
Abstract
Introduction
Osteoarthritis (OA) is more common in women, suggesting a link to estrogen. Estrogen supports cartilage homeostasis and metabolism via two receptors: ERα and ERβ. Mechanical stress is the primary cause of OA and affects cellular functions, including receptor activity. The link between excessive mechanical stress and ERs in cartilage is poorly understood. This study examined ER expression changes in chondrocytes under mechanical stress and the role of estrogen in cartilage degradation and ER regulation.
Methods
ATDC5 chondrocytes were treated with 17β-estradiol (E2; 1.0 × 10⁻⁷M) and subjected to cyclic tensile strain using the Flexcell Tension Plus FX-2000. Total RNA and proteins were extracted, and IL-1β and MMP-13 expression were analyzed using RT-PCR and western blotting. RT-PCR was used to examine the expression of ERα and ERβ.
Results
The CTS group had significantly downregulated ERα expression at all time points and upregulated ERβ expression after 3 and 6 hours compared with controls. In the E2+CTS group, IL-1β and MMP-13 expression were suppressed after 3 and 6 hours, coinciding with significant upregulation of ERβ compared with the CTS group.
Conclusions
Estrogen exerts a chondroprotective effect under excessive mechanical stress, suggesting that ERβ mediates this effect.
Introduction
Osteoarthritis (OA) is a degenerative joint disease characterized by the breakdown of articular cartilage, leading to pain and impaired mobility. Multiple factors are believed to contribute to the onset and progression of OA, including genetic predisposition, environmental influences, trauma, and hormonal changes.1,2 The incidence of OA is notably higher in women, and a potential association with estrogen has been suggested. In women who are postmenopausal, the prevalence of OA increases markedly as estrogen secretion declines, indicating that estrogen may play a protective role in maintaining joint homeostasis.3,4 Conversely, OA has also been reported in adolescent females, 5 suggesting that hormonal fluctuations during puberty and menopause may be linked to OA exacerbation; however, the underlying mechanisms remain unclear.
Estrogen is a steroid hormone involved in a wide range of physiological processes, including reproductive development, bone metabolism, cardiovascular regulation, immune responses, and neural function. 6 These effects are primarily mediated by binding to intracellular estrogen receptors (ERs). There are two major ER subtypes: ERα and ERβ. Upon binding to estrogen, these receptors translocate to the nucleus and regulate the transcription of specific target genes. ERα is the predominant receptor in many tissues, whereas ERβ has been reported to modulate ER signaling by antagonizing ERα in specific contexts. 7 Notably, ERs are implicated in tissue homeostasis and degenerative processes in bone and cartilage, and recent studies have highlighted the critical involvement of ER expression and function in OA pathogenesis. 8
Articular cartilage is routinely subjected to moderate mechanical stress during daily activities; however, excessive mechanical loading can trigger cartilage degeneration and inflammatory responses, which are considered key contributors to OA development.9,10 Mechanical stress, such as compression and shear forces, induces the expression of the proinflammatory cytokine interleukin-1β (IL-1β), which subsequently upregulates matrix metalloproteinase 13 (MMP-13), leading to cartilage degradation.10,11 Mechanical stress is also associated with ER regulation. For instance, mechanical loading has been reported to increase ERβ expression in rat tendon cells, 12 whereas it significantly downregulates ERα expression in osteoblasts. 13 These findings suggest that mechanical stress may alter ER expression in chondrocytes, thereby influencing estrogen signaling and contributing to cartilage degradation in OA. However, few studies have investigated the relationship between mechanical stress and ER expression in chondrocytes or its impact on estrogen-mediated effects.
This study aims to elucidate the changes in ER expression in chondrocytes under conditions of excessive mechanical stress and to explore the role of estrogen in cartilage degradation, with a particular focus on the regulatory mechanisms of ER expression.
Materials and Methods
Cell Line and Culture Conditions
Mouse ATDC5 chondrogenic cells were obtained from RIKEN Cell Bank (Tsukuba, Japan). ATDC5 cells were seeded at a density of 6 × 10⁴ cells/ well in Flexcell dishes (Flexcell International, Hillsborough, NC, USA) that were precoated with 25 μg/mL type II collagen (KOKEN, Tokyo, Japan). Cells were maintained in 2 mL DMEM/Ham’s F12 hybrid medium (Sigma, St. Louis, MO, USA) containing 10% fetal bovine serum (Bio Whittaker, Verviers, Belgium), 10 mg/mL human transferrin (Sigma), and 3 × 10⁻⁸ M sodium selenite (Sigma) in a humidified incubator with 5% CO₂ at 37℃. When the cells reached approximately 50% confluence, the culture medium was replaced with DMEM/Ham’s F12 hybrid medium containing 10 μg/mL bovine insulin (Sigma) and 37.5 μg/ml ascorbate-2-phosphatase (Wako Pure Chemicals, Osaka, Japan) to induce hypertrophic chondrocytes differentiation. Based on previous studies showing high expression levels of collagen type X and ERs in hypertrophic chondrocytes, 14 cells were cultured in differentiation medium for 14 days. To investigate the effect of estrogen, the cells were treated with 17β-estradiol (E2; Cayman Chemical, Ann Arbor, MI, USA). Prior to the addition of 17β-estradiol, the cells were transferred to serum-free medium and incubated for 24 hours. Based on previous reports indicating that 10⁻⁷M falls within the physiological concentration range in vivo, 15 the cells were treated with 17β-estradiol at a final concentration of 10⁻⁷M.
Application of Cyclic Tensile Strain (CTS)
ATDC5 cells were stretched using the Flexcell Tension Plus FX-2000 system (Flexcell International, McKeesport, PA, USA). It has previously been reported that loading 3% strain (0.17 Hz and 2 hours) on chondrocytes caused only slight or no biological reactions, however, loading at 3% to 10% strain (0.17-0.5 Hz, 2-12 hours) has been reported to induce anabolic responses. 16 We have investigated the effects of mechanical loading on chondrocytes in vitro and found that mechanical strain (10% elongation, 0.5 Hz, for 3, 6, and 12 hours) induced catabolic responses, including upregulation of inflammatory cytokines and matrix metalloproteinases. This condition was therefore defined as “excessive mechanical stress.”11,17,18 The apparatus was maintained at 37℃ in a humidified atmosphere with 5% CO₂. This computer-controlled apparatus can program a biaxial strain across laminin-coated culture wells.
Real-Time Polymerase Chain Reaction (RT-PCR)
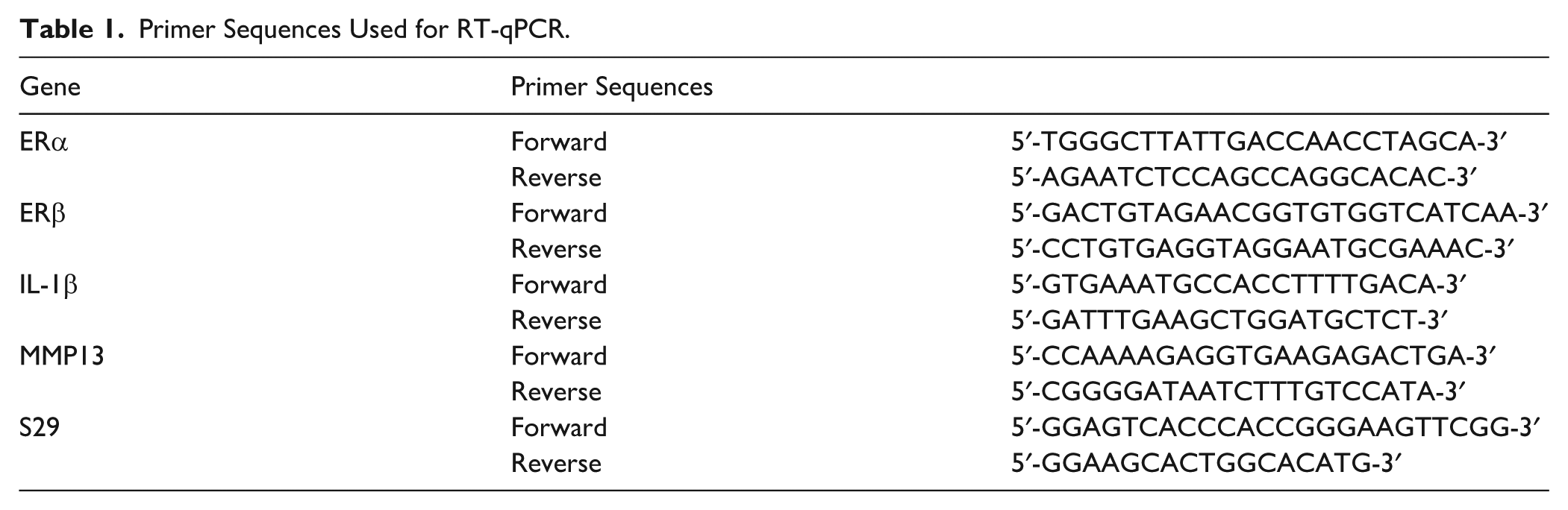
Total RNA was extracted from cultured ATDC5 cells using TRIzol reagent (Invitrogen Life Technologies Inc.) according to the manufacturer’s instructions. cDNA was synthesized using ReverTra Ace qPCR RT Master Mix (Toyobo, Osaka, Japan). Quantitative PCR (qPCR) was performed using the THUNDERBIRD SYBR qPCR Mix (Toyobo) on a LightCycler System (Roche Diagnostics, Mannheim, Germany) to determine the expression levels of the target genes. The primer sequences are listed in Table 1. All reactions were performed in triplicate, and mean values were used for analysis. Relative gene expression levels were normalized to those of S29.
Primer Sequences Used for RT-qPCR.
Western Blot Analysis
Total protein was extracted from each well using Triton buffer (50 mM Tris, 250 mM NaCl, 0.1% Triton X-100, 1 mM EDTA, and 50 mM NaF) supplemented with 1% (v/v) protease inhibitor cocktail and 1mM sodium orthovanadate. Protein concentrations were determined using the BioRad protein assay (BCA Protein Assay Kit, Thermo Fisher Scientific, Waltham, MA, USA). Equal amounts of protein were separated by electrophoresis on polyacrylamide gels (e-PAGEL, ATTO, Tokyo, Japan) and subsequently transferred onto polyvinylidene difluoride membranes using the iBlot 3 Gel Transfer Device and iBlot 3 Transfer Stacks (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. The membranes were blocked with 1% skim milk for 30 minutes at room temperature(approximately 22℃-25℃) and then incubated overnight at 4℃ with primary monoclonal antibodies: anti-IL-1β (CST), anti-MMP13 (CST), or anti-β-actin (Wako, Osaka, Japan).
Immunocytochemistry (ICC) Analysis
After 14 days for differentiation induction, ATDC5 cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature. Following fixation, the cells were washed 3 times with phosphate-buffered saline (PBS) and permeabilized using 0.1% Triton X-100 in PBS for 15 minutes. Nonspecific binding was blocked by incubating the cells with 1% bovine serum albumin (BSA) in PBS for 1 hour at room temperature. Cells were incubated overnight at 4℃ with primary antibodies diluted in BSA solution: anti-ERα (1:200) and anti-ERβ (1:200). After washing with PBS, the cells were incubated with fluorophore-conjugated secondary antibodies (Alexa Fluor 488-labeled anti-rabbit IgG, Life Technologies) for 30 minutes at room temperature in the dark. After removing the secondary antibody, the nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). All images were captured using a fluorescence microscope equipped with image analysis software (BZ-X800; Keyence Corporation, Osaka, Japan).
Statistical Analysis
All experiments were performed independently at least 3 times unless otherwise indicated. Data are presented as the mean ± SD. Prior to statistical analysis, the normality of the residuals and the homogeneity of variances were tested using the Shapiro–Wilk and Levene’s tests, respectively. To satisfy these assumptions, data were log-transformed where necessary. Statistical analyses were conducted using one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison test (JMP version 19.0; SAS Institute, Cary, NC, USA). A P < 0.05 was considered statistically significant.
Results
Immunofluorescence Staining of ERs in ATDC5 Cells
Immunofluorescence staining demonstrated the expression of ERs in ATDC5 cells. Both ERα and ERβ were localized in the nuclei and cytoplasm of hypertrophic chondrocytes (
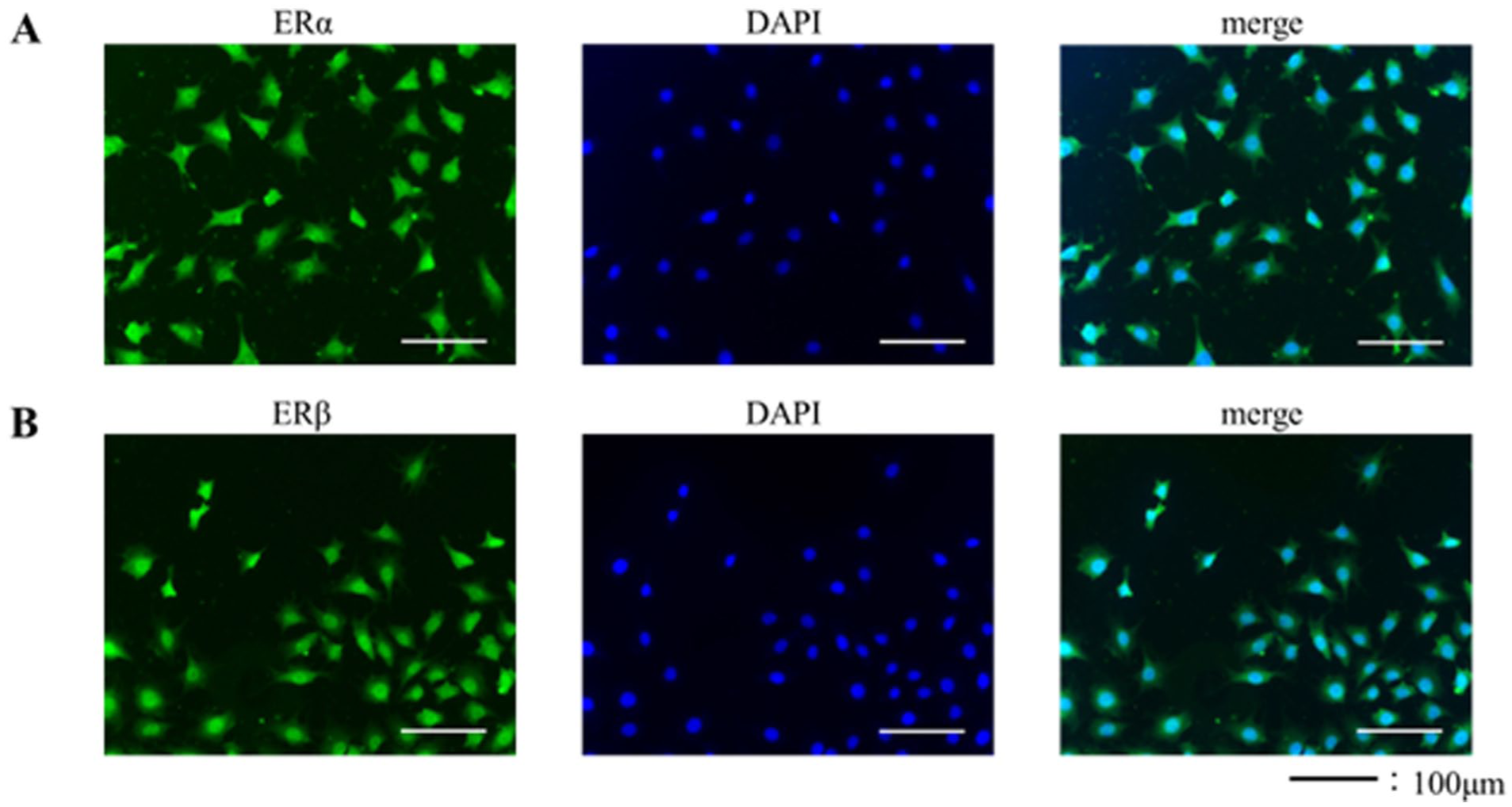
Immunohistochemistry for ERα and ERβ in ATDC5 cells.
Effect of Estrogen on Cartilage Degeneration Induced by Excessive Mechanical Stress
We investigated the effect of estrogen on chondrocyte degeneration induced by cyclic tensile strain (CTS). Compared with the CTS group, the E2+CTS group exhibited a significant downregulation of IL-1β mRNA expression at 3 hours (P < 0.001) and 6 hours (P = 0.0149)
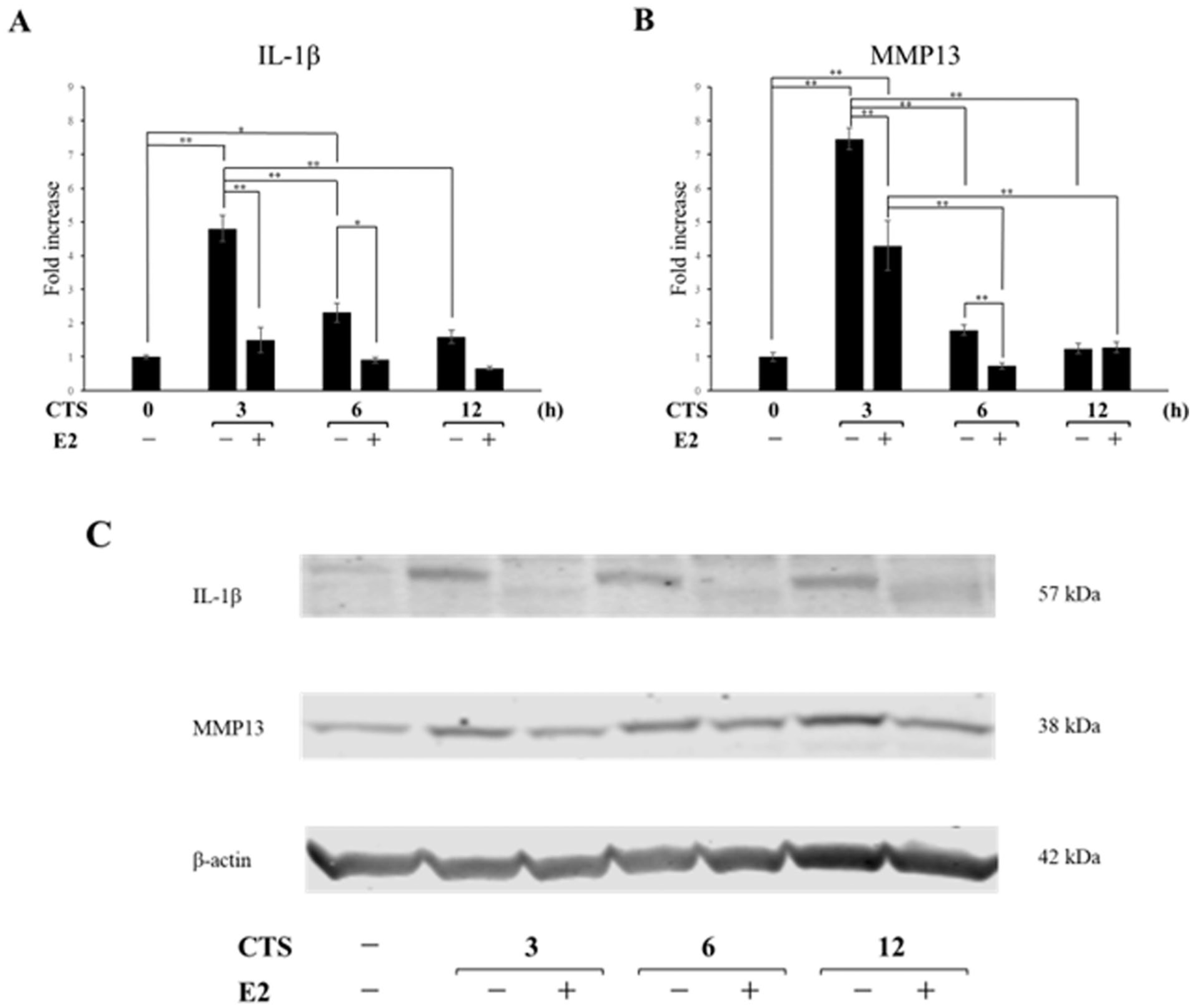
Effects of E2 on CTS-induced expression of proinflammatory factors.
Expression of ERα and β in ATDC5 Cells Treated With Estrogen
We investigated the changes in ER expression in ATDC5 cells following estrogen treatment. RT-qPCR demonstrated that ERα mRNA expression in the E2 group remained significantly lower than in the control group at all time points (all P < 0.001,
Effects of E2 on the gene expression of ERs. Chondrocytes were treated with E2 for 3, 6, and 12 hours. At each time point, ERα and ERβ mRNA levels were quantified by real-time RT-PCR and normalized to S29. Data are expressed as mean ± SD from three independent experiments (n = 3). E2 = estradiol; CTS = cyclic tensile strain; ERα = estrogen receptor α; ERβ = estrogen receptor β. *P < 0.05, **P < 0.01.
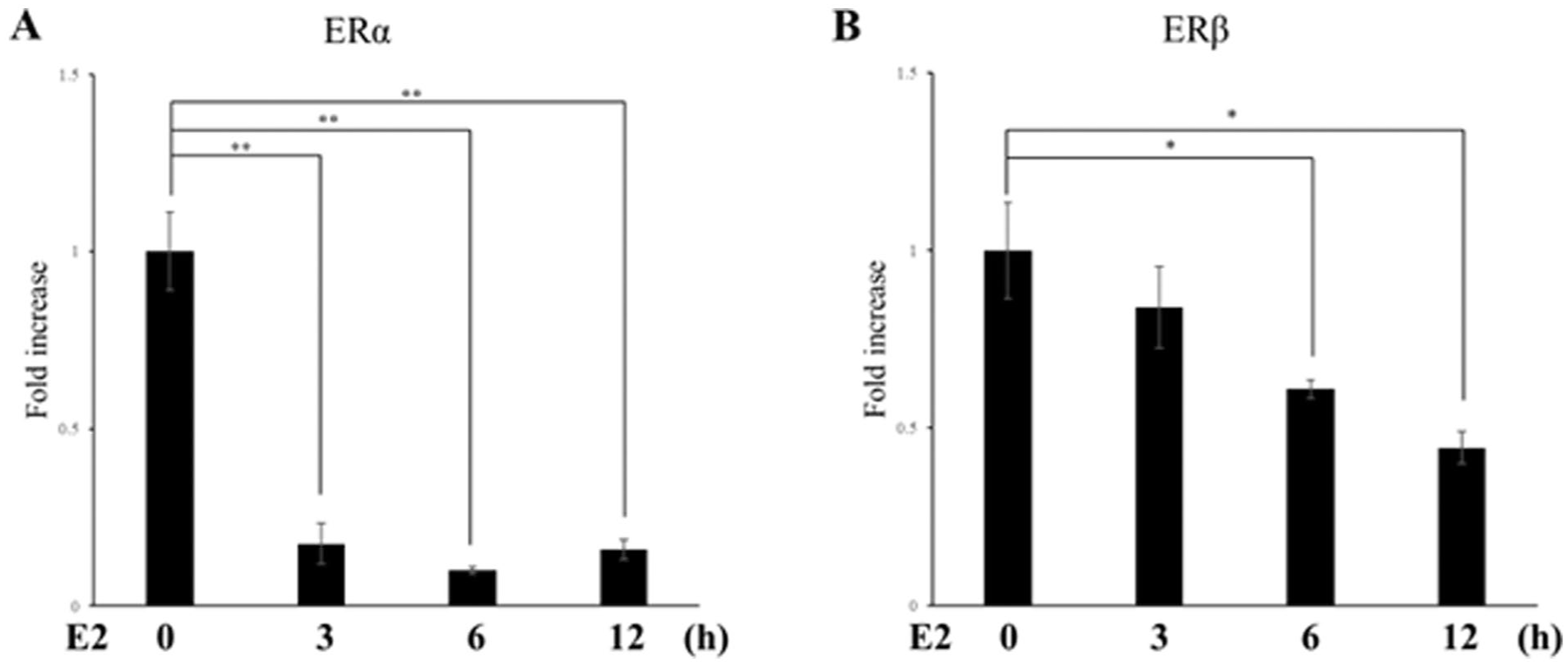
Regulation of ER Gene Expression in ATDC5 Cells Under Excessive Mechanical Stress
RT-qPCR analysis demonstrated the opposite regulatory patterns of ERα and ERβ under mechanical stress. CTS significantly suppressed ERα expression at all time points (P = 0.0134: 3 hours, P = 0.0023: 6 hours, P < 0.001: 12 hours,
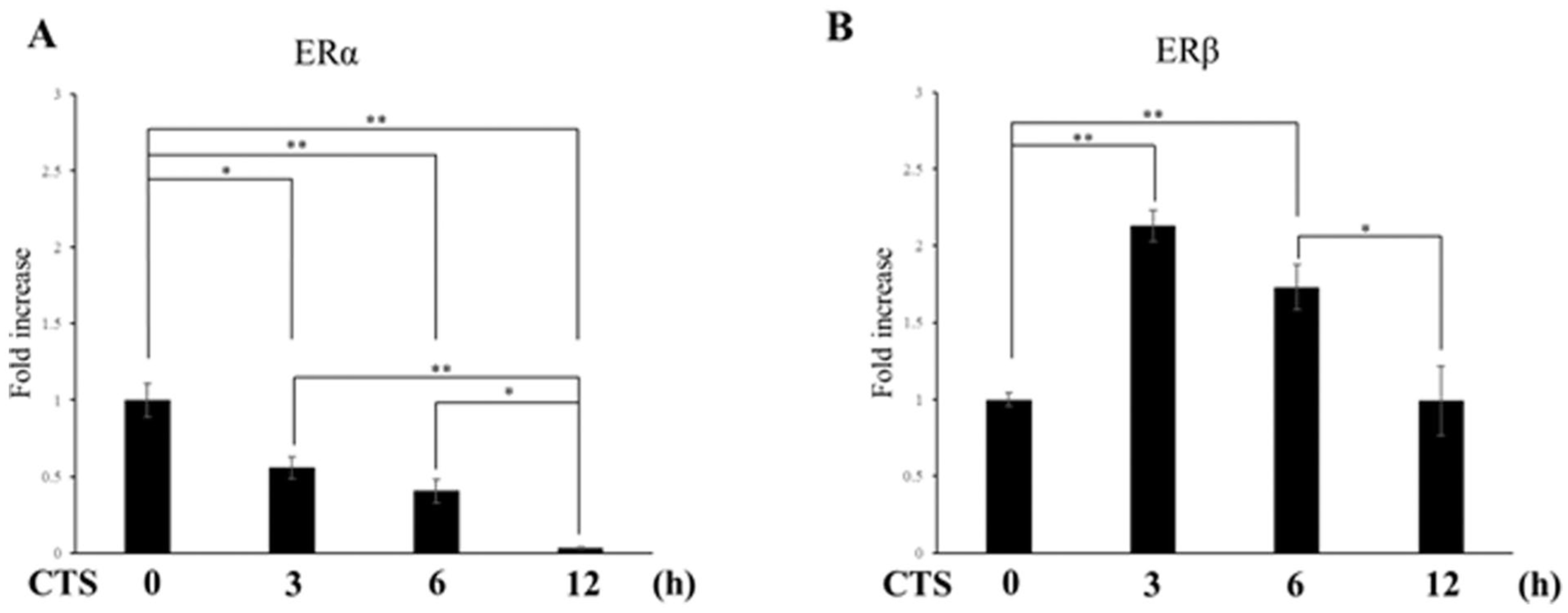
Effects of CTS on ER gene expression. Chondrocytes were exposed to high-intensity CTS for 3, 6, and 12 hours. At each time point, ERα and ERβ mRNA levels were quantified by real-time RT-PCR and normalized to S29. Data are expressed as mean ± SD from three independent experiments (n = 3). E2 = estradiol; CTS = cyclic tensile strain; ERα = estrogen receptor α; ERβ = estrogen receptor β. *P < 0.05, **P < 0.01.
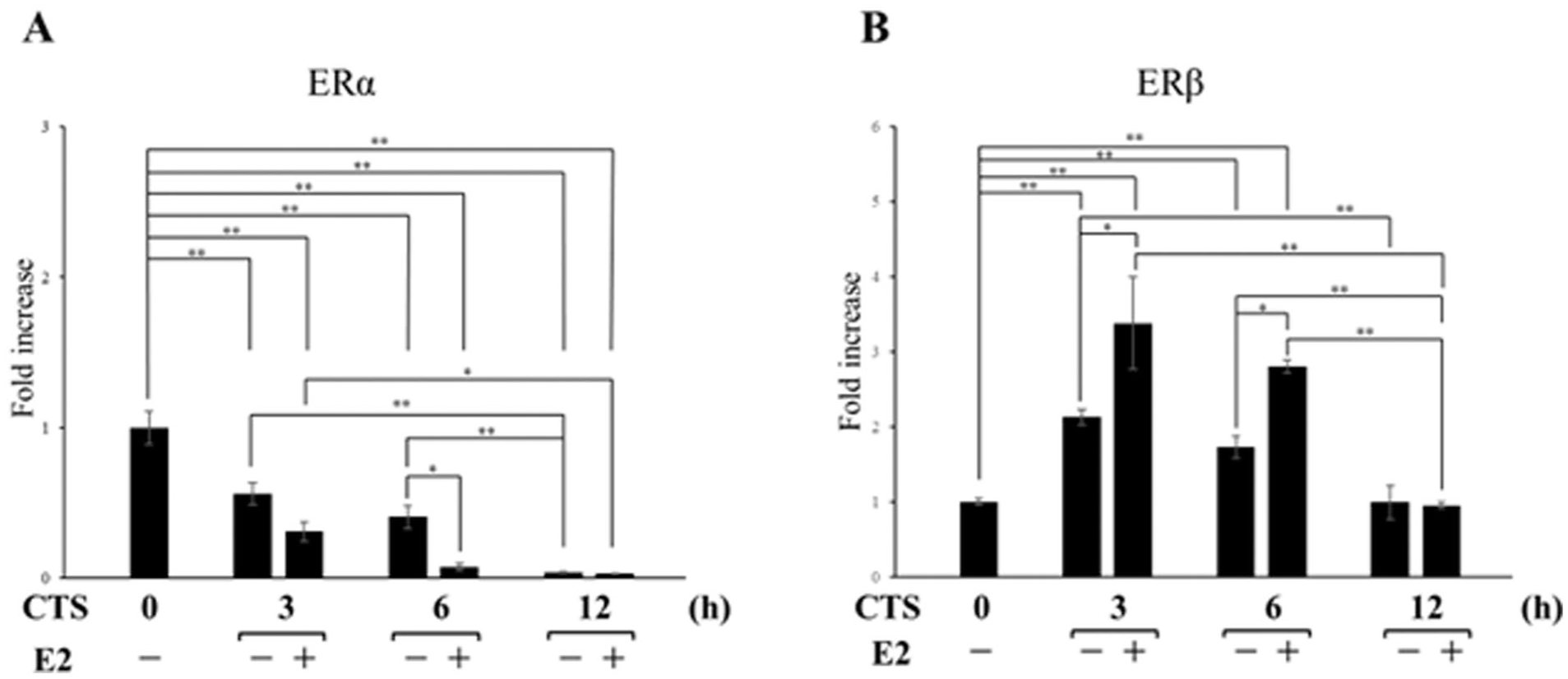
Effects of CTS+E2 on ER gene expression. Chondrocytes were exposed to high-intensity CTS (0.5 Hz, 10% isotropic strain)+E2(10⁻⁷M) for 3, 6, and 12 hours, and ERα and ERβ mRNA levels were quantified by real-time RT-PCR and normalized to S29. Data are expressed as mean ± SD from three independent experiments (n = 3). E2 = estradiol; CTS = cyclic tensile strain; ERα = estrogen receptor α; ERβ = estrogen receptor β. *P < 0.05, **P < 0.01.
Discussion
Osteoarthritis (OA) is more prevalent in women, particularly after menopause, implicating a potential role for estrogen in its pathogenesis. However, the precise mechanisms linking estrogen and OA remain to be elucidated. In this study, we demonstrated that estrogen exerts chondroprotective effects under excessive mechanical stress, which appears to be mediated, at least in part, by the modulation of ER expression.
In this study, estrogen treatment significantly suppressed the upregulation of cartilage degeneration factors induced by excessive mechanical stress in ATDC5 cells
First, we investigated the effect of estrogen treatment on ER expression in the absence of mechanical loading (i.e., CTS). In chondrocytes, estrogen treatment significantly downregulated ERα and ERβ expression
Next, we investigated the changes in ER expressions induced by CTS. The expression of ERα in chondrocytes was significantly downregulated at all time points under excessive mechanical stress
Thus far, we have discussed the independent effects of CTS loading and E2 treatment on chondrocytes. Here, we considered how ERs respond when these two conditions are combined. E2 treatment under CTS conditions reduced ERα gene expression at 6 hours compared with the CTS-only condition
In contrast, for ERβ, E2 treatment under CTS loading increased ERβ gene expression at 3 and 6 hours compared with the CTS-only condition
However, this study has several limitations. First, as it is an in vitro validation using the ATDC5 cell line, it does not fully replicate the diversity of chondrocytes in vivo nor the complex pathophysiology of OA.31,32 Second, the CTS protocol used in this in vitro study was designed to induce inflammatory responses in chondrocytes and does not replicate the cartilage-destructive conditions of osteoarthritis in vivo. These in vivo conditions were previously addressed in our ex vivo study using organ-cultured rat mandibular condyles. 33 Future studies are warranted to investigate ER behavior under these in vivo conditions. Furthermore, although we confirmed changes in ER expression under mechanical stress and its inhibitory effects on inflammation-related factors, our findings emphasize the protective role of ERβ. Nevertheless, the lack of loss-of-function experiments prevents direct proof of causality. To elucidate the physiological significance of ER signaling in response to mechanical stress and its therapeutic potential for OA, future studies using ERα and ERβ knockdown experiments and in vivo models are necessary.
In summary, our findings revealed that under excessive mechanical stress, estrogen supplementation led to distinct responses between ERα and ERβ. To the best of our knowledge, such differences in receptor reactivity have not been previously reported, and these insights have important implications for understanding the role of estrogen in various joint diseases.
Conclusions
This study demonstrated that estrogen exerts a chondroprotective effect under excessive mechanical stress, predominantly mediated by ERβ rather than ERα.
Footnotes
Acknowledgements
The authors thank all members of the Department of Orthodontics and Craniofacial Developmental Biology, Hiroshima University Graduate School of Biomedical and Health Sciences for their support of the present study.
Ethical Considerations
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The original contributions presented in the study are included in the article and its Supplementary Material. Further inquiries can be directed to the corresponding author.