
Abstract
Poor prognosis is associated with melanoma due to immunosuppression profiles, suggesting that immune alterations have an important role in the occurrence, growth, and metastasis of melanoma. Here, we found that PCC0208018, a small-molecule compound, enhanced T cell proliferation and activation to release interferon gamma (IFN-γ) and interleukin-2 (IL-2) without blocking the programmed cell death 1 (PD-1)/programmed cell death-ligand 1 (PD-L1) binding and did not directly affect tumor cell viability in vitro. Furthermore, PCC0208018 increased the phosphorylation of protein kinase B (PKB/AKT) as well as extracellular regulated protein kinases (ERK) in human peripheral blood mononuclear cells (PBMCs) in vitro. The secretion of cytokines induced by PCC0208018 was significantly suppressed by the PI3K inhibitor GDC-0941. In B16-F10 melanoma-harboring mice, PCC0208018 significantly inhibited tumor growth as well as increasing CD3+, CD3+CD4+, and CD3+CD8+ T cell abundance in tumors without affecting PD-L1 expression. This study showed that PCC0208018 potentially increased PBMCs proliferation and function by activating the phosphatidylinositol 3 kinase (PI3K)/AKT and mitogen-activated protein kinase (MEK)/ERK pathways to exert antitumor effects.
Introduction
In recent years, the number of melanoma cases has significantly increased and are often highly radio- and chemotherapy-resistant. High immunogenicity has been observed in cancer patients, and thus, cancer immunotherapy is the standard regimen for the treatment of melanoma. 1
T lymphocytes have a major role in cell-mediated immunity as well as cancer immunotherapy. Numerous immune checkpoint inhibitors targeting regulatory pathways can increase the antitumor responses of T cells in multiple tumor types, including advanced melanoma. Monoclonal antibodies that target programmed cell death 1 (PD-1)/programmed cell death-ligand 1 (PD-L1) axis, such as pembrolizumab and nivolumab, have been approved by the Food and Drug Administration (FDA) for the treatment of melanoma. However, compared with the disadvantages of monoclonal antibody therapies, small molecules show good affinity, high specificity, and good oral bioavailability without immunogenicity and immunotoxicity. Chemical inhibitors targeting the PD-1/PD-L1 pathway or PD-1/PD-L1 interactors, such as macrocyclic peptides, peptides, peptidomimetics, and small molecules, have been reported. 2
PCC0208018 is a 1,2,4-oxadiazole derivative (Figure 1(a)). It is considered the number one compound in the patent of Aurigene Discovery Technology, Ltd. (WO2015033299A1) and targets PD-1/PD-L1 to rescue mouse splenocyte proliferation in vitro. 3 However, its efficacy and mechanism are still unclear. As a result, we investigated the potential impact and mechanism of PCC0208018 on cytokine secretion in human peripheral blood mononuclear cells (PBMCs) in vitro. We also studied the antitumor and immunomodulatory activity of PCC0208018 in B16-F10 melanoma-harboring mice.
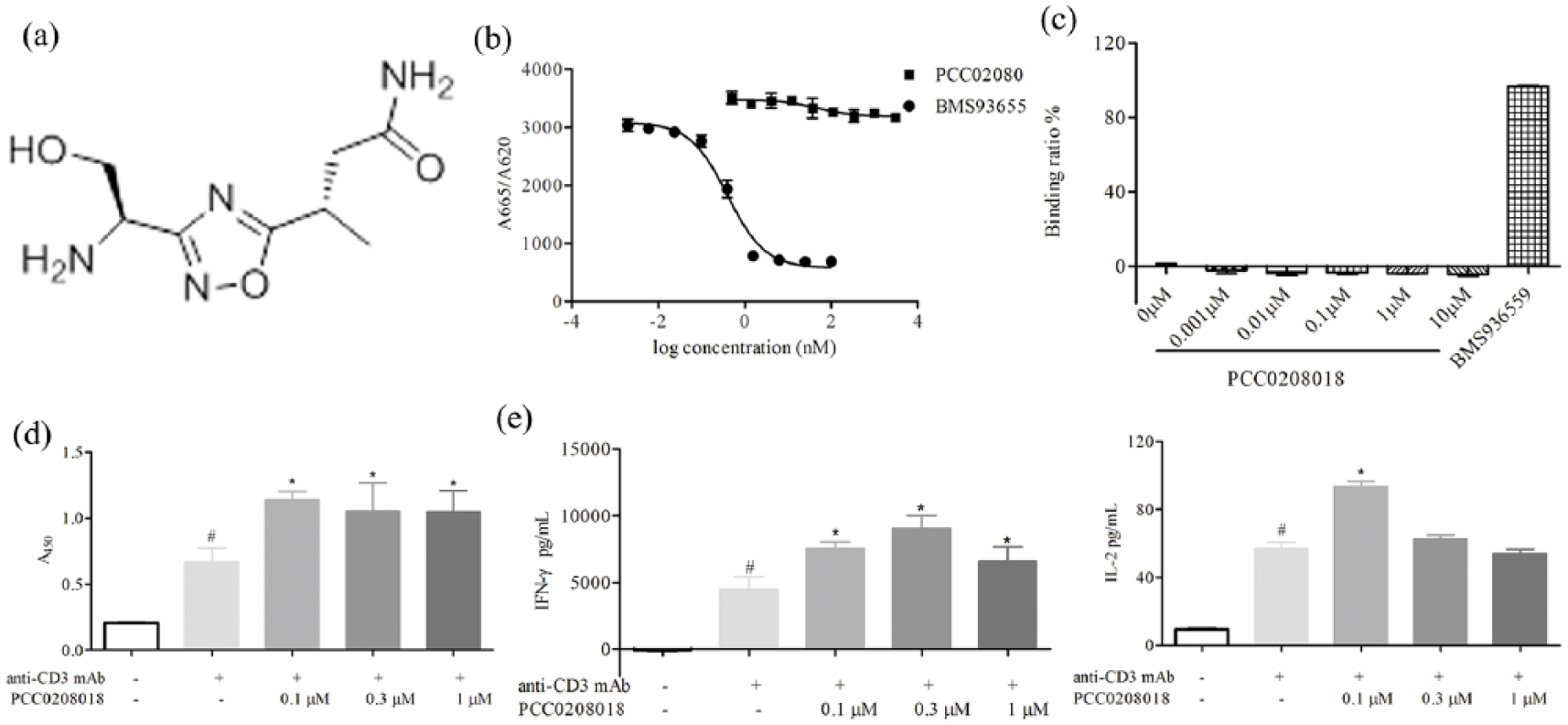
PCC0208018 increased PBMCs proliferation and cytokine secretion without interfering with PD-1/PD-L1 binding: (a) chemical structure of PCC0208018, (b) HTRF-based ligand inhibition assay of binding effects at the protein level, (c) binding assay of binding effects at the cellular level, (d) CCK-8 analysis of the effects of PCC0208018 on PBMCs proliferation, and (e) ELISA detection of the indicated cytokines in the supernatants of PBMCs with PCC0208018 at 4 days with anti-CD3 monoclonal antibody (0.5 mg/mL) stimulation.
Materials and methods
Chemicals
PCC0208018 (batch number 20171220 and purity > 98%) was purchased from Hanxiang Biological Technology Co., Ltd. (Shanghai, China), and it is also known as (S)-3-amino-3-(3-((R)-1-amino-2-hydroxyethyl)-1,2,4-oxadiazol-5-yl) propanamide, with a molecular formula of C7H13N5O3 and a molecular weight of 215.21 g/mol. For the in vitro studies, PCC0208018 was dissolved in sterilized water and then diluted in RPMI-1640 medium to the desired concentration.
Cell lines
B16-F10 melanoma cells and CT26 colon cancer cells were obtained from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and maintained at 37°C in 5% CO2 in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and 50 U/mL penicillin-streptomycin (100 μg/mL).
Animals
Male C57BL/6NCrl mice 4–5 weeks old, obtained from Vital River Laboratory Animal Technology Co., Ltd., were employed in the in vivo experiments. The animals were kept in a controlled environment at 25°C on a 12-h light/dark cycle, with food and water provided ad libitum. Each of the experiments related to animals was performed following the Guidelines for the Care and Use of Experimental Animals of the Experimental Animal Research Committee in Yantai University.
MTT assay
The effects of PCC0208018 on cell viability in vitro were evaluated by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assay. 4 B16-F10 and CT26 cells were plated in 96-well microplates at a density of 3 × 103 cells/well overnight and exposed to the indicated concentrations of PCC0208018 for 24, 48, and 72 h, as previously described. 5
Homogeneous time-resolved fluorescence binding assay
Using a PD-1/PD-L1 binding assay kit (Cisbio, Bedford, MA, USA), the binding effect was determined in accordance with the manufacturer’s instructions. BMS936559 was used as the positive control. The homogeneous time-resolved fluorescence binding (HTRF) ratio was calculated using the following formula: HTRF ratio = (A665nm/A620nm) × 104.
Ligand binding assay
PD-L1-overexpressing Chinese hamster ovary (CHO) cells (105 cells/50 µL) were treated with the indicated concentrations of PCC0208018 and BMS936559 for 15 min followed by incubation with recombinant human PD-1-Fc for 15 min. Anti-human IgG Fc-Alexa Fluor 647 was added and then incubated for another 15 min at 4°C. Using an Accuri™ C6 flow cytometer (BD, USA), the ratio of fluorescent cells was determined to calculate the binding efficiencies of PCC0208018 and BMS936559 to PD-L1 according to the formula: PD-L1 binding ratio (%) = (1–Ti/Vi) × 100, where Ti is the ratio of fluorescent cells in the positive control group versus the test group and Vi is the ratio of fluorescent cells in the positive cells versus the blank control group. All reagents were from Jackson Immuno Research (West Grove, PA, USA).
Cells proliferation assay
Human PBMCs (purity > 95%) were provided by LDEBIO Company (Guangzhou, China). We distributed PBMCs (105 cells/well) in 96-well plates with anti-CD3 monoclonal antibody (0.5 mg/mL) and different concentrations of PCC0208018 at 37°C in RPMI-1640 supplemented with 10% FBS, as previously described. 6 After 4 days, cell viability was assessed using the cell counting kit-8 (CCK-8) assay.
Enzyme-linked immunosorbent assay
Cryopreserved PBMCs were seeded (105 cells/well) into 96-well plates and stimulated with anti-CD3 monoclonal antibody and different concentrations of PCC0208018 for 4 days, as described earlier. To investigate the effects of the PI3K/AKT signaling pathway, human PBMCs were pre-incubated with GDC-0941 for 2 h and then stimulated with anti-CD3 monoclonal antibody and PCC0208018 in 96-well plates for 4 days in the presence of GDC-0941. Then, the supernatant was collected and interferon gamma (IFN-γ) and interleukin-2 (IL-2) levels were detected using human enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems, Minneapolis, MN, USA) following the manufacturer’s protocol.
Western blotting
Human PBMCs were treated with 0.5 μg/mL anti-human CD3 monoclonal antibody and varying concentrations of PCC0208018 for 4 days. The cell lysates were denatured and resolved by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto polyvinylidene fluoride (PVDF) membranes. 7 The membranes were blocked with 5% skimmed milk saline for 2 h at room temperature and then incubated with antibodies (1:1000) against p-AKT (Thr308), AKT, mammalian target of rapamycin (mTOR), extracellular regulated protein kinase (ERK), p-ERK (Thr202/Tyr204), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) overnight at 4°C. After several washes, the immunoreactive bands were hybridized with horseradish peroxidase (HRP)-linked anti-mouse or anti-rabbit antibody and examined using the Fluor Chem™ Q chemiluminescent system (Protein Simple Corp., Canada). The grayscale of every band was evaluated utilizing ImageJ software. Protein expression levels were normalized to that of GAPDH. All primary antibodies were from Cell Signaling Technology (Danvers, MA, USA).
B16-F10 tumor-harboring mouse model
B16-F10 tumors were induced by injecting 5 × 105 cells in Matrigel into the dorsal side of the male mice, as previously described. 8 On the second day, the tumor-bearing mice were arbitrarily assigned to four groups (n = 8). The mice intragastrically received 15, 30, or 60 mg/kg PCC0208018 in a volume of 0.1 mL/10 g once daily. Vehicle mice received the identical volume of deionized water. Body weight and tumor dimensions were quantified once every 2 days. Tumor volume was determined using the formula: V = 0.5 × length × width2. On Day 14, the animals were sacrificed and the tumors were excised. The inhibition rate (IR) of tumor growth was estimated using the formula: IR (%) = ((A–B)/A) × 100, where A and B are the mean tumor weights in the vehicle and treatment groups, respectively.
Flow cytometry analysis
Tumor T lymphocytes from the B16-F10-bearing mouse were collected and separated using a mouse tumor dissociation kit (Miltenyi Biotec, Bergisch Gladbach, Germany) and Ficoll-Paque PREMIUM, as previously described. 9 Then, the cells were stained with PE-Cy7 anti-CD3, PE anti-CD4, and fluorescein isothiocyanate (FITC) anti-CD8 monoclonal antibodies in the dark for 30 min in accordance with the manufacturer’s instructions. After rinsing twice with PBS, the stained cells were examined using an Accuri™ C6 flow cytometer, and the data were assessed using the C6 Flow Plus software (BD Biosciences, San Jose, CA, USA). All antibodies were from BD Biosciences.
Immunohistochemical analysis
Tumor expression of PD-L1, CD3, and CD8 was determined by immunohistochemical analysis. The samples were fixed in paraformaldehyde, dehydrated, and embedded in paraffin. The sections (4 µm) were processed for immunohistochemical staining as described previously. 9 Briefly, sections were blocked with 3% normal goat serum and incubated with antibodies against PD-L1 (1:100), CD3 (1:200), and CD8 (1:200) overnight at 4°C and then incubated with biotinylated secondary antibody, followed by avidin–biotin–peroxidase complex. Immunoreactivity signals were developed, and protein-positive cells were stained brown. Sections were examined under high-power microscopy (200×) using the Vectra automated quantitative pathology imaging system (PerkinElmer, Waltham, MA, USA).
Statistical analysis
For statistical comparisons, GraphPad Prism Software was used with one-way analysis of variance (ANOVA) and then Dunnett’s multiple comparison test. The data are presented as the mean ± SD. Statistical significance was set at a threshold of P < 0.05.
Results
Cytotoxicity of PCC0208018 in tumor cells in vitro
Direct cytotoxicity of PCC0208018 was investigated in mouse melanoma B16-F10 cells and colon cancer CT26 cells in vitro. PCC0208018 did not inhibit B16-F10 and CT26 cell survival at 0.005–50 µM (Table 1), which indicated that PCC0208018 did not directly inhibit tumor growth.
The effect of PCC0208018 on CT26 and B16-F10 cells viability (n = 3).

PCC0208018 promoted PBMCs proliferation and cytokine secretion in vitro without blocking PD-1/PD-L1 binding
HTRF binding assay results showed that PCC0208018 did not halt PD-1 and PD-L1 binding at concentrations ranging from 0.5 to 3000 nM, while the PD-L1 antibody BMS936559 showed high blocking activity (Figure 1(b)). Similarly, at the cellular level, PCC0208018 did not block PD-1 and PD-L1 binding from 0.001 to 10 µM (Figure 1(c)). We further investigated the effects of PCC0208018 on PBMCs function. The results showed that PCC0208018 at concentrations of 0.1, 0.3, and 1 µM increased human PBMCs proliferation (Figure 1(d)) and IFN-γ and IL-2 secretion (Figure 1(e)) stimulated by anti-CD3 monoclonal antibody. Thus, our data suggested that PCC0208018 promoted PBMCs proliferation and cytokine secretion in vitro without interfering with PD-1/PD-L1 binding.
Effects of PCC0208018 on PI3K/AKT and MEK/ERK pathways in PBMCs
To elucidate the influence of PCC0208018 on critical signaling pathways, we undertook western blotting and found that PCC0208018 (0.1, 0.3, and 1 µM) increased AKT Thr308 phosphorylation (Figure 2(a)) and mTOR expression (Figure 2(b)), while PCC0208018 at 0.3 and 1 µM enhanced upregulation of ERK phosphorylation (Figure 2(c)) compared with the anti-CD3 group (each P < 0.05). GDC-0941, an effective PI3K/AKT inhibitor, significantly suppressed the increase in IFN-γ production induced by PCC0208018 (Figure 2(d)). These data suggested that PCC0208018 may affect the function of PBMCs by activating the PI3K/AKT and MEK/ERK pathways.

PCC0208018 boosted the activation of the PI3K/AKT and MEK/ERK pathways in PBMCs. (a–c) Western blotting analysis of the expression of p-AKT (Thr308), mTOR, and p-ERK in PBMCs treated with anti-CD3 monoclonal antibody (0.5 mg/mL) and/or PCC0208018 (0.1, 0.3, and 1 µM) for 4 days and (d) ELISA detection of IFN-γ production in PBMCs pretreated with GDC-0941 for 2 h and/or PCC0208018 (0.1 µM) and anti-CD3 monoclonal antibody for 4 days.
Effects of PCC0208018 on tumor development in the B16-F10 tumor model
The antitumor activity of PCC0208018 was examined in melanoma xenografts. PCC0208018 significantly reduced tumor weight (Figure 3(a)) and tumor volume (Figure 3(b)) in contrast to the vehicle group (Day 14, P < 0.05). The data showed that 15, 30, and 60 mg/kg of PCC0208018 resulted in an IR of 32.2%, 37.1%, and 39.5%, respectively. None of the doses induced changes in body weight (data not shown). Then, the proportion of CD3+, CD3+CD4+, and CD3+CD8+ T lymphocytes in the tumors was evaluated by flow cytometry. As shown in Figure 3(d) and (e), the percentages of CD3+, CD3+CD4+, and CD3+CD8+ T cells were increased. Notably, the percentages of CD3+ and CD3+CD8+ T cells in the 60 mg/kg groups were significantly increased in contrast to the vehicle group. Immunohistochemistry was performed to observe the changes in PD-L1, CD3+, and CD8+ T cell expression in B16-F10 tumors (Figure 3(c)). No significant differences in PD-L1 expression levels were observed in B16-F10 tumors between the vehicle and PCC groups, while the expression of CD3+ and CD8+ T cells in the PCC groups was significantly increased compared with the vehicle group. These results suggested that PCC0208018 inhibited tumor development in B16-F10 tumors by improving immunity.

PCC0208018 promoted immunity and inhibited tumor growth in B16-F10 tumor-harboring mice: (a) tumor weights and (b) volumes. (c) Immunohistochemical staining to evaluate the expression of PD-L1, CD3, and CD8 (brown color; magnification 200×). (d) Flow cytometry analysis of the number of CD3+, CD3+CD8+, and CD3+CD4+ T cells in tumors. (e) The representative pictures.
Discussion
Malignant melanoma is an extremely aggressive immunogenic tumor that can influence the immune system to its advantage. Immunotherapy has revolutionized cancer treatment. Compared with immune checkpoint inhibitors, PCC0208018, a small-molecule compound targeting PD-1/PD-L1 interactions, has shown the ability to increase T cell proliferation. T lymphocytes have a major role in immune disease and can secrete different cytokines, which are intercellular signaling proteins that manage homeostasis in organisms and mediate host defense. 10 Our research implied that PCC0208018 enhanced PBMCs proliferation and the secretion of cytokines IFN-γ and IL-2, while PCC0208018 did not interfere with PD-1/PD-L1 binding in HTRF and ligand binding assays.
Recent studies have identified that the PI3K/AKT pathway and mitogen-activated protein kinase (MAPK) axis are involved in T cell activation and cytokine secretion. AKT, as the critical protein, has an important role in protein translation, T cell survival, cell cycle progression, and cell differentiation, and controls the acquisition of effector and memory phenotypes. In our study, PCC0208018 enhanced p-AKT and mTOR expression in PBMCs. Though the AKT results were minimal, the function could be blocked effectively by GDC-0941, a specific PI3K inhibitor, which suggested that PCC0208018 mediated this immunomodulation, at least partially, by the PI3K/AKT signaling pathway. ERK, as the critical intermediate in the MAPK pathway, influences thymocyte differentiation with positive selection and is essential for the proliferation of naïve T cells. 11 PCC0208018 induced p-ERK expression in PBMCs, indicating that the MEK/ERK pathway may mediate the T cell activation by PCC0208018.
Cancer immunotherapy mainly depends on T cells to exert antitumor effects. Tumor-infiltrating lymphocytes are important prognostic factors of melanoma, and higher levels are associated with better outcomes. 12 CD8+ T cells, as the most important tumor-infiltrating lymphocyte cell type to fight cancer, possess a remarkable ability to specifically target and eradicate tumor cells by releasing cytokines such as IFN-γ and IL-2. Our results suggested that the activation of T cells by PCC0208018 elicited antitumor effects.
In summary, PCC0208018 induced PBMCs proliferation and IL-2 and IFN-γ expression via the PI3K/AKT and MEK/ERK signaling pathways and inhibited tumor growth by increasing T cell infiltration. Our findings may contribute to the field of small-molecule therapeutics in immune modulation.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by the National Natural Science Foundation of China (grant no. 81473188), the Key Project of Shandong Province (grant no. 2017GSF218106), and the National Natural Science Foundation of China (grant no. 81503059).