
Abstract
Objective
Examination of intervertebral disc (IVD) regeneration in an ovine annular lesion model.
Hypothesis
Sulfation motifs are important functional determinants in glycosaminoglycans (GAGs). Previous studies have correlated 3-B-3(−) and 7-D-4 chondroitin sulfate (CS) motifs in tissues undergoing morphogenetic transition in development. We hypothesize that these motifs may also be expressed in degenerate IVDs and may represent a reparative response.
Design
Induction of disc degeneration by 5 mm or 6 × 20 mm lesions in the annulus fibrosus (AF) over 6 or 3 to 6 months postoperation (PO). Tissue sections were stained with toluidine blue–fast green, 3-B-3(−) and 7-D-4 CS-sulfation motifs were immunolocalized in 3-month PO 6 × 20 mm lesion IVDs. Sulfated glycosaminoglycan (GAG), 3-B-3(−), and 7-D-4 epitopes were quantitated by ELISIA (enzyme-linked immunosorbent inhibition assay) in extracts of AF (lesion site and contralateral half) and nucleus pulposus (NP) 0, 3, and 6 months PO.
Results
Collagenous overgrowth of lesions occurred in the outer AF. Chondroid metaplasia in ~20% of the 6 × 20 mm affected discs resulted in integration of an outgrowth of NP tissue with the inner AF lamellae preventing propagation of the lesion. 3-B-3(−) and 7-D-4 CS sulfation motifs were immunolocalized in this chondroid tissue. ELISIA quantified CS sulfation motifs demonstrating an increase 3 to 6 months PO in the AF lesion and a reduction in sulfated GAG not evident in the contralateral AF.
Conclusions
(1) Outer annular lesions underwent spontaneous repair. (2) Chondroid metaplasia of the inner 6 × 20 mm defect prevented its propagation suggesting an apparent reparative response.
Introduction
Low back pain (LBP) is a leading global musculoskeletal condition and a major public health concern with approximately 650 million people affected. 1 The global socioeconomic impact of LBP is increasing with changing trends in aging population demographics.1-3 United Nations World Ageing Data collected from 195 countries evaluated by the World Bank shows that the number of individuals aged ≥65 years will dramatically increase over the next 2 decades. 4 LBP is associated with degenerative disc disease (DDD) 5 and is a major global cause of disability 1 imposing a heavy burden on health care systems and significant direct and indirect socioeconomic impact 6 accounting for total annual health care expenditure of £11 billion in the United Kingdom in 2000,7,8], $9.17 billion in Australia in 2003, 9 and $50 to $90 billion annually in the United States.10-12 The World Health Organization (WHO) IRIS Low Back Pain Initiative stated that studies on IVD regeneration are a high priority research area, 13 LBP was made a national health care priority in Australia in 2009. 14 LBP treatment with opioid pain killing prescriptions in the United States is currently running at epidemic proportions. 15
Regeneration of the intervertebral disc (IVD) is limited by a low cell density and poor nutrient supply,16,17 there is a tremendous unmet need for the development of an effective treatment for intervertebral disc degeneration (IVDD) and associated LBP.18,19 The ovine model was developed to evaluate aspects of IVDD but was also useful in studies on disc regeneration and prospective treatments to either slow or prevent DDD or stimulate IVD regeneration.20-22 The ovine model of 199023,24 used a controlled 5-mm deep annular lesion, which produced IVDD over an 18-month period and was characterized using biochemical, 25 histochemical, Western blotting of aggrecan catabolism,26,27 and biomechanical evaluation.28-32 A steady decline in IVD proteoglycan/catabolism of aggrecan were major features,26,33 upregulation in decorin and biglycan biosynthesis, fragmentation of fibromodulin and biglycan was also observed27,34 with roles in collagen fibrillogenesis, matrix assembly, and signal transduction important in IVD regeneration.35-41 An influx of transforming growth factor–β (TGF-β) expressing cells 42 and in-growth of blood vessels 43 promoted lesion repair.43-45 A focal reduction in vertebral subchondral bone density, endplate vascularity, 46 remodeling of vertebral bone, 47 facet joint degeneration were also observed 48 and disruption of normal annular laminate structure, attachment to vertebral bone and internal nucleus pulposus (NP) organization.31,49,50 A modification of this model was developed in 2012 using a 6 × 20 mm annular lesion, 51 biochemical, compositional, gene profiling, and histopathological scoring confirmed its degenerative features. 22 Exogenously administered mesenchymal stem cells (MSCs) promoted repair of the annular lesion. Hyaluronan (HA) oligosaccharides generated by depolymerization of HA during IVDD promoted extracellular matrix (ECM) remodeling through matrix metalloproteinases (MMPs) and spontaneous repair of the outer annular defect. Spinal manipulation studies demonstrated annular lesions perturbed normal spinal biomechanics, neurophysiology, stabilization of vertebral lumbar motion segments, and dynamic dorsoventral lumbar spinal stiffness.52-55 Inflammatory cytokine-mediated multifidus remodeling may indirectly contribute to the generation of LBP.56-59
The aim of the present study was to use the ovine model 51 to examine changes in chondroitin sulfate (CS) sulfation associated with IVD repair. Monoclonal antibodies (MAbs) to the CS sulfation motifs, 3-B-3(−) and 7-D-4 have immunolocalized these epitopes in tissue development and pathological remodeling 60 in the bursae of Fabricius, 61 skin, 62 neural tissues, 63 articular cartilage,64-67 embryonic chick bone marrow, and epiphyseal cartilage. 63 CS-epitope mapping of proteoglycans in health and disease60,68-71 showed 3-B-3(−) and 7-D-4 associated with progenitor/stem cell proteoglycans in fetal rat IVD, human knee and elbow, and discrete progenitor/stem cell niches. Cell clusters displayed 7-D-4 and 3-B-3(−) in fibrillated adult OA articular cartilage and in annular remodeling.72,73 A positive correlation in 3-B-3(−) and 7-D-4 levels in areas of ECM remodeling is evidence of how the glycosaminoglycan (GAG) chain proteoglycan structure is dynamically modulated during pathological remodeling/repair, recapitulating changes seen in foetal skeletal development.60,74
Materials and Methods
Materials
All material and supplier details were as indicated earlier. 74
Animal Welfare and Ethics
A total of fifty 2- to 3-year-old merino wethers (castrated males) were purchased from local sale yards and held in open paddocks till required. All animal welfare and ethics for this work were approved by the University of Sydney Animal Care and Ethics Committee under ethics approval A45/6-2011/3/5544.
Antibodies
MAb 3-B-3 and 7-D-4 were kind gifts of Prof. B. Caterson, School of Biosciences, University of Cardiff, UK. Chondroitinase ABC-digested rat chondrosarcoma proteoglycan was used as antigen to raise MAb 3-B-3. 75 Antibody 7-D-4 was produced from 17-day embryonic chick bone marrow proteoglycan as antigen. 76
Methods
Animal Model Procedures
A total of 48 animals were used in this study, and these were separated into 3 groups. Group A (n = 12) was divided into 6 animals that received 5-mm annular lesions at L1-L2, L3-L4, L5-L6 and 6 nonoperated control (NOC) animals. These were sacrificed 6 months postoperation (PO). Group B (n = 24) were divided unto a lesion group (n = 12), which received 6 × 20 mm lesions at L1-L2, L3-L4, L5-L6, and NOC sheep (n = 12), 6 animals were sacrificed at 3 months PO and the remainder at 6 months PO in each group. Group C consisted of 18 animals, which received 6 × 20-mm lesions at L1-L2, L3-L4, L5-L6, 6 animals were sacrificed at 0, 3, and 6 months PO in this group, an additional 6 NOC animals were sacrificed at 6 months PO. IVDs from group A and B sheep were examined histologically. Group C IVDs were dissected into annulus fibrosus zone 1 (AF1), AF2, and NP and the tissue extracts analyzed by ELISIA (enzyme-linked immunosorbent assay) and their sulfated GAG contents determined. Thus, a total of 144 IVDs were analyzed in this study. These time points were selected based on earlier studies which demonstrated at the selected time points early stages of disc degeneration was evident. Six animals per time point containing 18 treated IVDs previously provided statistically significant data.
Zonal Dissection of IVDs and Extraction of Proteoglycans for Compositional and ELISIA Analysis of 3-B-3(−) and 7-D-4 CS Sulfation Motifs
These IVDs were horizontally bisected and photographed, then zonally dissected into the AF half containing the lesion site (AF zone 1), unaffected contralateral AF zone 2 and the central NP tissue zone. AF zone 1, AF zone 2, and NP were weighed, finely diced, and extracted with 4 M GuHCl containing protease inhibitors (10 mL/g tissue) for 48 hours and the extract clarified by centrifugation then dialyzed against MilliQ distilled water to remove GuHCl and freeze dried, redissolved in MilliQ distilled water and their sulfated GAG contents determined by reaction with the metachromatic dye 1, 9-dimethylmethylene blue. Their 3-B-3(−) and 7-D-4 CS sulfation epitopes were quantitated by ELISIA.
Native Composite Agarose–Polyacrylamide Gel Electrophoresis (CAPAGE) and Immunoblotting of IVD Extracts
Aliquots of IVD or shark cartilage extracts (5µg sulfated GAG/10 µL per well) were electrophoresed in 3-mm thick CAPAGE slab gels 77 and the gel stained directly with toluidine blue to visualize proteoglycans or electroblotted to nitrocellulose membranes by semidry blotting using 1/50 dilutions of 7-D-4 or 3-B-3 hybridoma conditioned media samples in 50 mM Tris-HCl, 0.1 M NaCl, pH 7.2 at 4°C, primary antibodies were localized using alkaline phosphatase conjugated goat anti-mouse IgM secondary antibodies (1/1000 dilution, 1 hour at room temp) and visualized using NBT/BCIP (nitro blue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate) substrates in 0.1 M Tris-HCl pH 9.2.
Histological Processing of IVDs
IVD spinal motion segments were trimmed of excess bone leaving a portion of the inferior and superior vertebral body attached to the IVD and fixed en bloc for 48 hours in 10% neutral buffered formalin, then decalcified in 10% (v/v) formic acid 5% (v/v) neutral buffered formalin for 10 to 12 days using frequent changes of decalcification solution and constant agitation till a needle could be inserted into the vertebral body with moderate pressure. The fixed decalcified IVD specimens were then cut into 5-mm slabs encompassing the lesion site and dehydrated in graded ethanol washes and xylene, and embedded in paraffin wax. Vertical microtome sections through the IVD and superior and inferior vertebral bodies were subsequently cut at 4 µm thickness and attached to SuperFrost Plus glass microscope slides (Menzel-Glaser, Germany) by baking at 55°C for 30 minutes in an oven and stored at room temperature overnight before use.
Immunolocalization of the 3-B-3(−) and 7-D-4 CS Sulfation Motifs
Vertical tissue sections through IVD lesion sites and inferior and superior vertebral bodies were deparaffinized in xylene and in a sequential series of ethanol solutions to water. Antigen retrieval was carried out with proteinase K digestion (20 µg/mL in Tris-EDTA buffer) for 10 minutes at 37°C and endogenous peroxidase activity blocked 3% H2O2 for 5 minutes. The slides were then washed with 50 mM Tris 150 mM NaCl, 0.05% (w/v) Tween-20 pH 7.2 (TBST) and blocked in 1% bovine serum albumin (BSA) in TBST. The slides were then incubated with primary antibody hybridoma conditioned media, clone 3-B-3 (1:500) or clone 7-D-4 (1:1000) diluted in 1% BSA in TBST for 16 hours at 4°C, washed twice with TBST before incubation with biotinylated anti-mouse IgG secondary antibodies (1:500; GE Healthcare) for 1 hour at room temperature. The slides were then washed twice with TBST, and streptavidin–horse radish peroxidase (HRP) (1:250; GE Healthcare) added for 1 hour at room temperature. Following further washes in TBST, NovaRED chromogen (Vector Laboratories; Burlingame, CA) was used for color development in HRP development buffer. The slides were then counterstained with hematoxylin (Gill’s #3, Vector Laboratories) for 6 seconds and rinsed in deionized water, then dehydrated through a graded ethanol series, mounted in Eukitt and imaged using a Mirax Digital Slide Scanner (Zeiss). Negative control slides were similarly processed using irrelevant isotype matched mouse Ig whole antibodies (2 μg/mL; Invitrogen, Carlsbad, CA) or with omission of primary antibodies. Both sets of controls were negative.
Toluidine Blue Staining
Glycosaminoglycans were stained in IVD sections for 10 minutes with 0.04% (w/v) toluidine blue in 0.1 M sodium acetate buffer, pH 4.0, followed by a 2-minute counterstain in 0.1% (w/v) fast green FCF.
Hematoxylin and Eosin Staining
IVD sections were stained in Mayers hematoxylin (5 minutes) to assess cellular morphology, rinsed in tap water blued in Scotts Blueing solution (1 minute) and counterstained in 0.0001% eosin (5 minutes), dehydrated in 95% (v/v) ethanol then absolute ethanol, cleared in xylene and mounted.
Sulfated GAG Analysis of IVD Tissue Extracts
The proteoglycan content of tissue extracts and proteoglycan standards were measured by their sulphated GAG contents by reaction with the metachromatic dye 1,9-dimethyl methylene blue using bovine tracheal CS as standard. 78
Isolation and Characterization of 7-D-4 and 3-B-3(−) Proteoglycan Standards
7-D-4 Standard
Shark cartilage purchased from a local Chinese traditional medicinal apothecary was rehydrated overnight in isotonic saline and finely diced and extracted in 10 volumes buffered 4 M GuHCl pH 5.8 + protease inhibitors with end-over-end stirring for 48 hours at 4°C. The tissue extract was clarified by centrifugation and CsCl added to a starting density of 1.4 g/mL and dissociative isopycnic density gradient ultracentrifugation conducted at 200,000 × g in a Sorval OTD75B vertical rotor for 48 hours. The tubes were manually fractionated into 9 equal fractions. Known volumes of fractions were weighed to construct a density gradient profile and aliquots were reacted with 1,9-dimethyl methylene blue to determine proteoglycan distribution. The densest fraction was used as a source of purified 7-D-4 standard.
3-B-3(−) Standard
Lumbar IVDs from 3 stillborn lambs were finely diced and extracted as outlined for preparation of the 7-D-4 shark cartilage standard and subjected to dissociative CsCl density gradient ultracentrifugation as outlined for the 7-D-4 standard except the D1 fraction was subjected to a second ultracentrifugation and the D1D1 fraction used as 3-B-3(−) standard.
Quantitation of the 7-D-4 and 3-B-3(−) Sulfation Motifs by ELISIA
Binding curves were made for the 7-D-4 and 3-B-3(−) standards to the Maxisorp microtiter plates to determine optimal standard proteoglycan coating concentrations for the ELISIA. The standards were redissolved in 50 mM Tris-HCl pH 7.2 containing 100 mM NaCl (TBS) containing 2% w/v BSA at 100 µg/mL and a series of serial doubling dilutions were prepared. Sextuplet replicates (50 µL) were applied to 96-well microtiter plates and binding undertaken at 4°C overnight, the well contents were then flicked out and the wells rinsed three times with TBS. Binding curves of proteoglycan standards were determined using 1,9-dimethyl methylene blue. 78 Plates were subsequently coated using a 50 µg/mL standard solution. A series of serial doubling dilutions of disc proteoglycan samples with 1/50 dilutions of MAb 7-D-4 and 3-B-3 hybridoma conditioned media samples in TBS were prepared and thoroughly mixed. After 1 hour at 37°C, aliquots (50 µL) of the serial dilutions were added to the coated plates and the plates incubated for 3 hours at 37°C. The plate contents were then flicked out and 200 µL of a 2% BSA in TBS blocking solution added. After 1 hour at 37°C the wells were emptied and washed 3 times with TBS then alkaline phosphatase conjugated anti mouse IgM secondary antibody was added for 1 hour at 37°C. The wells were rinsed again in TBS and ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) in development buffer (200 µL) was added. Color development was allowed to proceed for up to 1 hour at 37°C. The plates were read at 450 nm using a plate reader. ELISIAs were determined using 6 replicates and mean ± SD data used to plot ELISIA curves. 7-D-4 and 3-B-3(−) epitope levels were determined from the 50% inhibition values. 79
Statistical Methods
Spatiotemporal distributions of sulphated GAG and 3-B-3(−) and 7-D-4 CS sulfation motif ELISIA data in nonoperated control and lesion affected IVD tissue zones were compared using analysis of variance. Nonnormally distributed data were analyzed using Kruskal-Wallis 1-way analysis of variance. Significant differences in data sets were evaluated using the Mann-Whitney U test using GraphPad Prism software, the alpha level was set at 0.05.
Results
Frozen spinal segments were sectioned through the annular lesion site using a sledge microtome and the cut surfaces photographed ( Fig. 1 ). This demonstrated thickening of the outer annular lamella and macroscopic repair while the surgical incision in the inner AF defect was still clearly visible ( Fig. 1a ). Macroscopic views of bisected IVDs that had received 6 × 20 mm lesions showed the lesion confined to the outer third of the AF 3 months postoperatively ( Fig. 2a ) while at 6 months PO, blood vessels had penetrated deeper into the IVD and some discoloration of the NP was also evident ( Fig. 2b ). Localized thickening of annular lamellae spanning the original defect site was also a prominent feature of the 6-month PO defect IVDs ( Fig. 2b ). Comparison with age-matched NOC IVDs showed clear differences between the macroscopic appearances of NOC and 3- and 6-month lesion IVDs ( Fig. 2a-c ). Discs from the group C sheep were dissected into AF1, AF2, and NP tissue zones as shown ( Fig. 2d ) and proteoglycans were extracted from these tissue zones for compositional and ELISIA assays. Toluidine blue–fast green histology of NOC IVDs from group A and B sheep showed a widespread and uniform GAG distribution through the IVD except in the outer AF, which had markedly lower GAG levels ( Fig. 2e ). Comparison of the NOC IVD with a 6 × 20 mm lesion IVD showed a decreased disc height consistent with its lower GAG levels stained with toluidine blue, focal reduction in GAG levels in the outer AF lesion zone and propagation of the annular lesion through the IVD, clear evidence of the disc degeneration previously demonstrated in earlier studies using this type of annular lesion ( Fig. 2f ).
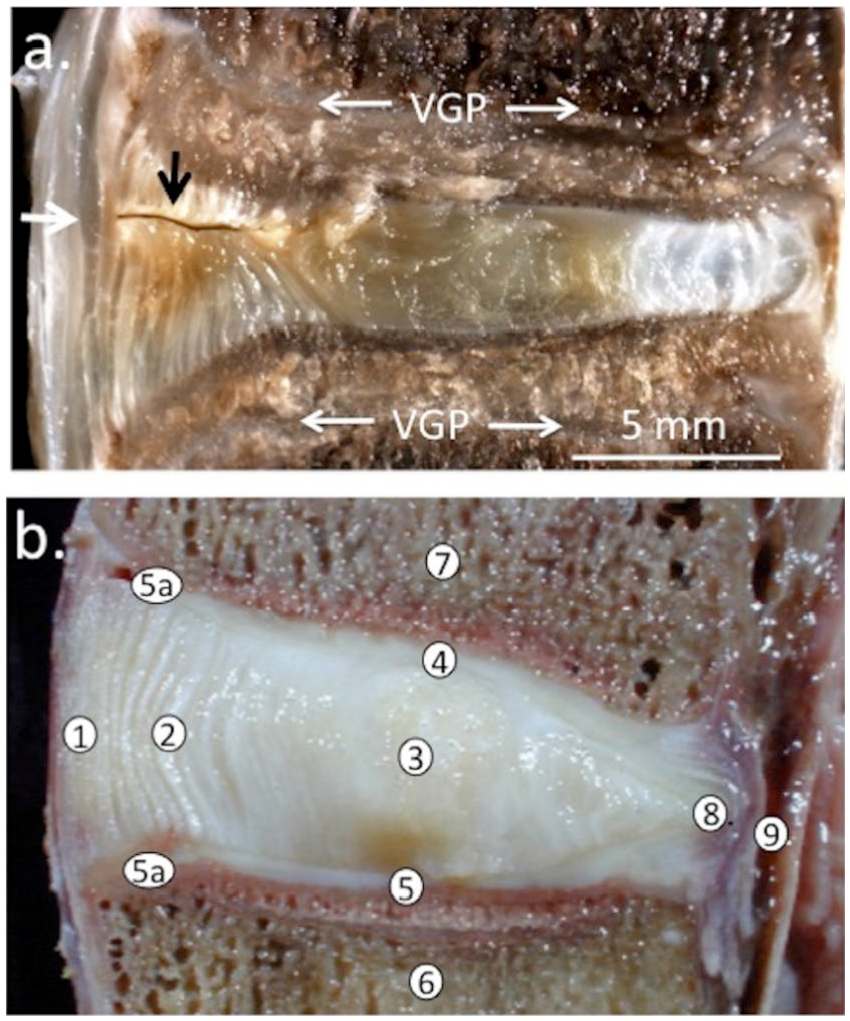
Depiction of a 5-mm deep controlled outer annular lesion in an ovine lumbar intervertebral disc (IVD) showing spontaneous repair in the outer annulus fibrosus (AF; small black arrow) and thickened annular lamellae 6 months after making the lesion, inner regions of the lesion remain unhealed. Macroscopic image of a nonfixed vertical sagittal section depicting the ovine IVD and adjacent superior and inferior vertebral bodies (
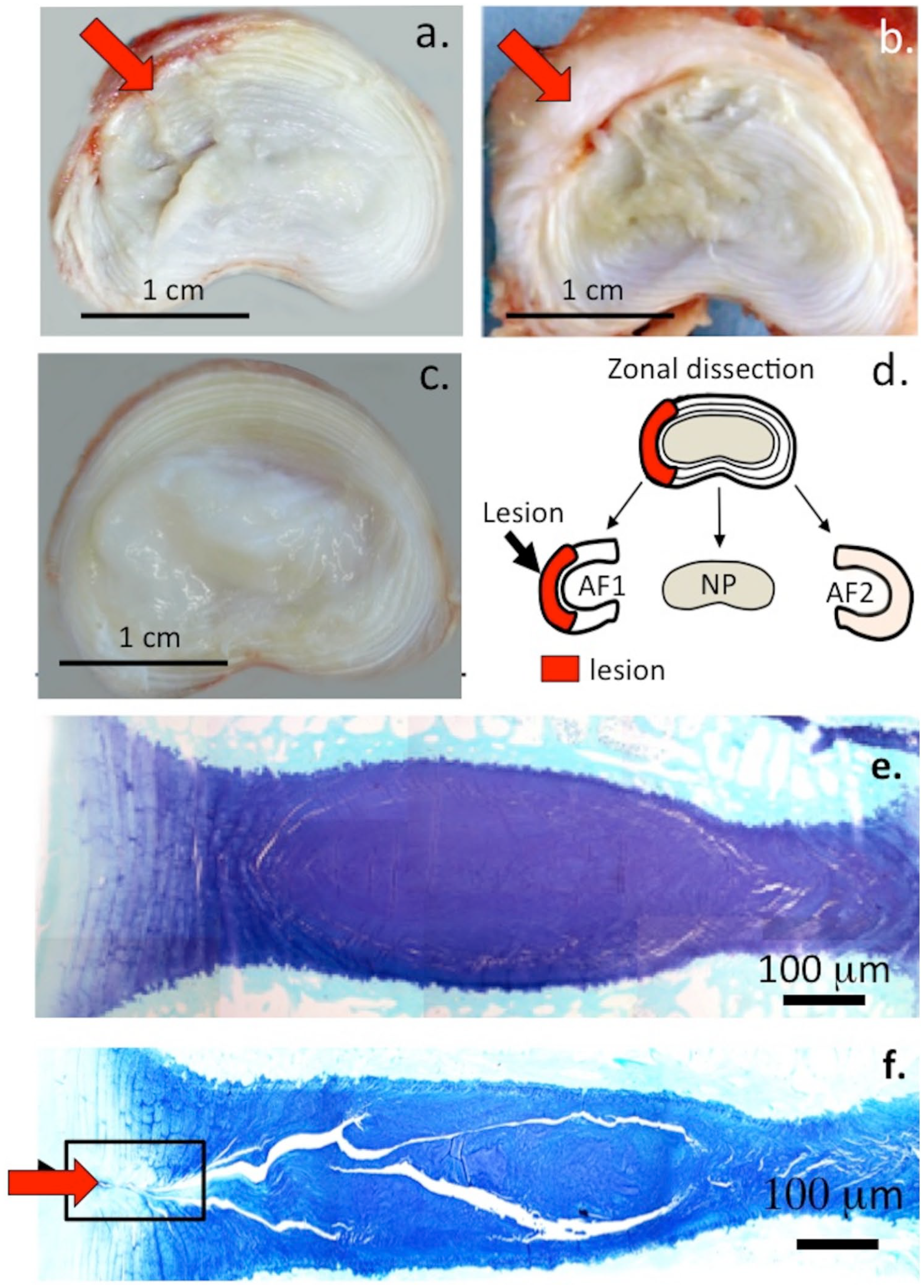
Morphological appearance of horizontally bisected 3- and 6-month postoperation (PO) 6 × 20 mm lesion affected (
Higher magnification views of an area equivalent to the boxed area in Figure 2f showed small groups of cells associated with the 5-mm lesion depicted in Figure 1 and focal loss of GAG ( Fig. 3a ). Small clones of cells were also evident in the boxed region of Figure 2f in the 6 × 20 mm lesion IVDs as well as a reduction in GAG staining ( Fig. 3b ). The cellular morphology of the inner ( Fig. 4a ) and outer AF ( Fig. 4b ) of NOC IVDs did not resemble that of the lesion IVDs ( Fig. 5a and b ) although occasional cell doublets were seen ( Fig. 4a ) but no evidence of small groups of cells.
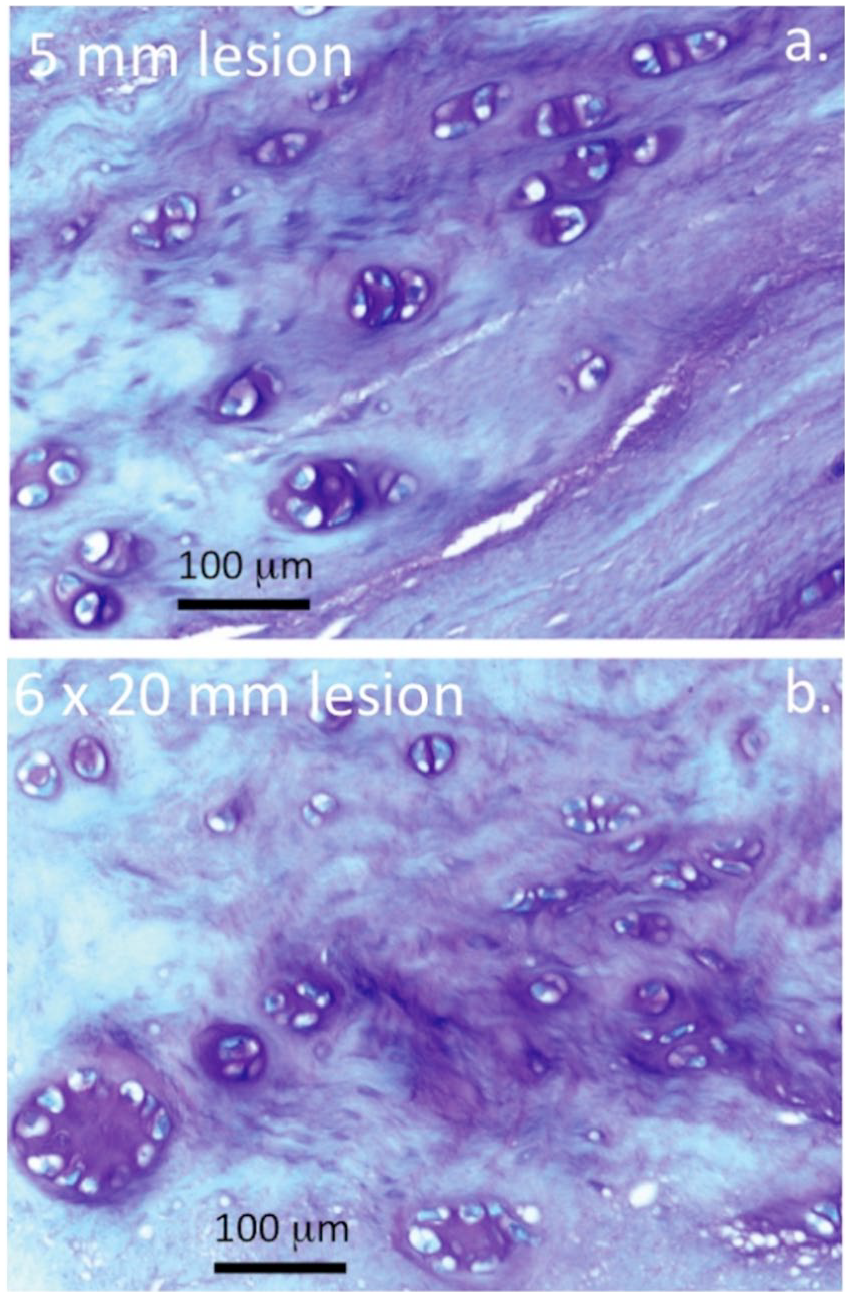
Toluidine blue histology of intervertebral disc (IVD) tissue sections depicting cell clustering and proteoglycan localization in ovine IVDs subjected to a 5-mm anterolateral annular lesion with degeneration induced for 6 months (
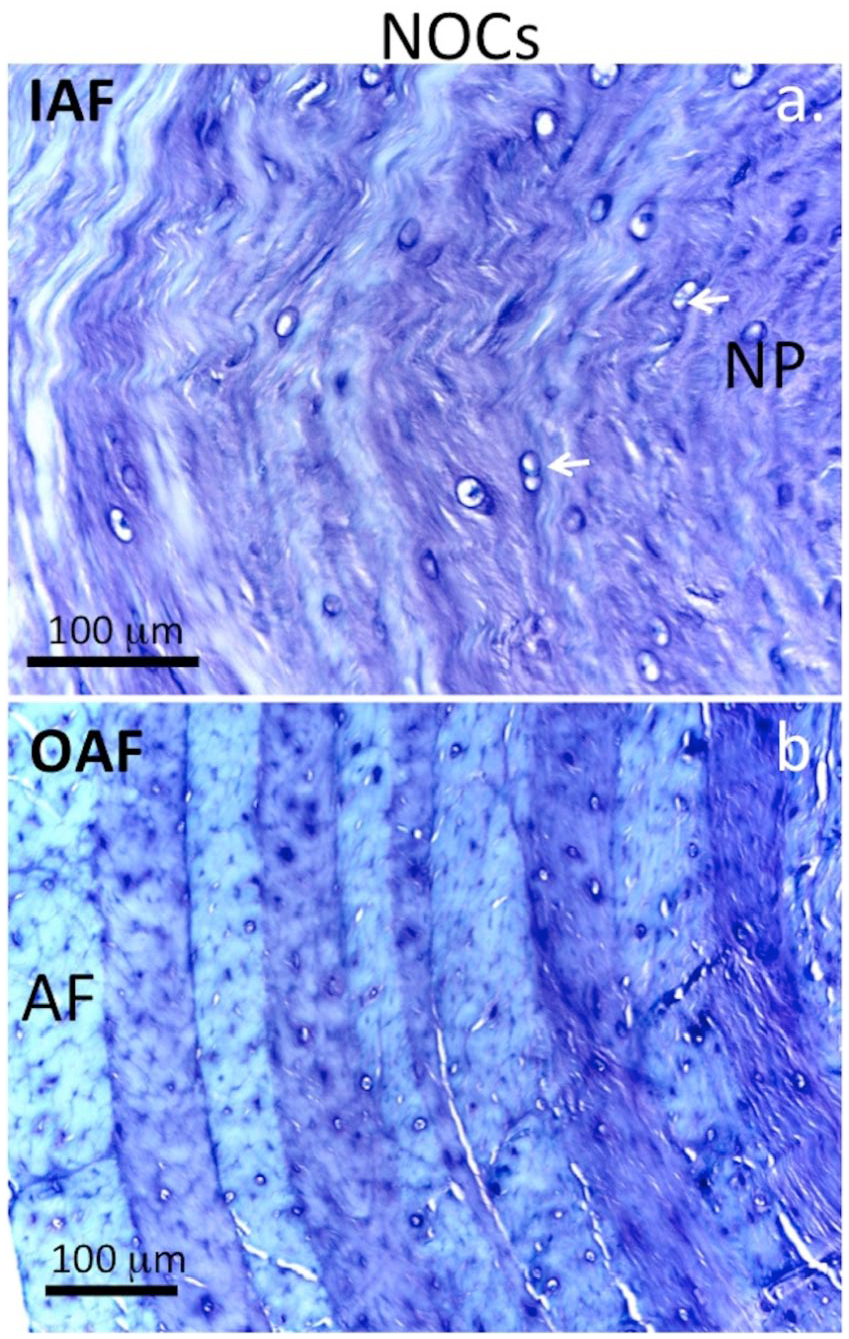
Medium power magnification images depicting the glycosaminoglycan (GAG) distribution and cellular morphology in the inner annulus fibrosus (AF) (
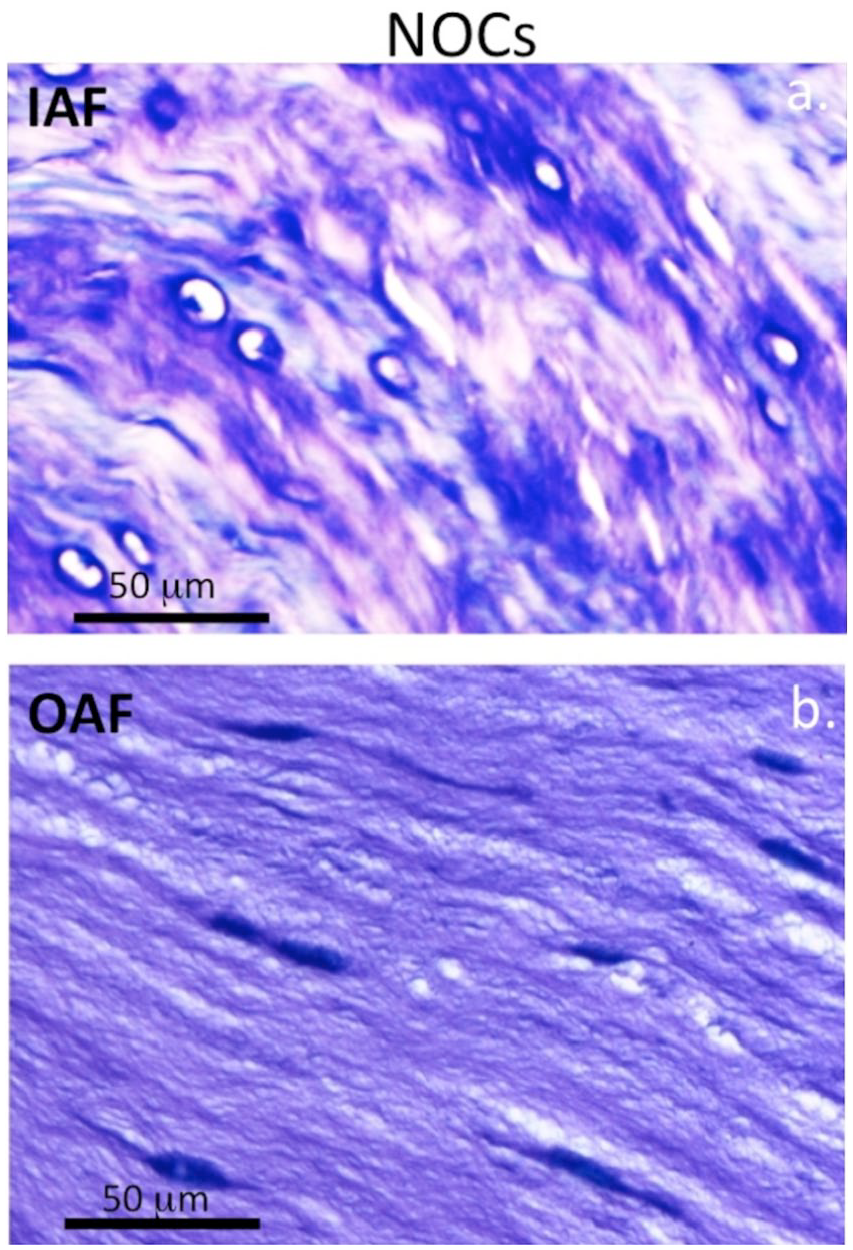
High power magnification images depicting the glycosaminoglycan (GAG) distribution and cellular morphology in the inner annulus fibrosus (AF) (
The 7-D-4 and 3-B-3(−) CS sulfation motifs were abundantly expressed in the NP of NOC IVDs but sparsely in the AF ( Figs. 6a and 7a ). In 6 × 20 mm lesion IVDs, an outgrowth of the NP along the line of the annular lesion contained abundant levels of these CS motifs ( Figs. 6c and 7c ). Higher magnification examination of these NP outgrowth tissues identified small groups of chondroid cells containing well-defined pericellular matrices rich in the 7-D-4 and 3-B-3(−) epitopes ( Figs. 6d , e and 7d , e ). These cells were contained in distinct nests in a matrix rich in the 7-D-4 and 3-B-3(−) epitopes ( Figs. 6e and 7e ). These chondroid cells were considerably larger than the AF or NP cells and of a dissimilar morphology and were only observed in the 6 × 20 mm lesion IVDs and not the 5-mm lesion IVDs. Examination of the 6 × 20-mm lesion IVD tissue sections stained with toluidine blue–fast green showed 11/36 IVDs displayed these NP outgrowths rich in GAG ( Fig. 8a-i ). These were prominent 3 months after making the annular lesion ( Fig. 8d-g ) but by 6 months PO, they had resorbed to a significant extent ( Fig 8a-c ). Higher power examination of this chondroid tissue demonstrated its integration with the lamellar layers of the inner AF apparently representing a repair response to the annular lesion ( Fig. 8i ). In disc specimens that displayed chondroid metaplasia, the annular lesion failed to propagate further into the IVD.
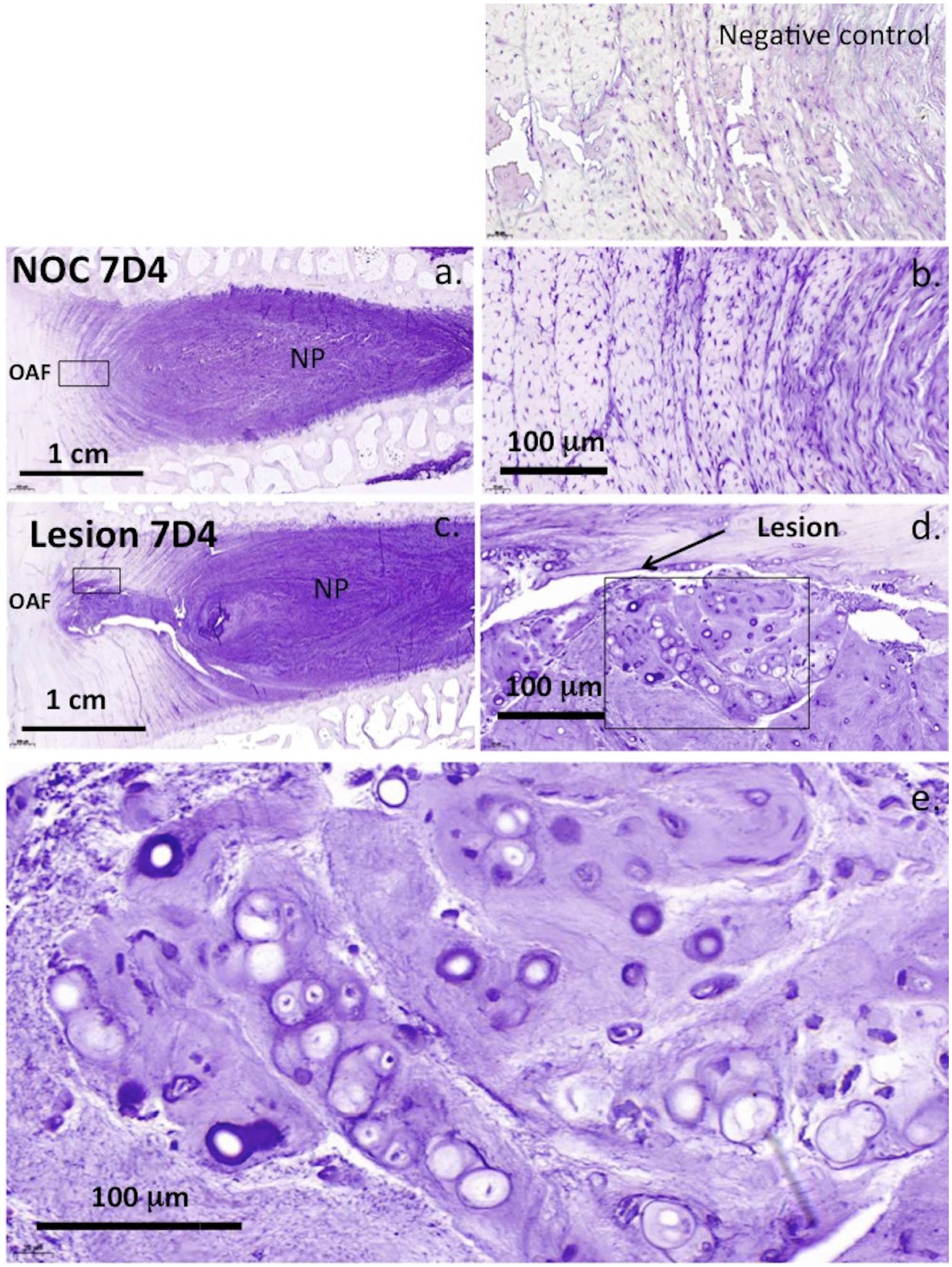
Immunolocalization of the 7-D-4 CS sulfation motif throughout the nucleus pulposus (NP) and inner annulus fibrosus (AF) to a lesser extent (
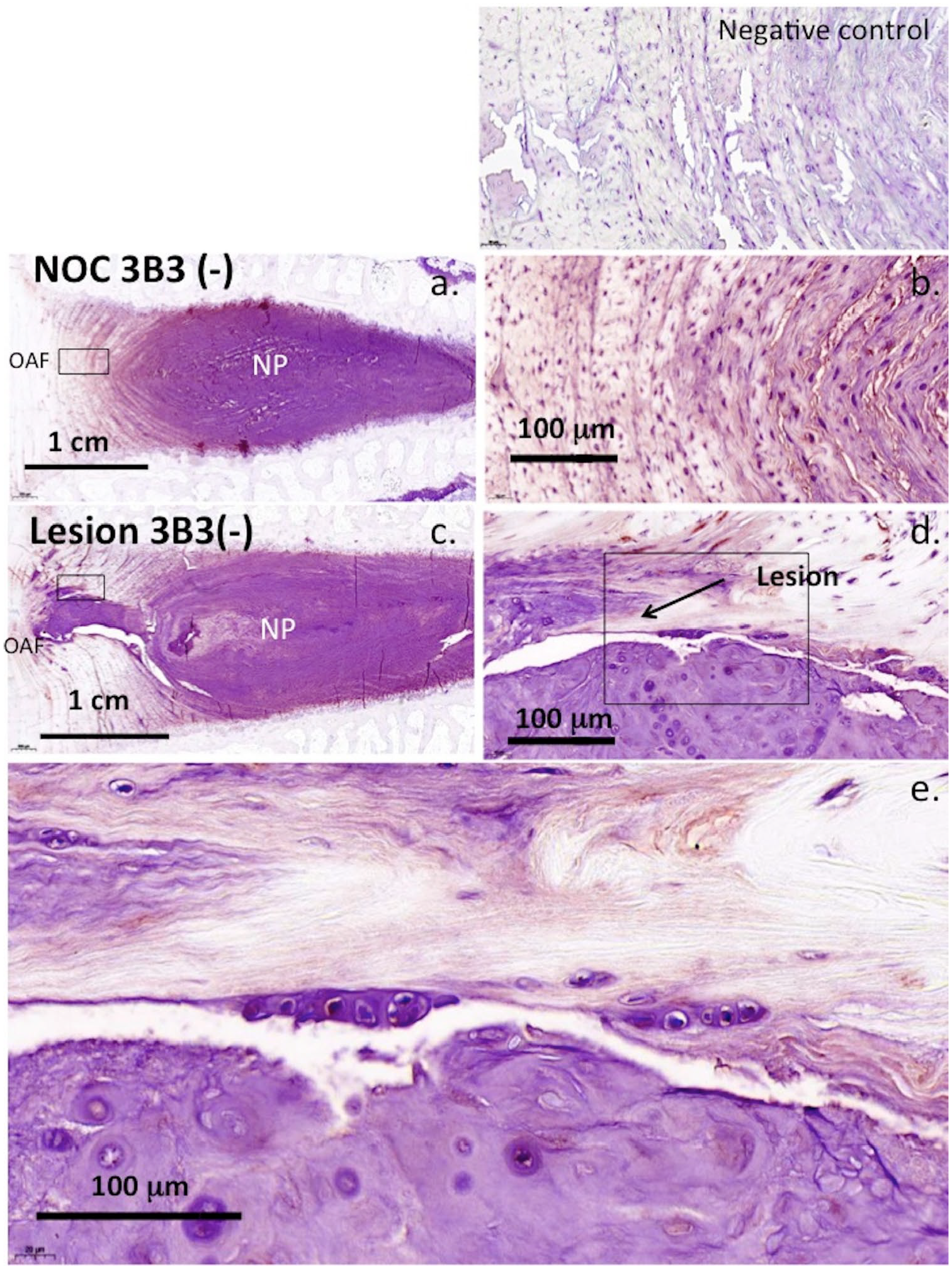
Strong expression of 3-B-3(−) CS sulfation epitope throughout the nucleus pulposus (NP) and inner annulus fibrosus (AF) to a lesser extent (
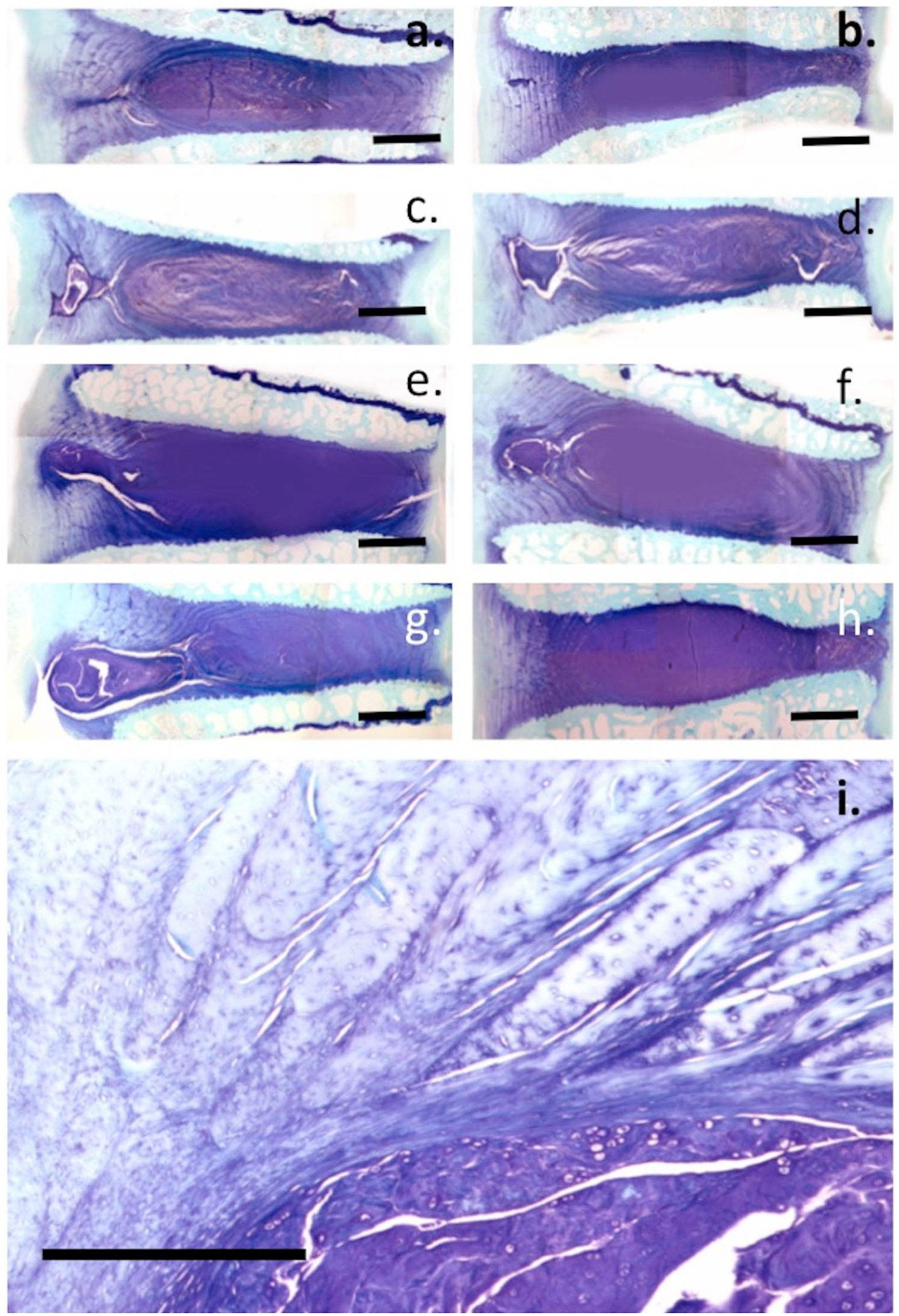
Toluidine blue–fast green stained tissue sections through annular repair regions depicting cartilaginous tissue that had been laid down in the lesion site through chondroid metaplasia. The images show examples of different stages of the resorption of the chondroid tissue mass 6 months postinduction of disc degeneration (
To further investigate the 7-D-4 and 3-B-3(−) epitopes in the ovine IVD tissues, ELISIAs were developed using proteoglycan standards isolated from shark cartilage rich in the 7-D-4 epitope and newborn ovine IVD enriched in the 3-B-3(−) epitope and the purified standards were characterized by CAPAGE, which is a native electrophoretic system that resolves proteoglycans on the basis of charge:mass ratio ( Fig. 9a ). Dissociative CsCl density gradient isopycnic ultracentrifugation fractionated proteoglycan species into high buoyant density (aggrecan, ρ > 1.55 g/mL) and low buoyant density proteoglycans (decorin/biglycan, ρ = 1.3-1.4 g/mL) identified on Immunoblots using the 7-D-4 and 3-B-3 antibodies ( Fig. 9 b-d ).
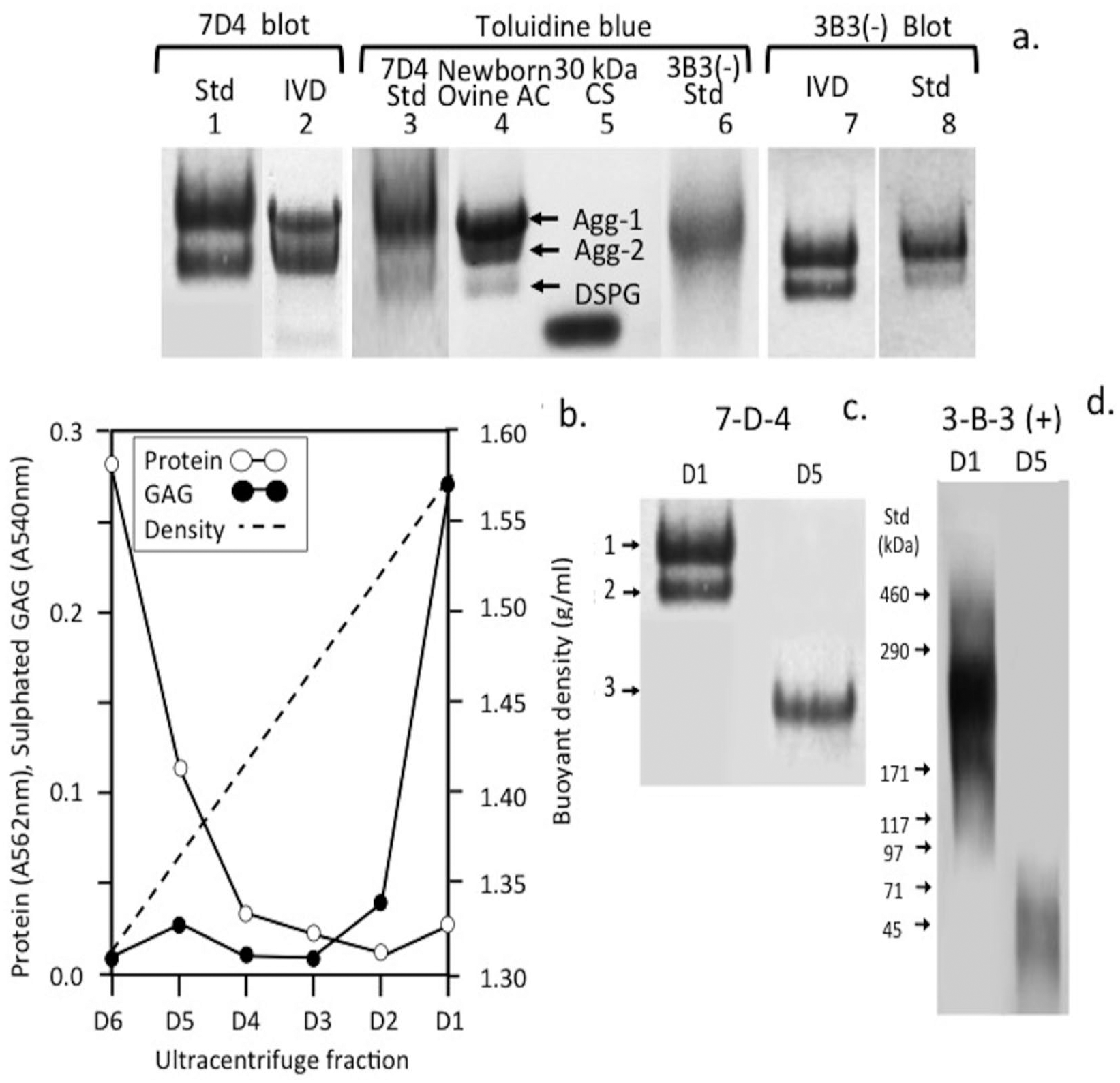
Composite agarose–polyacrylamide gel electrophoresis and identification of the 7-D-4 and 3-B-3(−) standards prepared from shark cartilage and newborn ovine intervertebral disc by immunoblotting using MAb 7-D-4 and 3-B-3 (
The purified 7-D-4 shark aggrecan and ovine IVD 3-B-3(−) aggrecan standards were used to develop ELISIAs to quantitate relative levels of the 7-D-4 and 3-B-3(−) epitopes in tissue extracts from the 6 × 20 mm lesion site (AF1 zone), the contralateral AF (AF2 zone) and NP from IVDs in which the annular lesion had been induced for 0, 3 and 6 months to induce disc degeneration ( Fig. 10 ). Binding curves were initially established for the proteoglycan standards to optimize coating of the ELISA 96-well microtiter plates ( Fig. 10a ) and standard inhibition curves established for quantitation of unknown samples ( Fig. 10b ). Sulfated GAG levels in the disc zonal extracts and NOC IVDs were also determined ( Fig. 10c and f ). Levels of the CS sulfation motif epitopes 7-D-4 ( Fig. 10d ) and 3-B-3(−) ( Fig. 10e ) were also quantitated by ELISIA compared with the levels in the AF1, AF2, and NP zones of NOC IVDs ( Fig. 10g and h ).
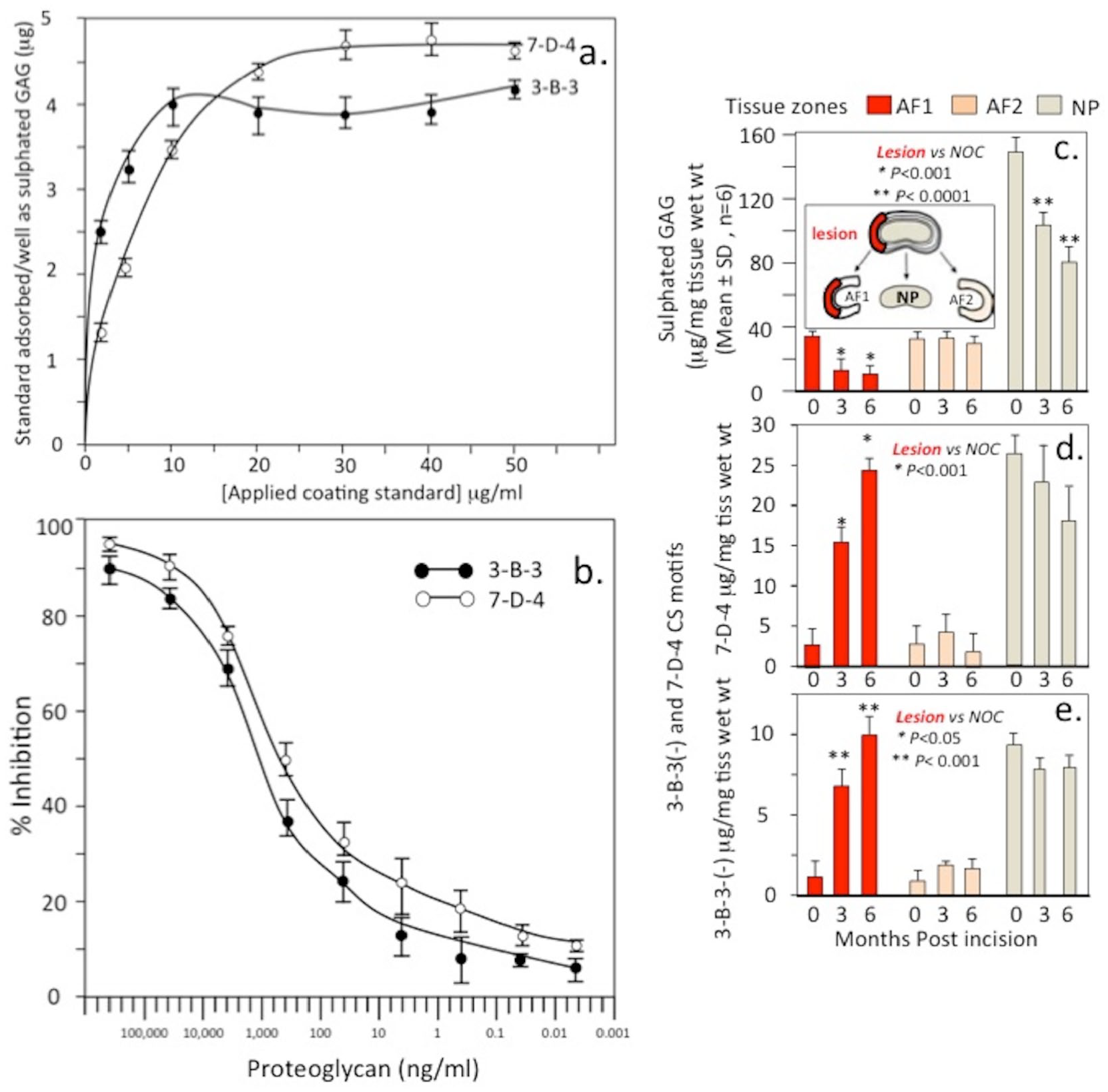
Measurement of 3-B-3(-) and 7-D-4 chondroitin sulfate (CS) sulfation epitopes by enzyme-linked immunosorbent inhibition assay (ELISIA) using shark cartilage 7-D-4 and 3-B-3(−) proteoglycan standards from newborn ovine intervertebral disc (IVD). ELISA standard binding curves for the proteoglycan standards (
Discussion
The ovine annular lesion model was originally developed to model human disc degeneration23,25 and along with the canine IVD80-83 is the best animal model for such studies providing important insights into IVD pathobiology. 20 After conducting numerous studies with the ovine model either in its original format using a 5-mm deep annular lesion 23 or in the modified form using a 6 × 20mm lesion 51 it became apparent that a number of IVD regenerative features were also apparent.21,22 Spontaneous repair of the outermost annular layers was a consistent feature with type I collagen spanning the original annular surgical incision. 84 This material had a disorganized structure compared with the collagen fiber organization in mature annular tissue as demonstrated by its reflectile properties when stained with PicroSirius viewed under polarized light. 85 This repair response was confined to the outer annular layers with the inner annular lesion propagating with time into the IVD where it bifurcated, underwent delamellation or developed into concentric or radial tears and also disrupted the normal annular attachments to the cartilaginous endplates.49,50 An influx of cells into the annular lesion site showed some cells expressed α-smooth muscle actin, a myofibroblast marker, these have a contractile phenotype and established roles in wound healing. 42 The main structural protein secreted by fibroblasts is collagen which is consistent with the fibrillar overgrowth tissue in the outer annular lesion and the laying down of thickened annular lamellae.49,50 Fibroblasts are important for the production of a new ECM, which supports cell ingrowth and tissue repair. Blood vessels also play an important role supplying oxygen and nutrients to the cells that proliferate in the repair tissue. An ingrowth of blood vessels into the annular lesion site, and expression of FGF-2 by cells in the outer lesion repair site previously observed supported annular repair.42,43 Expression of TGF-β at the lesion site also promoted collagen expression, cellular proliferation and ECM production conducive to repair processes. 42 A recent study has shown that HA oligosaccharides upregulated MMP production and activation by AF cells in vitro and anabolic gene expression conducive to the clearance of cell and ECM debris from granulation tissue and annular repair. 84 Inflammatory conditions at wound sites leads to HA depolymerization, generating the HA oligosaccharides, which promote the clearance of ECM debris from the defect site by induction of MMPs. These are angiogenic and in combination with TGF-β1, tumor necrosis factor–inducible gene 6 protein (TSG-6) and inter-α-trypsin inhibitor (ITI) can transform fibroblasts to a myo-fibroblastic phenotype conducive to wound repair.86-89 As shown in the present and earlier studies, localized thickening of annular lamellae was prominent in the repair of the outer 5 mm and 6 × 20 mm lesion.22,49,50,84 The 6 × 20 mm lesion is a large defect; its repair is a significant finding. Even so, biomechanical testing of annular repair tissue showed it to be biomechanically inferior to control tissue.29,30,90 Stabilization of the annular lesion site by pedicle plate fixation failed to improve annular repair 91 and also demonstrated the unstable nonhealed inner lesion was susceptible to propagation92,93 contributing to its poor healing properties 94 and alteration of the multidirectional flexibility of the lumbar spine. 95 The mechanical de-stabilisation induced by an outer AF lesion formed the basis of the development of the 6 × 20 mm annular lesion model of experimental disc degeneration used in the present study. 51 Internal stresses generated by cycles of compression-relaxation during normal dynamic spinal loading leads to micromotion/instability, which propagates the annular lesions. Whole body vibration induces disc degeneration possibly through physically exacerbating preexisting fatigue microfractures in disc ECM components 96 leading to the development of degenerative lesions and also induces inflammatory mediators such as interleukin-1β (IL-1β), multiple MMPs, and generates LBP. 97
Chondroid metaplasia was observed in ~20% of the 6 × 20 mm lesion IVDs in the present study but not in the 5-mm lesion samples. This is a benign condition, occurring in connective tissues in response to chronic mechanical stress and may explain why it was not observed with the 5-mm lesion. 98 Chondroid metaplasia of paraspinal tissues in the degenerative spine also occurs during remodeling of the erector spinae muscle at the fourth lumbar vertebra 99 and is used to describe degenerative histopathology occurring in tendinopathy. 100 The term chondroid metaplasia was introduced in the spinal work of Hansen 101 on the morphogenesis of aging disc tissues and the early onset of disc degeneration in chondrodystrophoid dog breeds but is also now recognized as a feature of nonchondrodystrophoid canine IVDs. 102 Chondroid metaplasia is defined as a proliferation of rounded chondrocyte-like cells surrounded in a hyaline cartilaginous matrix thus is an appropriate descriptive term for the NP outgrowth tissue we observed in the present study. This GAG-rich tissue contained proteoglycans decorated with the 7-D-4 and 3-B-3(−) CS sulfation motifs. These have also been observed in fetal human knee, hip, and elbow joint development where they are synthesized by resident chondroprogenitor cell populations.74,103-106 Chondroid cell colonies have also been observed in the ovine IVD and have been isolated from human IVD tissue as discrete groups of rounded cells surrounded by a basophilic pericellular matrix. 107 These cells expressed the 7-D-4 and 3-B-3(−) CS sulfation motifs and the NP stem cell markers CK8 and CK19 107 and had a cell surface CD protein profile by flow cytometry consistent with their identity as a progenitor stem cell population. 107 An earlier study immunolocalized aggrecan as a component of the basophilic pericellular matrices and ECM surrounding chondroid cell nests in ovine IVDs. 108 HA was also present pericellularly around cells within these chondroid cell nests. HA is a common component of the stem cell niche environment.109-111
As found in earlier studies21,22,108 the chondroid cells were significantly larger than the NP cells of the current study. Versican and aggrecan were previously shown to be ECM components of the NP consistent with its identity as a fibrocartilaginous tissue 108 ; however, versican was not detected in the ECM of the chondroid cell colonies demonstrating a clear difference between these tissues. In the present study, we also observed small groups of cells in the inner AF adjacent to the annular lesion atypical of disc cells in control IVD tissues. These appeared similar to cell clusters described in fibrillated articular cartilage in OA and which have been suggested to represent adult stem cells. 108 We previously suggested they may arise from chondroid cell nests in the NP, and proposed these were progenitor cells that became activated due to altered micromechanical effects induced by annular lesions in a process resembling the effect of surface fibrillation on cell clustering in OA articular cartilage.73,112 These cell clusters in the inner AF were surrounded in a GAG-rich matrix showing they were metabolically active and were capable of cell division; however, they were incapable of repairing the inner annular lesion. A chondroid tissue mass growing from an extension of the NP into the inner aspects of the annular lesion in ~20% of the lesion IVDs was a prominent feature of the present study. This tissue integrated with the inner annular lamellae distorting the normal arcade structure of this tissue but prevented the bifurcation of the annular lesion into inner aspects of the IVD apparently stabilizing the defect. This chondroid tissue was most prominent 3 months PO, and by 6 months, it had resorbed to a major extent leaving behind annular tissue of a normal appearance. We interpreted this as a reparative response since the lesion did not propagate further into the disc unlike other IVDs where the lesion propagated as far as the contralateral AF in the absence of chondroid metaplasia. Chondroid metaplasia occurred in only ~20% of the lesion affected IVDs, the reasons for this are currently unknown.
Quantitation of the 7-D-4 and 3-B-3(−) epitopes by ELISIA confirmed the histological findings demonstrating an increase in the AF1 zone (but not AF2) accompanied by a depletion in sulphated GAG levels. These CS sulfation motifs are also expressed in OA in fibrillated articular cartilage 67 ; however, the significance of this observation in ovine IVD tissues requires further evaluation.
Footnotes
Acknowledgments and Funding
Profs. Andrew and Christine Dart (Sydney University Veterinary Hospital) and Prof. Christopher B. Little (Institute of Bone and Joint Research, University of Sydney) are thanked for undertaking the sheep surgeries used in this study. The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by NHMRC Project Grant 1004032.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
All animal welfare and ethics for this work were approved by the University of Sydney Animal Care and Ethics Committee under ethics approval A45/6-2011/3/5544.
Animal Welfare
The present study followed international, national, and/or institutional guidelines for humane animal treatment and complied with relevant legislation.