
Abstract
Mechanical loading is recognized to play an important role in regulating the behaviors of cells in bone and surrounding tissues in vivo. Many in vitro studies have been conducted to determine the effects of mechanical loading on individual cell types of the tissues. In this review, we focus specifically on the use of the Flexercell system as a tool for studying cellular responses to mechanical stretch. We assess the literature describing the impact of mechanical stretch on different cell types from bone, muscle, tendon, ligament, and cartilage, describing individual cell phenotype responses. In addition, we review evidence regarding the mechanotransduction pathways that are activated to potentiate these phenotype responses in different cell populations.
Introduction
The bone and surrounding tissues experience many different types of mechanical loading. For example, compressive forces are experienced and tolerated well by articular cartilage. Shear stress resulting from interstitial fluid flow is tolerated in bone tissue and in tendons and ligaments, tensile stretch is tolerated in order to maintain skeletal anatomy. The effect of different forms of mechanical loading of cells has been extensively studied in vitro and in animal models and human subjects, where it is commonly reported to have stimulatory effects on those cells. In fact, fluid flow–induced shear stress actively enhances cellular events that potentiate bone formation and remodeling,1,2 and compressive loading of articular chondrocytes stimulates their proliferation capacity. In this review, we will focus specifically on the impact of mechanical stretch on bone and surrounding cell responses and in particular, the use of the Flexercell system as a means of applying that stretch.
In 1985, the first reported use of flexible-bottomed plates for studying the effect of mechanical stimulation on cell cultures was made. 3 The plates used a computer-controlled vacuum device to apply negative pressure to the underside of the membrane. When the vacuum was applied, the membrane on which firmly adhered cells were attached was stretched downward and consequently, a strain was imparted on the cells adhered to that membrane. A number of parameters under the control of the operator included the magnitude of pressure, waveform, frequency, duration, and incorporation of rest period over the time course of the experiment. As the system is easy to set up and these various parameters are easy to control, the Flexercell system has been a very good platform with which to study the effect of mechanostimulation in many different contexts (Figure 1).
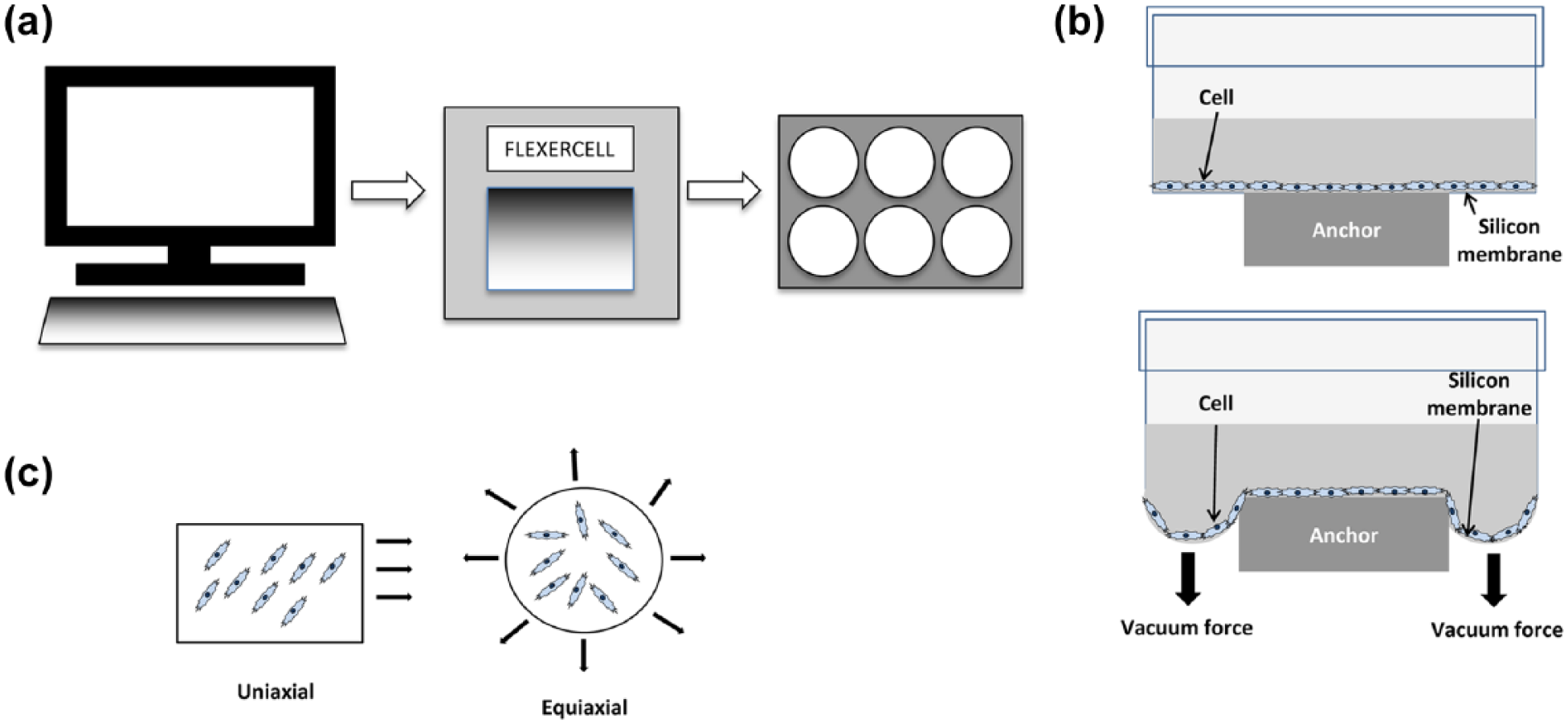
Experimental setup of the Flexercell system for applying strain to cell populations. (a) Schematic representation of the computer-controlled Flexercell system that applies strain to cell monolayers in custom-made six-well plates. (b) Cross-sectional diagram of one well of a six-well Flexercell plate at rest (top) and upon application of the vacuum (bottom). (c) Strain can be applied in a variety of ways including uniaxial and equiaxial.
One huge benefit of the widespread use of this system is that even though the many studies of bone and surrounding cell responses use different strain regimes (magnitude, frequency, duration, etc.), the fact that the base technology applies strain using a common method of application means that some comparability between different studies can be made. Strain is applied either uniaxially or biaxially, generally in two-dimensional (2D) culture (although there are some exceptions to this, where a three-dimensional (3D) culture module is used to characterize compressive force) on silicone membranes coated with type I collagen. Dishes can also be coated with other extracellular matrix (ECM) molecules too making it possible to characterize the effect of mechanical stretch on cells anchored to different ECM components, providing a 2D model of in vivo mechanical stimulation of tissues.
In the bone and surrounding tissues, mechanical signals are continually generated and experienced by the numerous cell types that make up the component tissues. Understanding the role of mechanotransduction and its effects at the cellular level can greatly enhance our understanding of the role of mechanical signals in maintaining normal healthy musculoskeletal physiology and also provide some insight to mechanisms of pathogenesis, for instance, during chronic overuse injury. In the following sections, we review the ever-growing body of the literature that describes bone and surrounding cell responses to mechanical strain applied using the Flexercell system.
Effect of mechanical strain on bone cell phenotypes
During bone loading in vivo, macroscale forces are transmitted to osteocytes through the canalicular network in the form of microscale shear stresses caused by rapid displacement of interstitial fluid. Prolonged cyclic loading of bone leads to increased mineralized tissue density and bone mass. It was hypothesized some time ago that mechanical force was a positive regulator of bone formation creating a shift in bone homeostasis toward the anabolic state, characterized by enhanced bone-forming activity by osteoblasts and a reduction in bone resorption by osteoclasts. 4 In addition to shear stress caused by fluid flow (see above; reviewed by Brindley et al. 1 and Yeatts and Fisher 2 ), tensile stretch applied to cells via physical deformation of culture surfaces5–7 can enhance key cellular and molecular events that lead to bone formation. A summary of notable studies is presented in Table 1.
Summary of Flexercell studies and key findings using osteoblastic cells.

ERK: extracellular-regulated protein kinase; VEGF: vascular endothelial growth factor; bFGF: basic fibroblast growth factor; OCN: osteocalcin; OPN: osteopontin; FAK: focal adhesion kinase; NF-κB: nuclear factor-κB; TGFβ1: transforming growth factor β1; COL: collagen; IGF1: insulin-like growth factor 1; IL: interleukin; PYK2: proline-rich tyrosine kinase 2; COX: cyclooxygenase; M-CSF: macrophage-colony stimulating factor; ALP: alkaline phosphatase; OPG: osteoprotegerin; MEK: mitogen-activated protein kinase (MAPK)-extracellular signal-regulated kinase (ERK) kinase; RANKL: receptor activator of NF-kB ligand; OSX: osterix; RUNX2: Runt-related transcription factor 2; Egr-1: Early growth response protein 1; HIC5: hydrogen peroxide-inducible clone-5.
Flexercell model not specified.
Impact on cell survival, proliferation, and growth
Tensile strain applied using the Flexercell increases osteoblast proliferation5,9,20,28 although the magnitude of strain appears to have a bearing on the degree of proliferation that is permissible. 9 Application of a high strain level of 10%–12% elongation at 0.1 Hz for 24 h stimulated proliferation in primary rat osteoblasts but the even greater mitogenic effect that transforming growth factor β (TGFβ) has in the absence of strain was suppressed by strain itself, 20 suggesting that mechanotransduction competes with TGFβ signaling to dampen its effects. Application of mechanical strain to osteocytes and very mature osteoblasts failed to stimulate proliferation, 29 and it was hence concluded that these cells respond differently to proliferative osteoblasts, instead regulating metabolic responses to strain reflective of their mechanosensing role in vivo.
Lower levels of strain (2.5% elongation at 0.1 Hz) were stimulatory to proliferation of human primary osteoblasts over 48 h, without promoting alkaline phosphatase (ALP) activity or secretion of type I collagen or osteoprotegerin (OPG), suggesting that the cells were maintained in a premature state. However, pre-treating cells with testosterone prior to application of strain promoted osteogenic gene expression at the expense of proliferation, 5 highlighting that the effects of mechanotransduction are subject to a mechanistic switch regulated by androgens.
A study assaying the effects of strain over a range of 0%–9% elongation for 8 h at 1 Hz revealed that proliferation of normal human osteoblasts was magnitude dependent, with the highest strain producing the greatest level of proliferation. 9 Conditioned medium obtained from cultures stretched to 9% elongation stimulated cell proliferation in static culture, indicating the release of a soluble factor.
In summary, the collated evidence indicates that stretch signals have a positive effect on osteoblast proliferation and they do so in a magnitude-dependent fashion. However, there will inevitably be a critical upper threshold at which stretch signals are no longer stimulatory to proliferation and are indeed detrimental to cell viability. Furthermore, biological molecules such as growth factors and hormones may alter the basal cell response to stretch, and this is particularly significant in vivo, where mechanical signals are just one of many facets of the cell microenvironment.
Matrix production
When a range of stretch magnitudes from 0% to 9% was applied to human primary osteoblasts, expression of the type I collagen mRNA, COL1αII, was highest at 9% elongation, whereas in the case of mature osteoblast markers osteopontin (OPN) and osteocalcin (OCN), highest expression was seen at 3% strain, declining thereafter, 9 indicating a biphasic response to tensile strain. This parallels the authors’ observations for proliferation, which was superior at the highest magnitude of 9%. Furthermore, COL1 expression by human primary osteoblasts was only upregulated at a low magnitude of 2.5% strain in the presence of testosterone, 5 further evidence that the effect of mechanical stimulation is subject to hormonal influence. For type III collagen expression, where a “high impact” strain regime of between 5% and 12.5% strain at 0.5 Hz for 24 h on human osteosarcoma cells was used, Liu et al. 21 found that COL3 mRNA was increased at 5% but then underwent a decline in the expression after 5%, whereas COL1 transcripts were stably increased at all strain levels compared to static control cultures. As type III collagen is normally deposited as an early wound matrix component, it is surprising that it was not increased at the higher strain level.
In summary, early matrix production events, that is, deposition of type I collagenous material, appear to preferentially take place at higher strain levels in parallel with the enhanced proliferation effect. This may reflect the wound healing environment in vivo whereby early events of osteoblast proliferation and matrix deposition take place in a mechanically unstable environment prior to the formation of rigid bone, placing large amounts of strain on cells.
Impact on cytokine/growth factor production
Flexercell studies have revealed that numerous growth factors are upregulated in osteoblasts in response to stretch signals. Over a range of stretch from 0% to 9%, vascular endothelial growth factor (VEGF) expression by primary human osteoblasts was highest at 9% stretch. 9 It also seems that the frequency with which stretch is applied influences the form of VEGF synthesized, as 1% stretch applied to both human and rat osteoblasts at various frequencies from 0.05 to 5 Hz revealed that high-frequency stretch promoted the expression of VEGF isoforms that are matrix bound and low frequencies stimulated production of soluble VEGF isoforms. 14 Upregulation of VEGF, along with macrophage-colony stimulating factor (M-CSF) have been reported to be mechanistically important for the recruitment of osteoclasts in the remodeling of bone, 22 and so stretch-induced VEGF expression is pivotal for the regulation of bone remodeling events in vivo as well as in revascularization of regenerating tissue.
TGFβ expression in human osteosarcoma cells was upregulated in response to mechanical stretch applied at 12% elongation. 12 TGFβ is a potent mitogen and can also promote collagen synthesis, both of which are important during the early stages of bone healing. However, binding of TGFβ to the TGFβ receptor-III is also enhanced by mechanical stretch, 20 and this interaction has been shown to inhibit the ability of TGFβ to activate osteoblasts.30–32 Therefore, mechanical strain seems to finely regulate TGFβ signaling and promote similar mitogenic and differentiation responses to TGFβ but via a mechanotransduction pathway that competitively inhibits TGFβ signaling.
Mechanical stretch also enhanced osteoblastic expression of insulin-like growth factor (IGF) and basic fibroblast growth factor (bFGF) at 12% maximal stretch at 3 Hz, over a 24-h period. 12 Both factors stimulate early osteoblast healing responses such as osteoblast proliferation and matrix synthesis33,34 and the early wound healing events induced in osteoblasts by high-magnitude stretch, such as proliferation and type I collagen expression (see above), may be a consequence of increased expression of these factors. Indeed, osteocytes, which act as mechanosensing cell in vivo, produce increasing levels of IGF in response to mechanical strain35–37 which may be one of the primary initiation events for the recruitment of osteoblast precursors during bone remodeling and regeneration.
Increased expression of several bone morphogenetic proteins (BMPs), in particular BMP2, in response to low strain of 1% stretch was also reported in mouse osteoblastic MC3T3-E1 cells, 38 yet in human primary osteoblasts subjected to stretch over a range of 0%–9% elongation, increased gene expression was only seen at 6% and above. 9 However, the overall positive effect of mechanical stretch on osteoblastic expression of BMP2 is further evidence of the stimulatory effect of mechanical stretch signals on osteogenesis.
In terms of the bone remodeling balance, mechanical stretch also seems to have a role in regulation of bone resorption by osteoclasts. Interleukin 6 (IL6) has a well-known role in osteoclast production during normal bone homeostasis. 39 Increased expression of IL6 was observed in human osteosarcoma cells stretched at 12% elongation at 0.05 Hz 12 and in human primary osteoblasts stretched at 2.5% elongation at 0.1 Hz. 5 In the latter study, OPG, an inhibitor of osteoclastogenesis, was produced when stretch was applied in combination with testosterone, but not in its absence, demonstrating the coordinated action of mechanotransduction and hormones. The effects of stretch on IL6 are magnitude dependent, as studies of mouse MC3T3-E1 osteoblastic cells exposed to a regime of 0%–9% elongation revealed that the greatest level of IL6 production was seen at 9% stretch and was dependent on prostaglandin E2 production by cyclooxygenase 2. 23
A key downstream consequence of IL6 activity in osteoblasts that leads to osteoclast activation is production of RANKL. 39 There is some debate in the literature about the effect of mechanical strain on RANKL expression. Some researchers have reported a magnitude-dependent decline in RANKL expression,6,13 and this was accompanied by an increase in the expression of the transcription factors Osterix and Runx2 13 and OPG. 6 However, Kim et al. 19 reported an increase in RANKL expression after exposure to mechanical strain. Nevertheless, in this study, a potential mechanism for preventing osteoclast activity in order to maintain an osteoanabolic process was discovered using MC3T3-E1 cells transfected with RANKL cDNA. The expression of RANKL increased in transfected cells, but when low-level mechanical strain was applied daily for 1 week, most of the protein was in membrane-bound form with a large reduction in soluble RANKL, thus reducing its bioavailability. 19 In any case, RAW264.7 cells that are capable of osteoclastic differentiation in the presence of RANKL failed to do so effectively in the presence of mechanical strain of 10% elongation at 0.5 Hz over 1 week. 40 Together with the strong hormonal influence as demonstrated for testosterone, 5 osteoclast activity is subject to further regulation upon exposure to strain.
Overall, mechanical stretch signals induce the expression of factors that invoke early wound healing events such as mesenchymal cell proliferation and angiogenesis. Furthermore, stretch-induced upregulation of factors such as VEGF and IL6 results in osteoclast recruitment and differentiation, which is in turn carefully regulated by androgens. Studies that have assayed growth factor production over a range of different elongations generally concur that growth factor production is magnitude dependent. This likely reflects the early wound healing response where the fracture site is highly unstable and growth factors are in abundance.
Cellular maturation and differentiation
Osteoblastic differentiation, maturation, and ultimately mineralized matrix deposition can be directly enhanced by mechanical strain applied using Flexercell. For example, stretch applied to immortalized mouse calvarial osteoblasts overnight at 2% elongation enhanced the expression of osteogenic transcription factors, osterix and runx2. 13 Over a range of elongations from 0% to 3.2%, a subsequent study found that runx2 was significantly upregulated only at 3.2% stretch but not at 2.4% or lower, and type I collagen expression was upregulated in proportion to magnitude of stretch. 7 In this study, OCN was only upregulated at 2.4%–3.2% elongation, whereas ALP (expressed earlier in osteogenic differentiation) was upregulated at 0.8%–1.6% elongation but not above.
The strain magnitude-dependent expression of OCN by human primary osteoblasts was further confirmed using an elongation range of 0%–9%. OCN expression was upregulated at 3%, 6%, and 9% elongation compared to 0% control cultures, 9 whereas in the case of osteonectin and OPN, greatest gene expression was seen at 3% and then underwent a gradual decline in basal levels by 9%. 9 At stretch magnitude as low as 1% elongation, bone nodule formation was induced in monolayers of mouse MC3T3-E1 and rat primary osteoblasts.27,41 Particularly intriguing is the fact that some researchers reported that early osteogenic events such as proliferation and collagen production were best at a high strain of 9%, whereas the expression of differentiation markers OPN and OCN was most favorable at low strain of 3% elongation, 9 and bone nodule formation was reported as low as 1% elongation.27,41 Other researchers reported that the osteoanabolic phenotype was enhanced in osteoblasts with increasing magnitude of strain up to 18% elongation, characterized by enhanced expression of OPG, 6 which is a potent inhibitor of osteoclast formation and activity. 42 However, it is not just the magnitude of strain that impacts the specific response of osteoblasts but the timing of application and duration of strain that determine the exact nature of cell responses. According to the work of Visconti et al., 27 applying strain after a prolonged static culture period (either 3 or 10 days) produced a greater number of bone nodules than applying strain after just 1 day of static culture. This suggests that a regime that promotes mineralized matrix formation should be applied after early events such as collagen matrix deposition have been given time to occur.
To summarize the impact of mechanical stretch on osteogenic differentiation and maturation, it is clear that lower magnitude strain in the region of <5% elongation is superior for directing these events, a contrast to early events such as proliferation and type I collagen production, which are optimally enhanced at greater magnitudes of stretch (around 9% elongation).
Effect of mechanical strain on skeletal muscle cells
Mechanical strain is transmitted through tendons and ligaments during locomotion, when muscle contraction occurs. Skeletal muscle itself generates contractile forces that are propagated through the muscle fiber network. The nature of force generation in skeletal muscle means that as well as contraction, cells experience stretch stimuli.
In order to fully critically appraise the information in this area, a brief description of how skeletal muscle differentiates and regenerates is appropriate. The tissue itself is characterized by multinucleate muscle fibers encapsulated by connective tissue, mainly collagen, fixed at both ends (junctions).
In terms of mechano-responsiveness, it could be suggested that skeletal muscle is the most relevant tissue in the human body. It is composed of elongated multinucleate muscle fibers that develop from the “end-to-end” fusing of mononuclear muscle precursor cells (MPCs) in a process known as differentiation. In development, these mononuclear cells are derived from the somites and, depending upon the anatomical location, the MPCs either migrate to differentiate at their final destination, for example, limb muscle or differentiate at or near their final destination, for example, head and neck muscles. However and astoundingly, skeletal muscle is a not a solely post-mitotic tissue. MPCs located in the periphery of post-natal muscle fibers can become activated by a variety of stimuli including mechanical. This activation triggers a series of events that lead to the asymmetric division of these cells. This process can be recapitulated with in vitro cell culture and is illustrated in Figure 2.

A series of processes from MPCs to differentiated myotubes.
It is therefore very important to understand at which part of the process the cells are being stimulated using the Flexercell apparatus where a large number of studies have demonstrated that mechanical strain activates muscle cells at different stages of maturity from skeletal myoblasts to satellite cells,43–47 and a summary of muscle cell studies is presented in Table 2.
Summary of Flexercell studies and key findings using muscle cells.
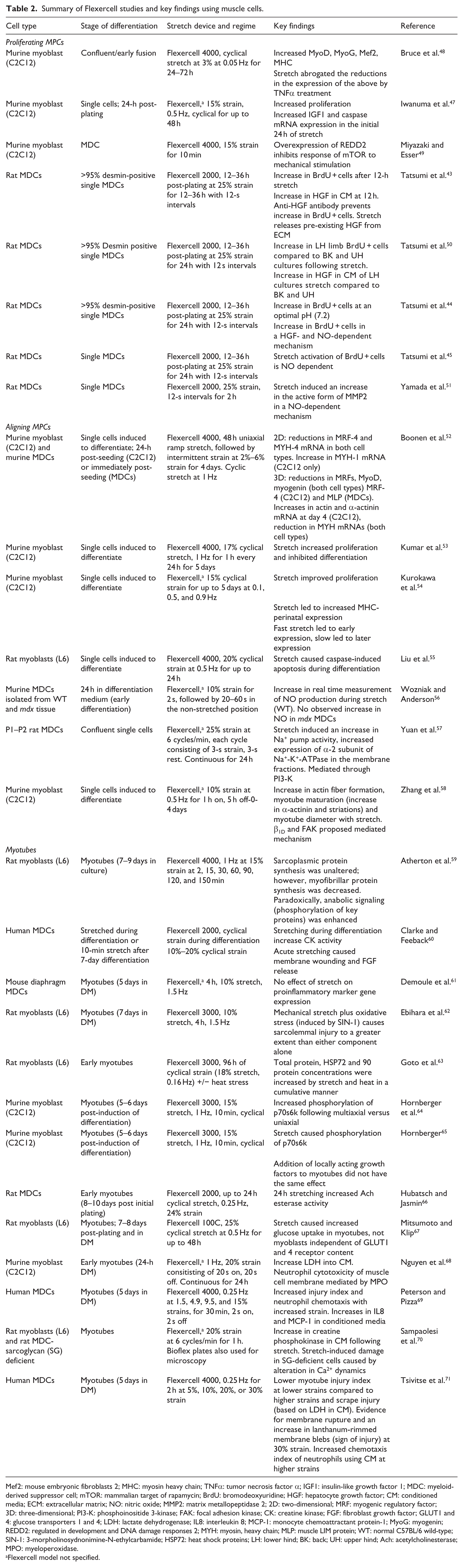
Mef2: mouse embryonic fibroblasts 2; MHC: myosin heavy chain; TNFα: tumor necrosis factor α; IGF1: insulin-like growth factor 1; MDC: myeloid-derived suppressor cell; mTOR: mammalian target of rapamycin; BrdU: bromodeoxyuridine; HGF: hepatocyte growth factor; CM: conditioned media; ECM: extracellular matrix; NO: nitric oxide; MMP2: matrix metallopeptidase 2; 2D: two-dimensional; MRF: myogenic regulatory factor; 3D: three-dimensional; PI3-K: phosphoinositide 3-kinase; FAK: focal adhesion kinase; CK: creatine kinase; FGF: fibroblast growth factor; GLUT1 and 4: glucose transporters 1 and 4; LDH: lactate dehydrogenase; IL8: interleukin 8; MCP-1: monocyte chemoattractant protein-1; MyoG: myogenin; REDD2: regulated in development and DNA damage responses 2; MYH: myosin, heavy chain; MLP: muscle LIM protein; WT: normal C57BL/6 wild-type; SIN-1: 3-morpholinosydnonimine-N-ethylcarbamide; HSP72: heat shock proteins; LH: lower hind; BK: back; UH: upper hind; Ach: acetylcholinesterase; MPO: myeloperoxidase.
Flexercell model not specified.
Impact on cell survival, proliferation, and growth
In the vast majority of studies, it is striking that cyclic stretching of single MPCs upregulates both proliferation and expression of genes and proteins that are required for further differentiation: MYOD1, myogenin (MYOG), MEF2A, p21 (CDKN1A), and IGF1. 47 Furthermore, several genes encoding muscle structural proteins are upregulated, including myosin heavy chain (MyHC) isoforms and α-tropomyosin (TPM1), and this ultimately leads to increased myotube production. 46 Intriguingly, differences in stretch-induced satellite cell responses have been reported from muscles isolated from distinct anatomical regions that contain primarily fast-twitch or slow-twitch fibers. 50 Further evidence of stretch favorably enhancing cell growth, tissue building, and regeneration includes an increase in total protein levels in response to mechanical stretch, 63 and the whole process of stretch-induced cell growth is dependent on mammalian target of rapamycin (mTOR) activation. 72 However, an antagonist, REDD2, is also expressed in skeletal muscle and is capable of inhibiting stretch-activated mTOR activity. 49 Nitric oxide synthase (NOS) becomes upregulated 46 and bFGF expression is increased, also in a strain-dependent fashion. 60 NOS drives the synthesis and release of nitric oxide (NO) from mature muscle myotubes. 56 Together with FGF, NO has well-known mitogenic potential and initiates a “wound healing” response. It does this, at least in part, by activation of satellite cells via matrix metallopeptidase 2 (MMP2)-mediated release of hepatocyte growth factor (HGF) from ECM 51 and subsequently leads to HGF signaling.43–45,73,74 It is particularly noteworthy that single MPCs are very tolerant of a wide range of strain with values ranging from 2% to 25% eliciting similar (sometimes with a dose response) effect.
Impact on the differentiation process and differentiated cells (myotubes)
As MPCs are “preparing” to fuse into multinucleate myotubes, their sensitivity to strain appears to increase 75 although the global responses remain fairly similar—including increased proliferation rates and increased indicators of fusion (Table 2).
In stark contrast to the data seen with single cells, MPCs fused into myotubes respond in quite different ways. One of the most striking observations is that “tolerance” to varying strain rates is lost. Rates nearing in excess of 20% elicited a variety of deleterious effects such as increased creatine kinase (CK) and lactate dehydrogenase (LDH) activity,60,68,70,71 membrane wounding and cell “injury,”60,71,76 inflammatory cytokine and wound repair growth factor release,60,76 sarcolemmal disruption, 62 increases in molecular chaperonins, 63 and increased acetylcholinesterase production. 66 Overall, stretching myotubes appeared to have no real effect on parameters related to cell proliferation and differentiation; however, there is some evidence of changes to cellular metabolism such as increased glucose uptake 67 and phosphorylation of a number of anabolic targets such as ribosomal S6 kinase P70S6K59,72, focal adhesion kinase (FAK), protein kinase B (Akt), 4E binding protein 1 (4EBP1), eukaryotic elongation factor 2 (eEF2), extracellular-regulated protein kinase 1/2 (ERK1/2), eukaryotic initiation factor 2α (eIF2α), and eukaryotic initiation factor 4E (eIF4). 59 Notably, however, myofibrillar protein synthesis was actually suppressed. 59
In conclusion, there is a good deal of data that have been collected, and the system has, for skeletal muscle, some utility for single MPCs work concentrating on some aspects of cellular signaling. There are clearly wider considerations when it comes to interpretation of the myotube data and also around the biomimetic nature of the systems and the interplay with other cell types. 77 The most powerful use is for comparing the same conditions across varying cell types, as described in this review.
Effect of mechanical strain on tendon and ligament cells
Tendon and ligament are important connective tissues, essential for healthy musculoskeletal physiology. Ligaments bind skeletal components together, maintaining their correct anatomical arrangement. Tendons connect muscles to bone and facilitate the transmission of contractile force from those muscles to control movement. They are therefore able to withstand high mechanical strain during normal physiological conditions. However, chronic overuse or pathologically high magnitudes of strain result in damage, causing considerable pain and impaired mobility. Using the Flexercell system, the effect of mechanical strain on cellular constituents of tendons and ligaments can be studied, facilitating our understanding of how these tissues respond to, and propagate, mechanical signals (Table 3). This in turn may help us to understand the kind of mechanical stimulation appropriate for engineering replacement tissues.
Summary of Flexercell studies and key findings using ligament cells.

PDL: periodontal ligament; IL: interleukin; COX2: cyclooxygenase 2; PGE2: prostaglandin E2; MMP: matrix metallopeptidase; TIMP: tissue inhibitors of metalloproteinases; ALP: alkaline phosphatase; ACL: anterior cruciate ligament; MCL: medial collateral ligament; UNCL: ulnar collateral ligament; TGFβ1: transforming growth factor β1; M-CSF: macrophage-colony stimulating factor; OCN: osteocalcin; EGF: epidermal growth factor; EGF-R: epidermal growth factor receptor; BMP: bone morphogenetic protein; VEGFA: vascular endothelial growth factor A; TENS-L: tensile strain of low magnitude; TENS-H: tensile strain of high magnitude; LFC: Ligamenturn flavum cells; C-FOS: FBJ murine osteosarcoma viral oncogene; SOX9: SRY (sex determining region Y)-box 9; MSX1: Msh homeobox 1.
Flexercell model not specified.
Impact on cell survival, proliferation, and growth
One of the earliest studies using Flexercell demonstrated that applied cyclic stretch alone was not capable of increasing flexor tendon proliferation, but when applied in combination with platelet-derived growth factor (PDGF), or a cocktail containing both PDGF and IGF, synthesis of DNA was significantly enhanced. 96 This could be prevented by blocking gap junction activity with the gap junction inhibitor octanol, indicating the importance of cell–cell communication for cell proliferation. 97 Therefore, in terms of proliferation, fibroblastic cells appear not to respond as favorably to mechanical stretch signals as osteoblasts or MPCs. For tissue engineering purposes, it will be necessary to screen the appropriate combination of mechanical stretch and growth factors to create functional tendon tissue with appropriate cell number or seed the scaffold with the desired number of cells required to form mature tissue at the outset.
Matrix production
Mechanical stretch has been widely reported to upregulate type I collagen synthesis by tendon cells97,98 and ligament cells.85,89,90,99 Curiously in the case of the knee joint, type I collagen is upregulated by fibroblasts of the anterior cruciate ligament (ACL)85,99 but not those from the medial collateral ligament (MCL). 85 Conversely, type III collagen is upregulated upon application of strain in MCL cells, but not ACL.85,99 These differences in collagen expression profiles might explain differences in healing of ACL versus MCL evident in vivo, with lack of healing in ACL due to lack of type III collagen production. 85 They also deliver a resounding reminder that seemingly comparable cell populations from different topographic sites can behave rather differently to each other, as demonstrated with topographically distinct skin fibroblast populations that have distinct phenotype and gene expression profiles.100–102
In fibroblasts from ligamentum flavum, mechanical stretch induces the expression of collagen types I, III, and V via a TGFβ-dependent mechanism. 89 In static culture, estrogen increases the expression of collagen types I and III,99,103 but combined with mechanical stretch signals, actually reduces their expression, along with biglycan.99,103 The compound effect of stretch and estrogen might account for the up to eightfold increase in risk of ACL injury in female athletes.104–106
Aside from synthesis of matrix components, it is important to have a controlled balance between proteases that degrade ECM proteins and their inhibitors during matrix turnover and remodeling, and a number of different proteases are upregulated in periodontal ligament (PDL) fibroblasts upon mechanical stimulation. For example, plasminogen activator (PA) is upregulated in mechanically stretched PDL cells.91,107 PA is a wide-spectrum serine protease that activates plasminogen to produce plasmin, which in turn activates MMPs to degrade ECM components and cell adhesion molecules, 108 important events during matrix remodeling, cell mobilization, and differentiation. Furthermore, transcript levels of MMPs themselves (MMP1 and MMP2), along with their inhibitors (tissue inhibitors of metalloproteinase 1 and 2 (TIMP1 and TIMP2)) are upregulated relative to unstretched controls, 79 again indicating that mechanical stimulation enhances matrix remodeling, which is essential for maintaining structural integrity and strength in ligament tissue in vivo. It has also been reported that cell aging leads to increased PA activity, 109 which may reflect the impaired healing capacity that is typically seen in aged individuals.
Exposing tendon cells to IL1 decreased their elastic modulus, thereby increasing their survival after exposure to high-magnitude stretch in 3D collagen gels (see Table 4). 115 Coupled with this, IL1 enhanced elastin expression but suppressed type I collagen expression by tendon cells cultured within the constructs, 114 which will have a bearing on the mechanical properties of bioengineered constructs that are produced this way. Therefore, a potential means of tuning the mechanical properties of engineered tendon tissues for regenerative therapy may be achievable via regulation not just of the biomaterial substrate but also of the tendon cells themselves.
Summary of Flexercell studies and key findings using tendon cells.

IL1β: interleukin 1β; PDGF-BB: platelet-derived growth factor-BB; IGF1: insulin-like growth factor 1; PGE2: prostaglandin E2; VEGF: vascular endothelial growth factor; COX2: cyclooxygenase 2; MMP: matrix metallopeptidase; JNK: c-Jun N-terminal kinase.
Flexercell model not specified.
Impact on cytokine/growth factor production
Growth factors and cytokines can have both autocrine and paracrine effects in the local microenvironment where they are produced. Therefore, the effect of stretch signals applied using Flexercell can give some indication regarding the potential influence of mechanical stretch signals in vivo and also potentially enable “priming” of cells to upregulate particular factors for therapeutic applications.
Stretching PDL cells for 24 h (5% elongation at 0.05 Hz) enhanced TGFβ production after 7 days 84 but only in PDL cells from adult teeth and not deciduous teeth. This perhaps reflects the intrinsic difference in longevity between the two as TGFβ has a prominent role in matrix formation, maturation, and scar tissue formation that is not needed by deciduous teeth that are shed. On the other hand, stretch signals led to a decrease in M-CSF levels, even after pre-stimulation with vitD3, 84 which normally stimulates M-CSF production. VEGF production by PDL cells was also upregulated after stretching,94,95 as were osteogenic factors BMP2 and BMP6. 94 VEGF production by tendon fibroblasts was similarly increased and together with the simultaneous upregulation of PGE2 expression was suggested as a mediator of inflammation and fibrosis, as typically responsible for carpal tunnel syndrome. 113 This suggests that stretch-mediated upregulation of growth factors and cytokines is not necessarily a positive event for fibroblastic cells, and risk of scare tissue formation due to excessive stretch in vivo can perhaps be modeled in vitro using Flexercell systems.
The magnitude of strain seems to be an important factor in determining cell responses. Low-magnitude tensile strain (from 2% to 8%) was found to have anti-inflammatory effects on PDL cells.78,119,120 For example, IL1-mediated transcription of IL1, IL6, and IL8 was blocked, whereas production of IL10, which in turn can block synthesis of IL1, IL6, and IL8, 121 was enhanced. 119 In this low range, strain was also found to significantly inhibit IL1-mediated activation of COX2 gene expression and PGE2 synthesis in a dose-dependent fashion, 78 reinforcing the argument that low-magnitude strain is anti-inflammatory. In contrast, high strain levels (10%–18.5% strain) induce pro-inflammatory responses such as nuclear factor-κB (NF-κB)-mediated COX2 gene expression. 78 Moreover, high strain levels (9% and 18%) significantly enhanced production of IL1 and PGE2 and the effects were more prominent in PDL cells from aged tissue versus young. 92 Cellular aging was also found to impact significantly COX2 gene expression, with aged (late passage) PDL cells producing higher COX2 mRNA levels than young cells after strain. 122
High strain levels (18% elongation applied for 12 h at 0.1 Hz) were also found to significantly enhance NO production, 123 which has many physiologic functions that include inflammation, which is likely in this case given the other reported inflammatory mediators upregulated by high-magnitude strain. In rabbit Achilles tendon fibroblasts, the application of strain in the presence of IL1 also led to a 17-fold increase in pro-MMP3 production, but no difference in COX2 expression was observed compared with IL1 treatment alone. 124 Strain-induced expression of inflammatory genes could be suppressed by the addition of exogenous ATP to the culture. 117
Overall, it seems that a growth factor/cytokine expression profile associated with inflammation is produced in response to mechanical stretch signals that is magnitude dependent. However, low-magnitude stretch is actually anti-inflammatory, and this biphasic response runs parallel with physiologic scale observations where chronic overuse results in injury and inflammation.
Cellular maturation and differentiation
Functional tendons are dependent on adequate cell–cell communication and gap junctions are crucial for this. 97 Specific gap junction proteins n-cadherin and vinculin are both significantly upregulated in tendon cells after exposure to cyclic stretch 116 along with connexin-43, which is localized to gap junctions at the ends of actin filaments. 118 Enhanced cell–cell communication is very important for stretch-mediated responses, particularly so in 3D constructs populated with tendon fibroblasts, where application of mechanical strain using a non-Flexercell method resulted in a threefold increase in tensile strength of constructs compared with non-loaded constructs after 7 days of loading. 125 In addition, supplementing cultures with the anabolic steroid, nandrolone decanoate, synergistically enhances matrix remodeling and ultimately tensile strength of bioengineered tendons. 126 Therefore, applying mechanical strain to tissue engineered constructs creates a more physiologically relevant tissue. Moreover, when considering information gained from many published Flexercell studies, translating the relevance of this information from 2D to 3D microenvironment needs to be considered.
PDL cells can acquire a bone-like phenotype, and mechanical strain is generally considered to promote and enhance this. Wescott et al. 94 subjected PDL cells to 12% cyclic strain for up to 24 h and then screened an array of 78 different genes associated with osteogenic differentiation and bone metabolism. Several genes associated with osteogenesis were upregulated, including BMP2, BMP6, ALP, SOX9, MSX1, and vascular endothelial growth factor A (VEGFA), suggesting that a program of osteogenic gene expression was invoked. However, there seems to be a minimum duration of exposure to strain, or culture period following exposure, that is required for osteogenic induction to take place as short-term exposure (6 h) was insufficient to enhance the expression of any key osteogenic genes.93,94 Over a longer culture period of 48 h, the expression levels of type I collagen and decorin were elevated after stretch of up to 18% elongation. 90 Continuous cyclic strain of 9% or 18% elongation at 0.1 Hz applied to PDL cells for up to 6 days resulted in enhanced osteoblastic differentiation characterized by increased ALP activity and expression of OCN, with paralleled decrease in mitogenic potential and epidermal growth factor (EGF) receptor expression.88,127 Differentiation was inhibited by the addition of EGF to the culture medium, which reversed the profile.
In a separate study, ALP synthesis was downregulated in PDL cells subjected to cyclic stretch of 15% elongation at 0.5 Hz and even treatment with vitamin D (an agonist of ALP production) was unable to compensate. 80 Therefore, the PDL contains a source of multipotent cells that can differentiate toward a bone-forming phenotype, and a careful balance between both biological and mechanical signals is required to maintain the niche and direct cell fate. Furthermore, at high strain levels it seems that a fine balance between osteogenic differentiation and inflammation occurs. Similar to osteoblasts, in PDL cells, cyclic strain also suppresses osteoclastogenesis as although osteoclast genes OPG and RANKL are both upregulated, 128 the former inhibits the stimulatory effects of the latter.
In summary, mechanical stretch signals applied to tendon and ligament cells using Flexercell have been reported to be largely positive, with matrix synthesis and remodeling being upregulated when using moderate strain levels. Moderate strain levels also have documented anti-inflammatory properties and are able to abrogate the effects of cytokines on these cell types. Conversely, high strain levels induce inflammatory pathways and this likely reflects the potential tissue damage caused by high strains in vivo.
Effect of mechanical strain on cartilage cells
Hyaline cartilage has a crucial protective role in the bone–bone contacts, such as knee, where it protects the ends of opposing bone structures and dissipates loads evenly across the full surface area of the cartilage. Consequently, in addition to the load bearing that is typically associated with cartilage function, the constituent chondrocytes also experience tensile and shear stresses.129–131 A summary of studies assessing the impact of mechanical stretch on chondrocytes using Flexercell is presented in Table 5.
Summary of Flexercell studies and key findings using chondrocytes.

IL1β: interleukin 1β; NF-κB: nuclear factor-κB; MMP: matrix metallopeptidase; iNOS: inducible nitric oxide synthase; NO: nitric oxide; TNFα: tumor necrosis factor α; COX2: cyclooxygenase 2; PG: proteoglycan; PKC: protein kinase C; ROS: reactive oxygen species; TMJ: temporomandibular joint; PGE2: prostaglandin E2; TNFR: tumor necrosis factor receptor; IκB: nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor; HA: Hyaluronan.
Flexercell model not specified.
Impact on cell survival, proliferation, and growth
Chondrocyte proliferation is upregulated in response to strain. 151 It is not just magnitude of strain that can impact cell responses, but also frequency. This was demonstrated when mechanical stretch of 3% elongation was applied to chondrocytes at 2, 30, or 150 cycles/min (corresponding to 0.03, 0.5, or 2.5 Hz), with the higher frequency increasing DNA synthesis. 152 From this observation we can conclude that a delicate interplay between several factors including magnitude and frequency of applied stretch can influence cell behavior.
Matrix production
Multiple studies using hyaline cartilage have concluded that application of mechanical stretch increases production of the major collagen component of cartilage, type II collagen,139,140,152,153 and the integrin subunit α2,139,153 which binds to collagen. Similarly, the proteoglycan aggrecan is also upregulated140,143,152,153 as are several important cartilage glycosaminoglycans (GAGs) including chondroitin-6-sulfate, hyaluronic acid, and dermatan sulfate. 154
However, not all studies agree as stretch was found to downregulate transcript levels of aggrecan and type II collagen, but this could be reversed by the addition of IL4. 144 Hyaluronic acid was also found to undergo depolymerization in the presence of strain due to generation of reactive oxygen species. 146 These conflicting observations can perhaps be explained by the recurring theme of magnitude-dependent effects. In the case of matrix production, particularly proteoglycans that along with their component GAGs are important for producing a highly hydrated matrix, the effects of stretch are magnitude dependent. Low-magnitude strain enhances proteoglycan production and matrix synthesis, whereas high-magnitude strain diminishes it resulting in ECM degradation and onset of inflammation.132,133,135,136,155 The process is dependent on protein kinase C activity. 136 Moreover, application of high-magnitude cyclic strain to cultured chondrocytes was found to upregulate the expression of MMPs that degrade cartilage ECM.133,135,155 Cyclic strain is able to prevent suppression of proteoglycan synthesis caused by IL1138,142 and tumor necrosis factor α (TNFα) 87 but again may be magnitude dependent.
Impact on cytokine/growth factor production
In terms of inflammation, the same biphasic response as seen for tendon/ligament cells can be observed. Exposure of chondrocytes to low magnitudes of strain (<8% elongation) is reported to invoke anti-inflammatory effects, whereas high strain levels (up to 18% elongation) augment the inflammatory response. 132 This indicates some conservation in responses to different cell populations from throughout the musculoskeletal system with respect to mechanical signal responsiveness. In particular, low-level strain has generally been observed to block the expression of inflammation-related genes that augment cartilage destruction.87,132,134,137,142,147,149,150 For example, using high frequency and low strain (3% elongation at 25 MHz), IL1 pro-inflammatory activity was inhibited, due to block of IL1-mediated NF-κB nuclear translocation, 134 in a transforming growth factor-β-activated kinase-1 (TAK1)-dependent fashion. 156 This prevented subsequent transcription of inflammatory gene targets, which normally result from NF-κB activation. Similarly, TNFα-mediated inflammatory responses were blocked by strain in vitro. 87
There are some anomalies to this general trend however, with high-magnitude strain (20% elongation) applied at low frequency (0.05 Hz) reported to produce anti-inflammatory effects by downregulating IL1-dependent inducible NOS production. 137 Additionally, cyclic strain of 7% elongation at 0.5 Hz was found to induce IL1β expression and that of MMP13 and cathepsin B, 133 both proteolytic enzymes associated with cartilage pathogenesis.
Interestingly, one study reported that in the presence of a single strain regime of 10% elongation at 0.5 Hz, both catabolic/pro-inflammatory and anabolic responses were observed. 140 While production of NO, PGE2, and COX2 (characteristic of a pro-inflammatory response) were increased, so too were expression levels of type II collagen and the cartilage-specific proteoglycan aggrecan, as mentioned above. This early response was then replaced by elevations of expression of TGFβ1 and TGFβ3, along with MMP1, indicating that matrix remodeling ensues. Collectively, the above data echoes at the cellular level the patient response where gentle exercise and joint mobility are reported to be beneficial for inflamed joints.157,158
In summary, Flexercell studies indicate that low to moderate levels of strain generally promote cartilage anabolic responses, terms of chondrocyte proliferation, GAG, and proteoglycan production, while also conferring anti-inflammatory effects. However, high levels of strain have a tendency to drive inflammatory responses and provide a good deal of correlation to the trauma and injury associated with high strain and overuse in patients.
The effect of strain applied using Flexercell on mechanotransduction mediators
Most of the bone-stimulating effects of mechanical strain occur due to the activation of mechanotransduction pathways, and the major cell surface receptors involved in these pathways are the integrins. In human osteosarcoma cells, a strain regime of 20 kPa at 0.05 Hz resulted in the upregulation of β1 integrin mRNA expression11,28 along with redistribution of β1 integrin subunits from the cytoplasm to the cell membrane. 11 However, αv integrin expression and distribution were unaffected by mechanical strain, 11 indicating that functional activation and continued recruitment of integrins in response to strain are specific to certain subunits of integrin. Interestingly, vitronectin and OPN, both ligands for αv-containing αvβ3, induce greater calcium influx into osteoblasts than other matrix molecules, 159 and the lack of change in αv integrin subunit expression and distribution upon mechanical loading using Flexercell suggests that the cell response to this form of mechanical strain is not calcium-dependent. It has also been reported that both α2β1 and α5β1 can transmit the effect of stretching from the extracellular environment to activate ERK downstream. 24 However, it seems that the α-subunits are the most potent mediators, not β1, and their signal transduction capacity is actin dependent.24,160 Disruption of the actin cytoskeleton, for example, by treatment with cytochalasin D, an actin-depolymerizing agent, attenuates stress response pathways in osteoblast cells.161–163 This indicates the importance of the cytoskeleton in sustaining mechanotransduction from extracellular input signals.
Mechanotransduction downstream of integrins involves the coordinated regulation of several other key molecules. Src kinase interacts with integrin intracellular domains 24 and FAK 10 at the focal adhesion site when mechanical strain is applied. In osteoblasts, this universally leads to stretch-induced activation of mitogen-activated protein kinase (MAPK) pathways and in particular ERK, a key MAPK effector molecule that activates mechanoresponsive transcription factors.7,8,10,13,15,18,24,26,164 At the focal adhesion site formed upon mechanical activation of integrins, FAK also undergoes enhanced and sustained association with another tyrosine kinase, proline-rich tyrosine kinase 2 (PYK2),10,16 which seems to stablize FAK and remove the inhibitory Hic5 adaptor protein from the focal adhesion complex upon activation. 16 Collectively, these data confirm that integrins and their associated intracellular partners are necessary for signal transduction downstream of the strain stimulus to be executed.
Canonical wnt signaling in Flexercell-stretched osteoblasts is highly activated, with enhanced β-catenin transcriptional activity reported.17,25 This leads to upregulation of numerous wnt target genes such as those encoding cell cycle proteins, the gap junction protein connexin-43, and other transcription factors. 25 Phosphorylation of AKT can also promote β-catenin transcriptional activity via glycogen synthase kinase 3 (GSKβ) inactivation and subsequent translocation of β-catenin to the nucleus, where the wnt target genes Wisp1 and Cox2 are then transcribed. 165 Upregulation of the wnt target gene encoding connexin-43 seems to have a role in enhancing cell–cell communication among mechanically stimulated osteoblasts by enhancing gap junction communication due to increased levels of phosphorylated connexin-43 protein,166,167 and in tendon cells its colocalization with actin presumably facilitates cell communication in the direction of force application.
The role of integrins in mechanotransduction is explicit as they anchor cells to the ECM substrate in which they reside.168,169 Less obvious candidates for mechanotransduction are cell surface receptors that do not have an active role in physical anchorage of cells to ECM. It is well known that androgens, in combination with mechanical cues, can control bone metabolism and invoke a shift from high turnover to osteoanabolic mode. Typically, mechanical signaling via the estrogen receptors (ERs) enhances osteoblast proliferation. 170 ERs have a critical role in mechanotransduction and ERK activation, as osteoblasts from ERαβ−/− mice lacking both α and β forms of ER were unable to activate ERK when stimulated with 5% elongation at 0.05 Hz. 8 This phenotype could be partially rescued by transfecting ERαβ−/− osteoblasts with cDNA for either receptor or fully rescued by transfection with cDNA for both receptors. Additionally, the active estrogen compound estradiol acts in synergy with mechanical strain to promote ER-dependent ALP activity. 171 U2OS osteosarcoma cells that do not express ERs and have intrinsically low ALP activity were compared with transfected U2OS cells expressing either ERα or ERβ for COL1 expression and ALP activity after being subjected to 1.5% cyclic strain at 0.05 Hz. Mechanical strain increased ALP activity and decreased COL1 expression in ERα and ERβ mutants, with estrogen having a synergistic effect on ALP activity. However, estrogen itself is dispensable to the process of ER-mediated mechanotransduction as transfecting ERαβ−/− osteoblasts with mutant ERs incapable of binding to estrogen did not impair ERK activation, demonstrating that the process was not dependent on estrogen. Even so, it cannot be discounted that differences in the cell lines or the strain regimes applied to cells may have a bearing on the outcome.
Integrins that mediate cell adhesion initiate mechanotransduction signals and have shown significant impact on ligament cells upon stretch stimulation. In PDL fibroblasts, assessment of integrin gene expression showed that both α6 and β1 were upregulated by stretch. 79 Cells from the ACL or MCL plated onto type I collagen, fibronectin, or laminin express elevated levels of both α5 and β1 when stretched at 5% elongation for 2 h, 81 and in avian tendon cells, src kinase phosphorylation is also increased after mechanical stimulation. 96 In 3D collagen gels loaded with ACL fibroblasts, strain enhances both α5 and β1, but not in α1 expression. 82 However, the expression of the α5 integrin subunit in PDL cell monolayers subjected to stretch was actually found to be downregulated upon application of 20 kPa negative pressure to each well for 12 h. 79
As well as integrin-mediated mechanotransduction, calcium signaling has an important role in cell responsiveness to mechanical forces. Calcium signaling between MCL cells is enhanced by mechanical stretch, as determined by their ability to maintain a calcium wave between cells both aligned longitudinally in microgrooves and in non-aligned cells. 86 In the case of tendon cells, activation of JNK, a “stress-activated protein kinase,” occurs in response to mechanical strain, and the level of JNK activation is dependent on magnitude of strain applied. 111 It also seems to take place downstream of calcium signaling as increased intracellular calcium levels also enhance JNK activation independent of strain. 111 Chronic, sustained activation of JNK in tendon cells could result in cell death and tendon injury.
There was also prominent upregulation of ulnar collateral ligament (UNCL) protein in PDL cells after applied strain. 83 UNCL has a central role in mechanotransduction pathways based on evidence that loss-of-function mutations to the homolog in Drosophila result in failure of mechanotransduction 172 and so might have important function in ligament physiology.
Summary
Cell behavior is regulated by both biochemical and physical signals. The importance of biophysical cues in determining cell phenotype is becoming more widely understood, as are mechanotransduction pathways that are activated upon mechanical stimulation. Application of mechanical strain, most commonly applied using the Flexercell system, can provide physical stimulation to cells in the bone and surrounding tissues and aid our understanding of mechanical strain impact on health and injury. It may also potentially be used as a means of priming cell components of the tissues prior to seeding them into scaffolds for tissue engineering and in vivo transplantation.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Priority Research Centers Program (Grant No. 2009-0093829), the Global Research Laboratory Program (Grant No. 2015032163) and KRIBB Research Initiative Program, Republic of Korea.