig. 3A ). In a different study, the same group explored the ability of hESCs to feel nanotopography by culturing the cells on a smooth surface or a rough surface. 112 Results from their nanotopography tests showed that hESCs altered the patterns of their focal adhesion formation in response to the underlying surface texture. Specifically, hESCs cultured on a smooth surface resulted in a focal adhesion formation on the periphery of the cell, whereas a random distribution of focal adhesion was observed for hESCs cultured on the rough surface ( Fig. 3B ). Moreover, hESCs cultured on a smooth surface were able to spread, form colonies, and self-renew, whereas hESCs cultured on rough surface exhibited a decrease in adhesion rate, colony size, and pluripotency. 112 These results collectively show that defined physical properties of the ECM and substrates can cause a cellular response, which critically affects hPSC growth, undifferentiation, and colony formation. In addition, these processes are mediated by the mechanosensory machinery and NMMII activity in hPSC, as is the case in hMSC. Similarly, differentiation of hPSCs could be influenced by the substrate’s mechanical properties. Recently, Keung et al.113,114 demonstrated that a soft polyacrylamide hydrogel substrate up-regulated early neural marker SOX1 in the absence of extrinsic neural inducers. However, the effect was limited to the first stage of differentiation, and further downstream differentiation required soluble neural factors. Interestingly, they observed that at different substrate stiffness, the cells’ proliferation rate and pluripotency did not vary during the first 3 days of culture, although colonies were significantly smaller and more compact on the softer ECM (0.1 kPa) than on the more rigid one (75 kPa). Nevertheless, the soft microenvironment promoted neural differentiation better than the rigid substrate, suggesting that the compact colony formation had a higher impact for differentiation than spread colonies did as seen on the rigid substrate.114,115
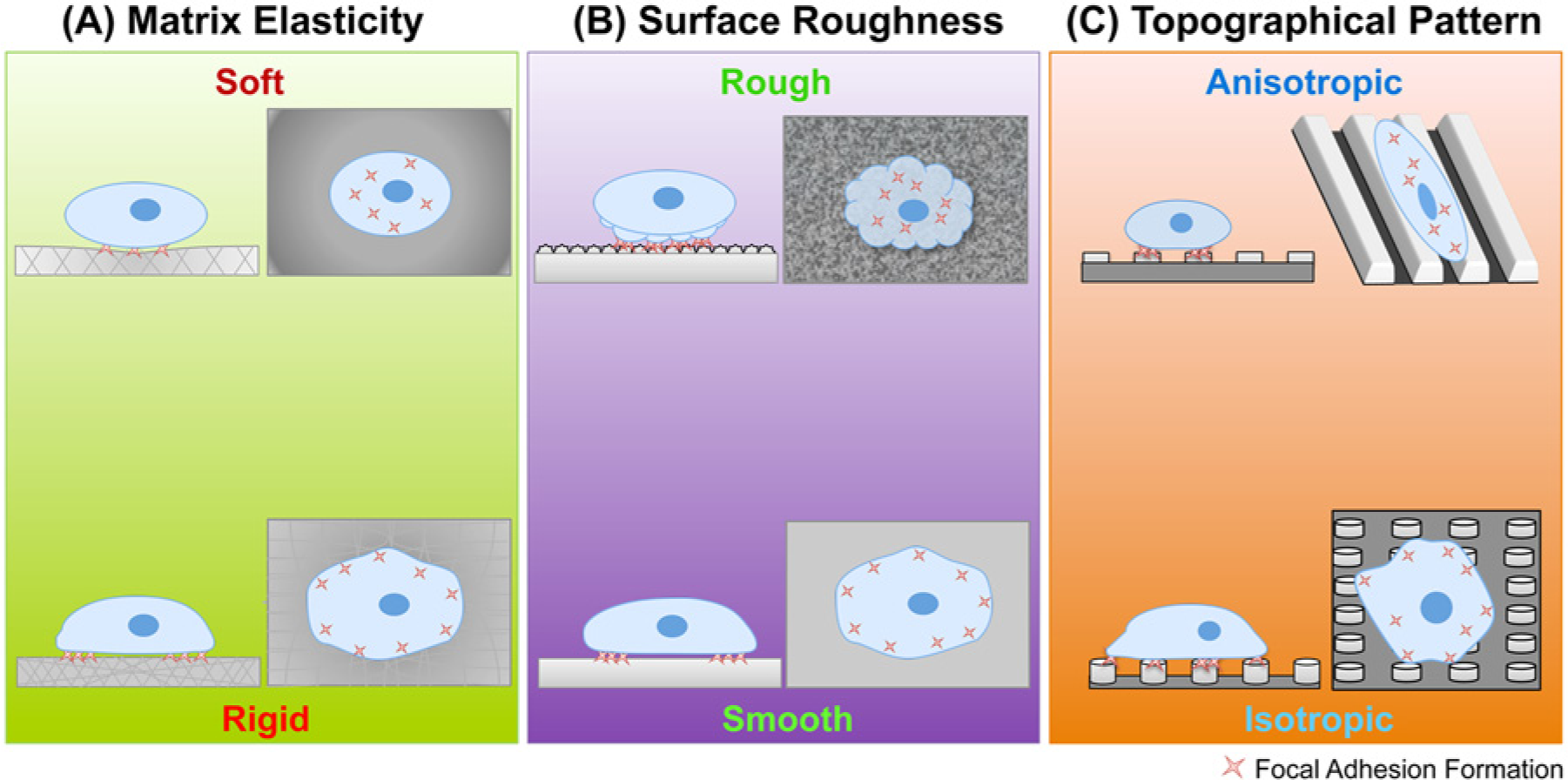
Substrate mechanical properties affecting behavior of human pluripotent stem cells. (
During embryogenesis, mechanical forces are particularly important for lineage specification to the primary germ layer: mesoderm, endoderm, and ectoderm.14,32 Zoldan et al. 116 studied how mechanical properties of a variety of 3D porous polymer scaffolds in vitro can guide hESCs into either one of the germ layer. The group reported hESCs remained undifferentiated in the scaffold with the highest elasticity (>6 MPa), suggesting it supports maintenance of pluripotency. In addition, the group observed that hESCs cultured on substrates with high (1.5–6 MPa), intermediate (0.1–1 MPa), and low (<0.1 MPa) elasticity expressed gene markers specific for mesodermal, endodermal, and ectodermal differentiation, respectively. They speculated that the scaffold elasticity affected motility, traction force, and cytoskeletal tension and that the modulation of such cellular conditions could result in changes in morphology, adhesion to the ECM, cell-cell contact, and transcription relevant to differentiation, thus allowing hESCs to adjust to the microenvironment. 116
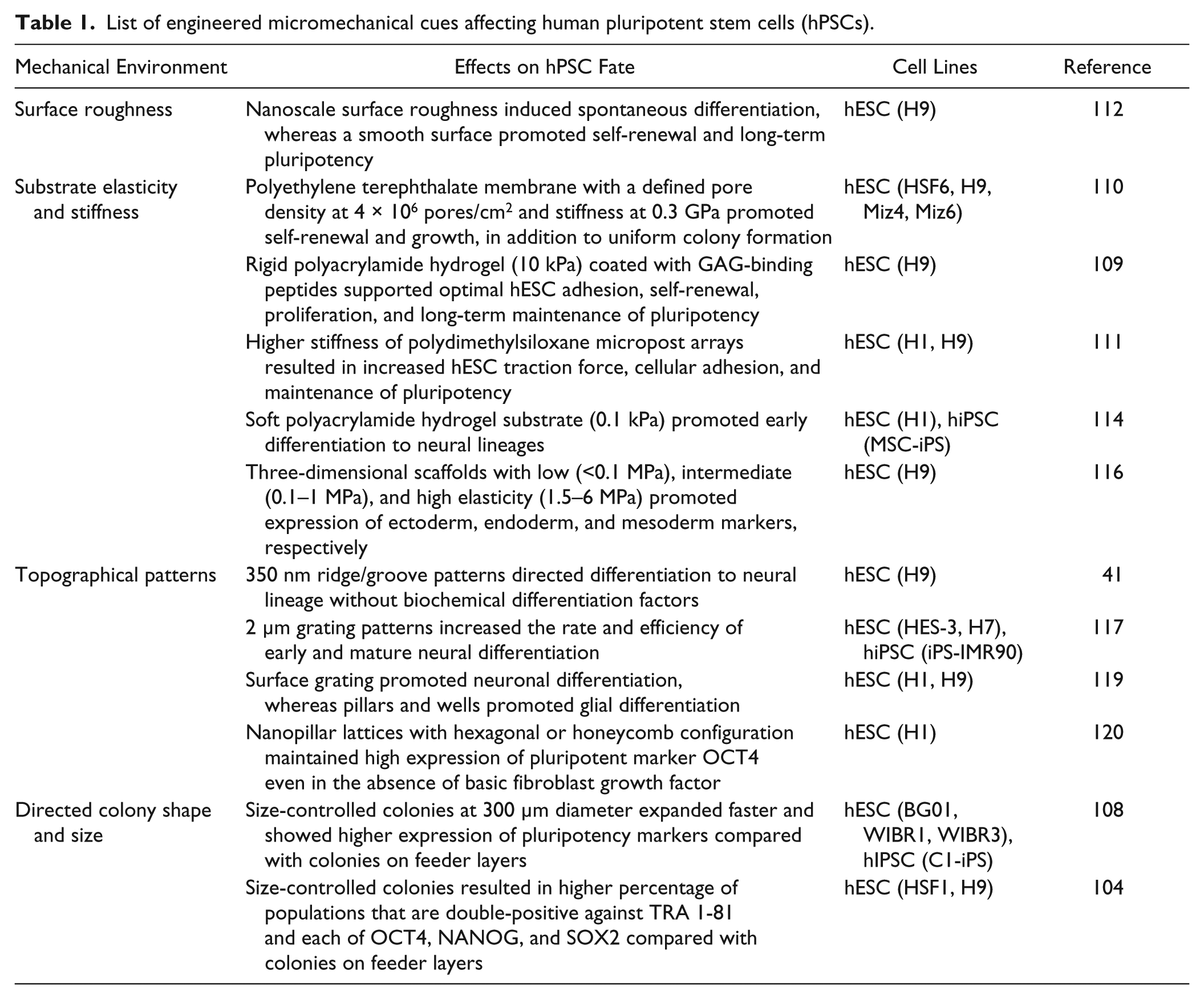
Other engineered micromechanical environments have been employed to explore optimal mechanical conditions to direct hPSC fate. For instance, Chan et al. 117 demonstrated how a specific topography can influence neural differentiation of hPSCs, even in the absence of chemical differentiation factors. They cultured hESCs on a 2 µm pitch, linear grating and observed cell and colony morphology being elongated along the grating direction, mimicking a neural lineage state even in a chemical condition promoting the undifferentiated state ( Fig. 3C ). When cultured in differentiation-inducing media, hPSCs quickly committed to a neural lineage, showing the collaborative effect of both physical and chemical cues. 117 The synergistic role of chemical cues and physical cues presents great potential for leveraging control of hPSCs by the engineered microenvironment; however, identifying optimal cross-talk brings the need for screening multiple physical cues that support differentiation into many cell types.41,118 For this reason, Ankam et al. 119 developed a multiarchitectural chip (MARC) to screen multiple combinations of topography designs specifically for ectoderm differentiation. The multitude of topography designs in the MARC varies in geometry such as grating, pillars, and wells, as well as their 3D dimensions. Overall, they observed that neuronal fate was highly favored on anisotropic geometries, such as linear gratings, where cell morphology was aligned and elongated along the grating direction, inducing neuronal differentiation. In contrast, they observed that glial fate was promoted on isotropic geometries such as pillars and wells, where the cells stayed round and spread along all directions of the patterned substrate ( Fig. 3C ). 119 The spreading morphology of hESCs on pillars was also observed by Kong et al. 120 ; however, in this study, a higher percentage of OCT4 (a marker of undifferentiated state) was observed in cells cultured under hexagonal and honeycomb pillar lattice configuration. It was suggested that the pillar patterns attenuated focal adhesion formation to shift dependency on cell-cell contact for survival and self-renewal. 120 Altogether, physical cues such as substrate stiffness and topography have a profound impact on hPSC regulation, which in turn can dictate its survival, self-renewal, or differentiation. Recent demonstrations of hPSC control using micromechanical environments are summarized in Table 1 .
List of engineered micromechanical cues affecting human pluripotent stem cells (hPSCs).
Summary and Future Perspective
With help from recent progress in micro- and nanotools and technologies, researchers are beginning to understand the mechanobiology of hPSCs and specific physical cues associated with cellular regulations. For instance, similar to hMSCs, it is now evident that hPSCs are equipped with molecular machinery for sensing various physical cues in their surroundings. These mechanosensors are essential interpreters of cell-ECM and cell-cell physical interactions and translate physical cues into intracellular biochemical signals. Subsequently, the biochemical signals can direct many cellular activities such as regulating cell polarity for cell migration, contraction, or expansion. Changes in cell polarity are regulated by the NMMII activity. In addition, cytoskeletal tension produced by hPSCs can dictate regulations for survival, self-renewal, pluripotency, and lineage specification. For this reason, some have tried to characterize the intracellular mechanics of hPSCs and their differentiated derivatives to uncover mechanical phenotypes indicative of hPSC lineage specification.121–123 Nevertheless, investigation of hPSC mechanobiology is still at an early stage, and much remains to be uncovered. For example, the majority of work done so far has only probed the effects of physical cues on either maintenance of pluripotency or initial commitment to differentiation lineages but not on long-term, terminal differentiation into fully mature cells. As the core value of hPSCs is their ability to differentiate into many, if not all, types of human tissues, understanding roles of physical cues in downstream differentiation will be particularly important for basic science as well as for clinical applications in which such knowledge is required to fully exploit differentiation potential of the cells.
To further investigate the impact of individual physical cues, new tools that allow for decoupling of physical properties and high-throughput screening methods will be desired. Innovative platforms such as the MARC 119 and TopoChip 124 have shown great promise by enabling high-throughput screening of defined topography on a single platform. However, as the density of test beds on chip increases, precise fluid handling (e.g., media exchange without chemical cross-talk among neighboring test sites) and analytical measurement technique beyond simple immunostaining and fluorescence measurement on chip needs to be developed. In particular, passaging cells in a multistep differentiation process, in which chemical stimulants are sequentially changed according to specific stages down the lineage commitment, will be extremely challenging in a high-throughput array format. Furthermore, upon successfully identifying potent physical factors that affect hPSC fate, screening of combinatorial cues of both physical and chemical factors should take place. Given that the in vivo stem cell niche comprises dynamic signaling of chemical and physical cues, an optimal microenvironment in vitro would most likely require a combination of multiple factors to unleash the full potential of the stem cell by maximizing synergistic effects. However, the factorial of multiple chemical and physical parameters suffers from an exponentially large combinatorial problem. To make matters worse, factoring timing in as one of parameters will inevitably make the challenge almost impossible. Therefore, new innovations, perhaps as a combination of nanofabrication, fluid handling, molecular imaging, automation, and optimization algorithm, will be necessary to complete such an extremely daunting task.
In addition, from the point of view of hPSC-based cell therapy, culture methods need to be scaled up to supply a clinically relevant number of transplantable cells. Three-dimensional cultures in which hPSCs are cultured within a scaffold or suspended as clusters with or without microcarriers in a culture medium are the likely solutions to meet such demands at reasonable cost. However, in such a platform, implementing physical cues such as defined surface topography or matrix stiffness would be difficult and require development of a whole new set of engineered tools and technologies to harness physical cues to control of stem cell fate.
All of these requirements highlight the fact that mechanobiology of hPSCs is a truly multidisciplinary field and offers a unique opportunity for exciting collaborations among stem cell biologists, chemists, materials scientists, physicists, engineers, and more.
Footnotes
Acknowledgements
The authors wish to thank Mr. Brent Kalish for help with proofreading the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by an initial complement fund from Bourns College of Engineering and a Regents’ Faculty Fellowship from University of California, Riverside.