
Abstract
Objective:
The objective of the present study was to investigate the presence and distribution of 2 basement membrane (BM) molecules, laminin and collagen type IV, in healthy and degenerative cartilage tissues.
Design:
Normal and degenerated tissues were obtained from goats and humans, including articular knee cartilage, the intervertebral disc, and meniscus. Normal tissue was also obtained from patella-tibial enthesis in goats. Immunohistochemical analysis was performed using anti-laminin and anti–collagen type IV antibodies. Human and goat skin were used as positive controls. The percentage of cells displaying the pericellular presence of the protein was graded semiquantitatively.
Results:
When present, laminin and collagen type IV were exclusively found in the pericellular matrix, and in a discrete layer on the articulating surface of normal articular cartilage. In normal articular (hyaline) cartilage in the human and goat, the proteins were found co-localized pericellularly. In contrast, in human osteoarthritic articular cartilage, collagen type IV but not laminin was found in the pericellular region. Nonpathological fibrocartilaginous tissues from the goat, including the menisci and the enthesis, were also positive for both laminin and collagen type IV pericellularly. In degenerated fibrocartilage, including intervertebral disc, as in degenerated hyaline cartilage only collagen type IV was found pericellularly around chondrocytes but with less intense staining than in non-degenerated tissue. In calcified cartilage, some cells were positive for laminin but not type IV collagen.
Conclusions:
We report differences in expression of the BM molecules, laminin and collagen type IV, in normal and degenerative cartilaginous tissues from adult humans and goats. In degenerative tissues laminin is depleted from the pericellular matrix before collagen type IV. The findings may inform future studies of the processes underlying cartilage degeneration and the functional roles of these 2 extracellular matrix proteins, normally associated with BM.
Introduction
Throughout life, chondrocytes are continually engaged in processes related to the maintenance and remodeling of cartilage. In the aging population, chondrocytes undergo major changes in degenerative diseases such as osteoarthritis (OA) and degenerative disc disease affecting the chondrocytic phenotype and the overall tissue composition, architecture, and properties, with many critical aspects of the mechanisms involved in cartilage homeostasis, remodeling, and degeneration still unknown. The distinct pericellular zone of extracellular matrix (ECM) immediately surrounding chondrocytes may hold keys to understanding these mechanisms. As noted in a previous review, 1 light microscopy studies dated to 1888 and electron microscope investigations over the past several decades have described the various ECM structures surrounding chondrocytes. Even allowing for the fact that some of the pericellular features observed by light and electron microscopy may be artifacts of tissue processing,2,3 it is clear that there is a distinct zone of pericellular matrix (PCM) just outside the cell membrane, which differs in its composition and ultrastructure from (and separates the cell from) the interterritorial ECM of cartilage.3,4
In many tissues of the body (viz., muscle and nerve tissue types) the separation of cells from their ECM is due to the presence of basal lamina, and in most organs the separation of various tissue types (viz., epithelium-connective tissue, epithelium-muscle, and epithelium-epithelium) is due to the presence of basement membrane (BM). The BM is traditionally thought of as a layer of certain proteins, notably laminin and collagen type IV, and proteoglycans that is visible even with light microscopy, adhering: epithelium to connective tissue (e.g., skin and blood vessels); epithelium to smooth muscle (e.g., ureter); and epithelium to epithelium (e.g., the glomerulus). Of the ultrastructurally distinct layers comprising the BM in the transmission electron microscope, one—the basal lamina—can be seen enveloping muscle, nerve, and fat cells, 5 separating them from their ECM.
Several molecules normally associated with BM have been reported in cartilage, and some specifically in the PCM, including laminins6-8 and collagen type IV.7,9,10 In recent years, groups have identified select BM molecules in a number of cartilage tissues in various species.6,7,9,11,12 We have recently described the temporal expression of laminin and collagen type IV in chondrogenesis of mesenchymal stem cells in pellet cultures. 13 Despite the likely prospect that pericellular BM molecules play a role in the physiology and pathology of cartilage, there have been no prior studies of the distribution of laminin and type IV collagen in the PCM of arthritic hyaline and degenerative intervertebral disc cartilages in humans, prompting this investigation.
The aim of the present study was to investigate the spatial distribution of laminin and collagen type IV in normal and osteoarthritic cartilage tissues of different type and origin by immunohistochemical analysis. Because of the difficulty in obtaining reliably healthy human cartilages we also included healthy cartilage samples from the goat for comparison.
Methods
Tissues
Human samples, which were obtained with approval of our institutional review board, included the following: articular cartilage and meniscus from patients undergoing total knee arthroplasty for osteoarthritis; intervertebral disc (IVD) specimens from human autopsies and from patients undergoing discectomy; and skin samples archived in the Department of Pathology ( Table 1 ). Goat tissues obtained from skeletally mature goats included the following: articular cartilage; IVD; menisci; and patellotibial enthesis ( Table 1 ). To validate the presence of basement membrane molecules in healthy human cartilage tissues as shown by others6,14 we analyzed the nondegenerated margins of the resected tissue from 2 non-osteoarthritic patients undergoing autologous chondrocyte implantation. The normal articular cartilage histology of these samples was verified on hematoxylin and eosin–stained sections.
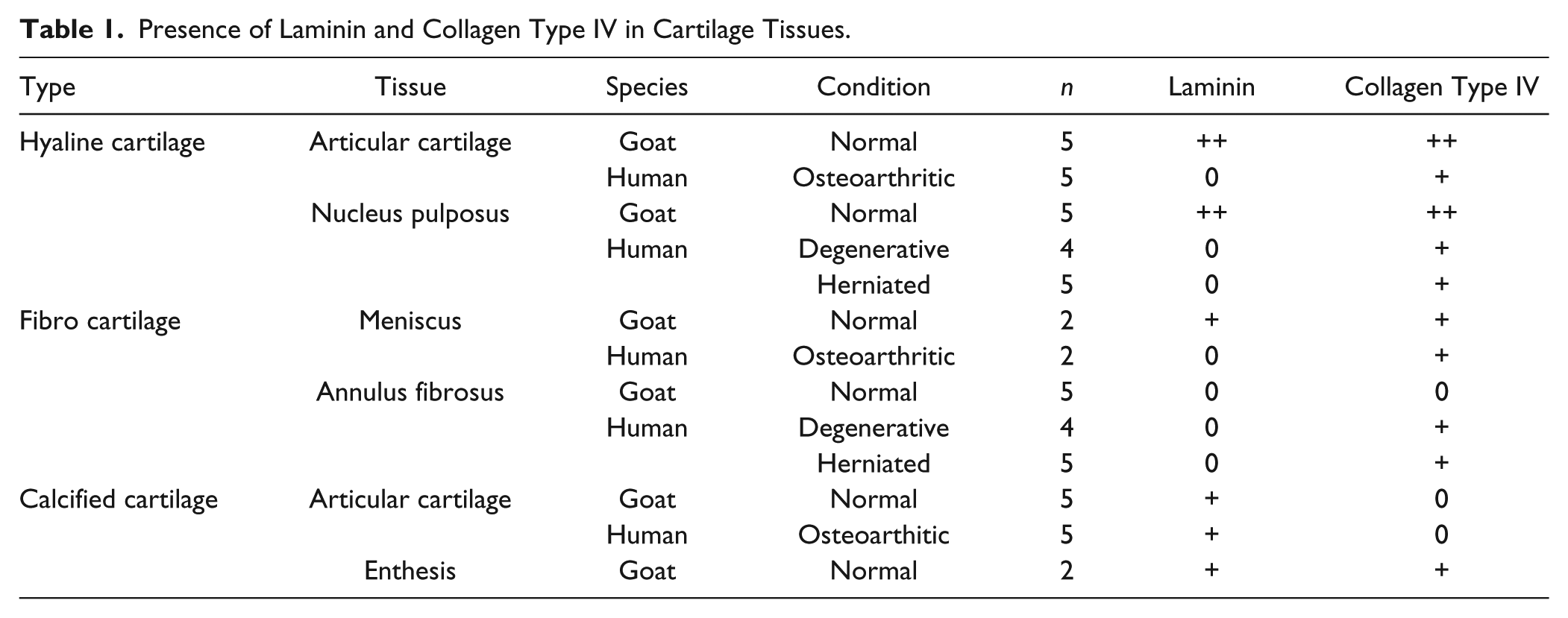
Presence of Laminin and Collagen Type IV in Cartilage Tissues.
Tissue Processing and Preparation
All tissues were fixed in 10% formalin for up to 1 week. Adjacent bone was decalcified using ethylenediamine tetraacetic acid. The tissue was dehydrated in a graded series of alcohol (70% to 100%), cleared in xylene, and embedded in paraffin (Tissue Processor, Leica Microsystems, Buffalo Grove, IL). The embedded tissue was cut into 7-µm thick sections (Finesse ME, Thermo Scientific, Kalamazoo, MI) and mounted on microscope slides, and put on a slide warmer overnight. The sections were then deparaffinized in xylene, rehydrated in a graded series of alcohol (100% to 80%), and rinsed in water, prior to histological and immunohistochemical staining.
Histology and Immunohistochemistry
Sections were stained with hematoxylin and eosin for evaluation of tissue morphology. Immunohistochemistry with polyclonal rabbit antibodies for laminin (Abcam, ab11575, Cambridge, MA) and collagen type IV (Abcam, ab6586) was performed on all tissues using a Dako Autostainer (Dako Universal Staining System, Carpinteria, CA). The slides were pretreated with protease for 40 minutes, and then incubated with the primary antibody for 30 minutes at room temperature. Negative controls in each staining cycle were labeled with negative control rabbit immunoglobulin (Dako X0903) instead of the primary antibody but with the same concentration as the primary antibody. Labeling was developed using streptavidin–horse radish peroxidase and aminoethyl carbazole following the manufacturer’s instructions (Dako). The slides were counterstained with Mayer’s hematoxylin.
Evaluation
The evaluation was performed on light microscope (Olympus BX51, Olympus America, Center Valley, PA) images captured using Picture Frame Software (Picture Frame, version 2.3; Karl Storz Imaging, Goleta, CA). The tissue morphology was defined on hematoxylin and eosin sections. Polarized light microscopy was employed to confirm differentiation between hyaline and fibrous ECM, based on the birefringent appearance of the collagen bundles—particularly in sections in which the distinction was questionable using conventional transmitted light. The location of the staining with the antibodies was noted for each slide. In order to determine the consistency and differences in our findings, we compared the presence and distribution of the 2 basement membrane molecules in normal chondrocytes in articular cartilage with nucleus pulposus, normal hyaline cartilage with fibrocartilage, normal hyaline cartilage with degenerative hyaline cartilage, and normal fibrocartilage with degenerative fibrocartilage. A semiquantitative evaluation of the cell staining was performed. Depending on the percentage of cells with positive PCM staining, the sections were assigned a grade from 0 to +++: no cell staining (0); <5% of cells staining (+); 5% to 50% (++); and >50% (+++). The percentages of samples with various semiquantitative grades were analyzed using Fisher’s exact test. Two-tailed P values less than 0.05 were considered significant.
Results
General Findings
Laminin and collagen type IV consistently surrounded chondrocytes in healthy hyaline cartilage in goats ( Fig. 1 ; Table 1 ), and were also found in nonosteoarthritic, normal appearing articular cartilage from humans ( Fig. 1 ). In stark contrast, in normal goat meniscus fibrocartilage, no positive stain for either of the basement membrane molecules was observed even though the cells displayed the typical chondrocyte morphology, being rounded and residing in lacunae ( Table 1 ). No nonchondrocytic cell (i.e., not residing in a lacuna) was positive for either of the basement membrane molecules in any of the tissues, except for the basement membrane in blood vessels, which served as an internal positive control when present. No positive stain staining of laminin or collagen type IV was observed in the ECM (interterritorial matrix). When positive staining was observed, it appeared in a continuous halo surrounding the cells, with a thickness of 0.5 to 1 µm. While the continuity of the halo was consistent, variation in the intensity of the chromogen was observed. Goat chondrocytes in hyaline matrix (nonhypertrophic) were approximately 10 µm in diameter (i.e., the internal diameter of the halo), while human chondrocytes (nonhypertrophic) in hyaline matrix were approximately 12 to 15 µm in diameter. The human and goat skin samples that were used as external, intraindividual positive controls displayed positive staining for laminin and collagen type IV in the basement membrane underlying the epidermis and surrounding the blood vessels (data not shown). None of the negative immunohistochemical control sections displayed the red chromogen indicative of positive staining ( Fig. 2 ).
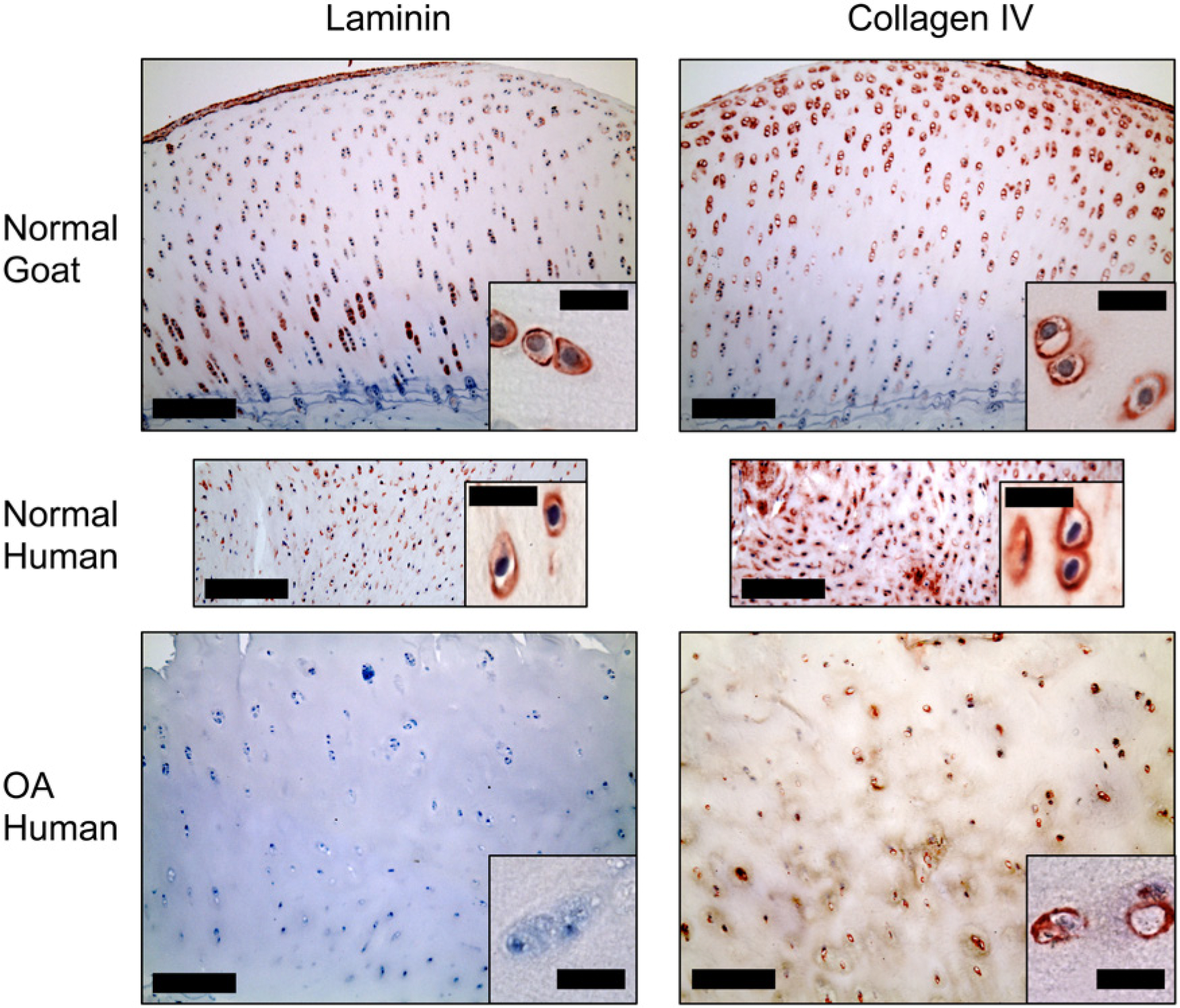
Articular cartilage. Pericellular stain for collagen type IV was seen in both normal and degenerated cartilage. Laminin-positive pericellular stain was only seen in normal articular cartilage. Bars: Large image = 200 µm, small image = 20 µm. OA = osteoarthritis.
Negative controls did not show positive stain. Bar = 30 µm.
Articular Cartilage
In the normal goat articular cartilage both laminin and collagen type IV were clearly seen in the chondrocyte PCM ( Fig. 1 ) in all of the 5 samples ( Table 1 ) with a grade of ++. Collagen type IV showed a more intense stain in the PCM of the superficial zone cells with a decreasing intensity in the PCM of cells toward the deep zone. Superficial chondrocytes appeared to have thicker band of pericellular collagen type IV positive stain when compared to deeper zone chondrocytes, but this could not be confirmed to a satisfactory degree using light microscopy. Clusters of proliferating cells were positive for collagen type IV but not laminin (data not shown). Positive staining for type IV collagen was only very rarely found in the calcified cartilage zone, whereas laminin was often found below the tidemark ( Fig. 3 ). Conversely, the laminin staining was more homogenous throughout the layers of the cartilage but appeared more intense in regions of cell columns. Intact samples of normal goat articular cartilage also demonstrated a distinct layer of laminin and type IV collagen on the articulating surface.
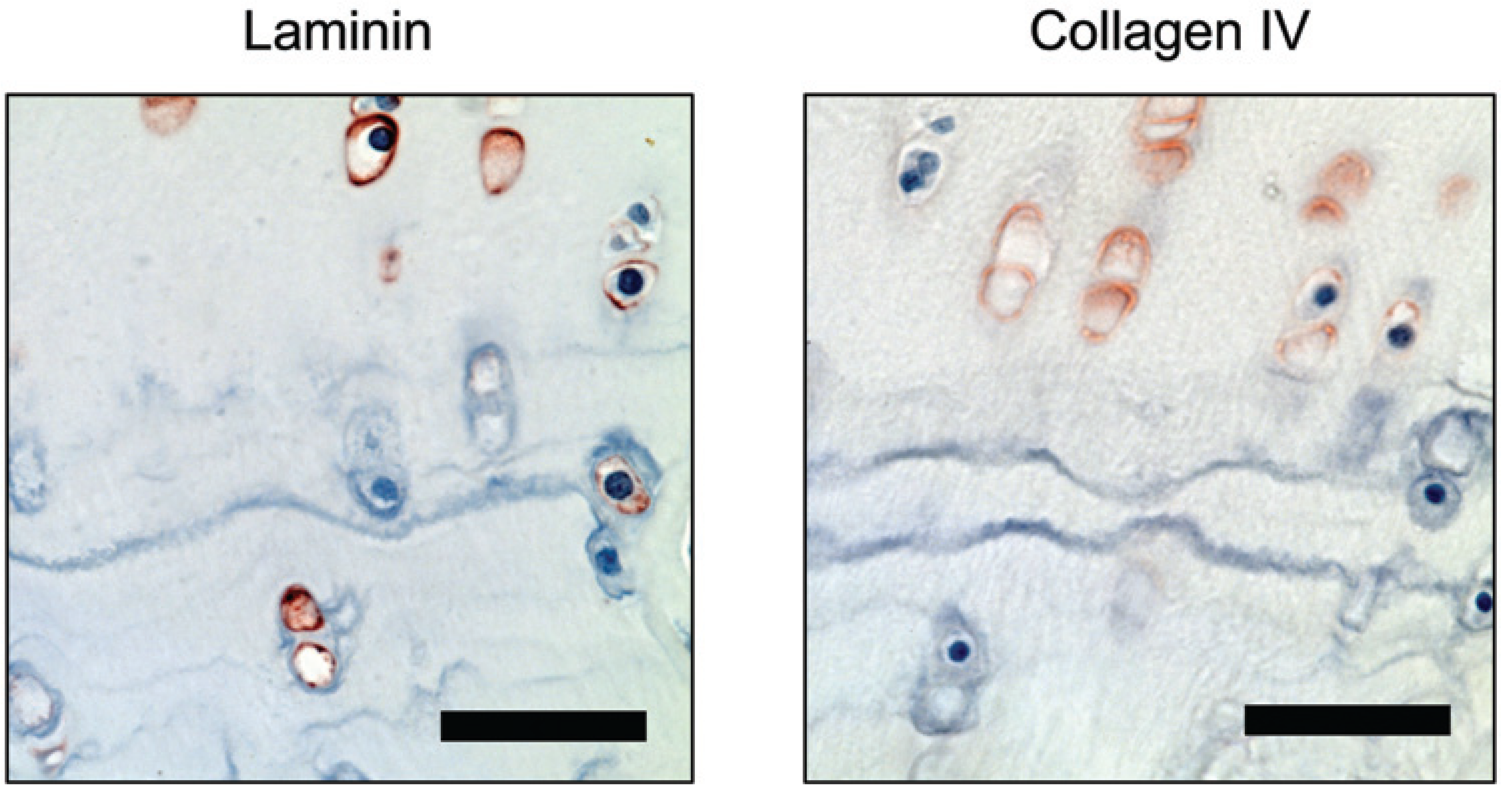
Articular cartilage. Pericellular stain for was found under the tidemark whereas collagen type IV was generally absent in the pericellular matrix below the tidemark. Bar = 50 µm.
In comparison, human OA cartilage was negative for laminin in all of the 5 samples, while collagen type IV staining was evident, although to a lesser extent than that seen in healthy cartilage in the goat ( Fig. 1 ), with a grade of + (<5% of cells). Comparing the 5 of 5 normal goat samples staining positive for laminin with the 0 of 5 human OA samples, Fisher’s exact test yielded a highly statistically significant difference with P = 0.008. Laminin and collagen type IV appeared to co-localize consistently in the PCM of healthy chondrocytes in caprine hyaline cartilage, while this was not the case in the OA samples, where collagen type IV was sometimes found without the presence of laminin.
Intervertebral Disc
In the healthy goat samples, the PCM in the nucleus pulposus was positive for laminin and collagen type IV ( Fig. 4 ) in all of the 5 samples ( Table 1 ) while the annulus fibrosus region was predominantly negative for both molecules ( Fig. 5 ). Here again, Fisher’s exact test yielded P = 0.008, when comparing the number of samples staining for laminin in the two regions. The human degenerative disc tissue from autopsy and discectomy was negative for laminin in all samples, while positive pericellular staining for collagen type IV was found in some areas in both the nucleus pulposus and annulus fibrosus regions ( Table 1 ). More collagen type IV pericellular stain was found in the discectomy samples compared to the autopsies. Just as was found in articular cartilage, cell clusters of proliferating cells were positive for collagen type IV but not laminin ( Fig. 6 ).
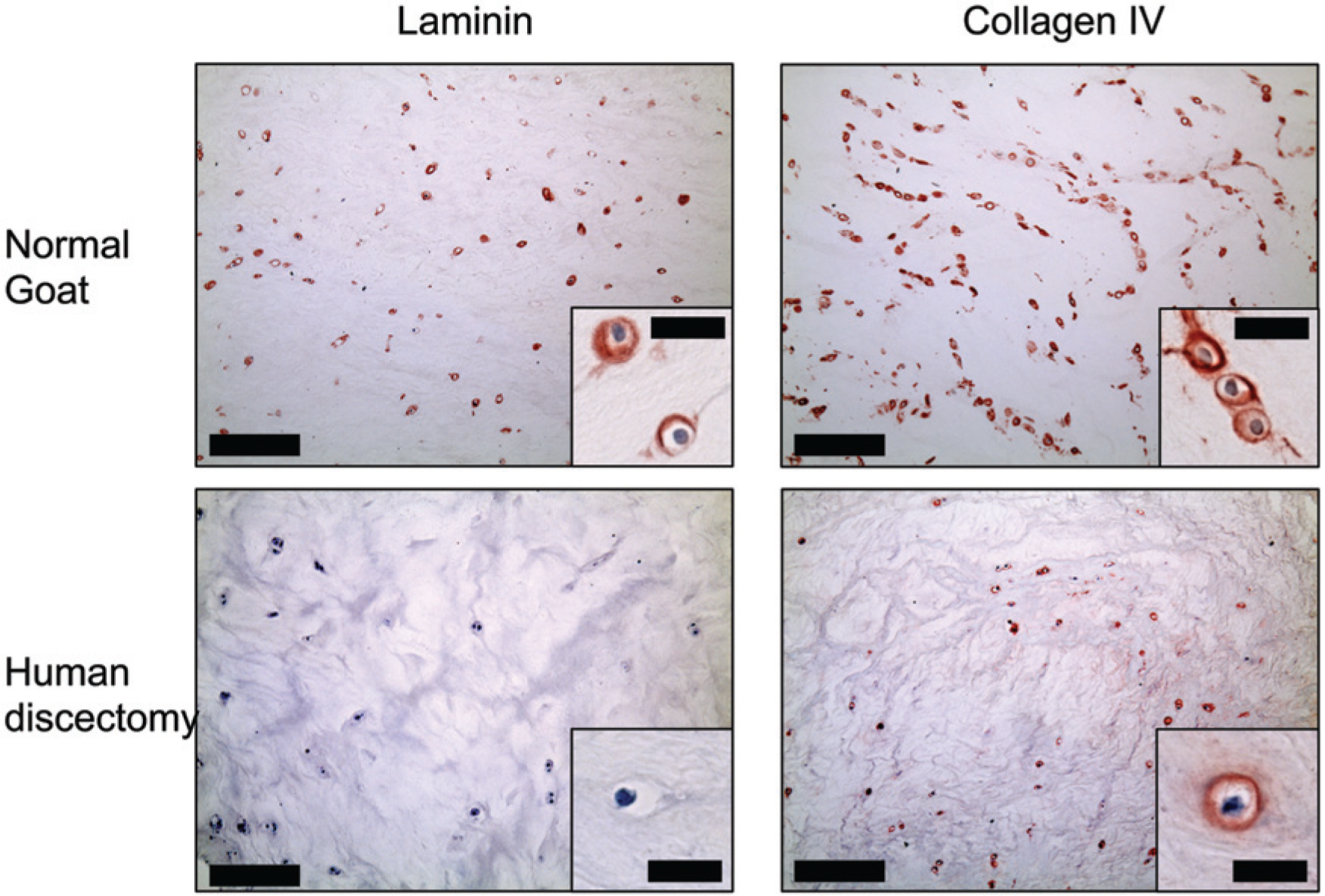
Nucleus pulposus region of the intervertebral disc. Collagen type IV was positive in the region in normal as well as degenerated discs from human autopsy (shown on image) and from discectomy, whereas laminin-positive stain was not observed in the degenerated nucleus pulposus tissue. Bars: Large image = 200 µm, small image = 20 µm.
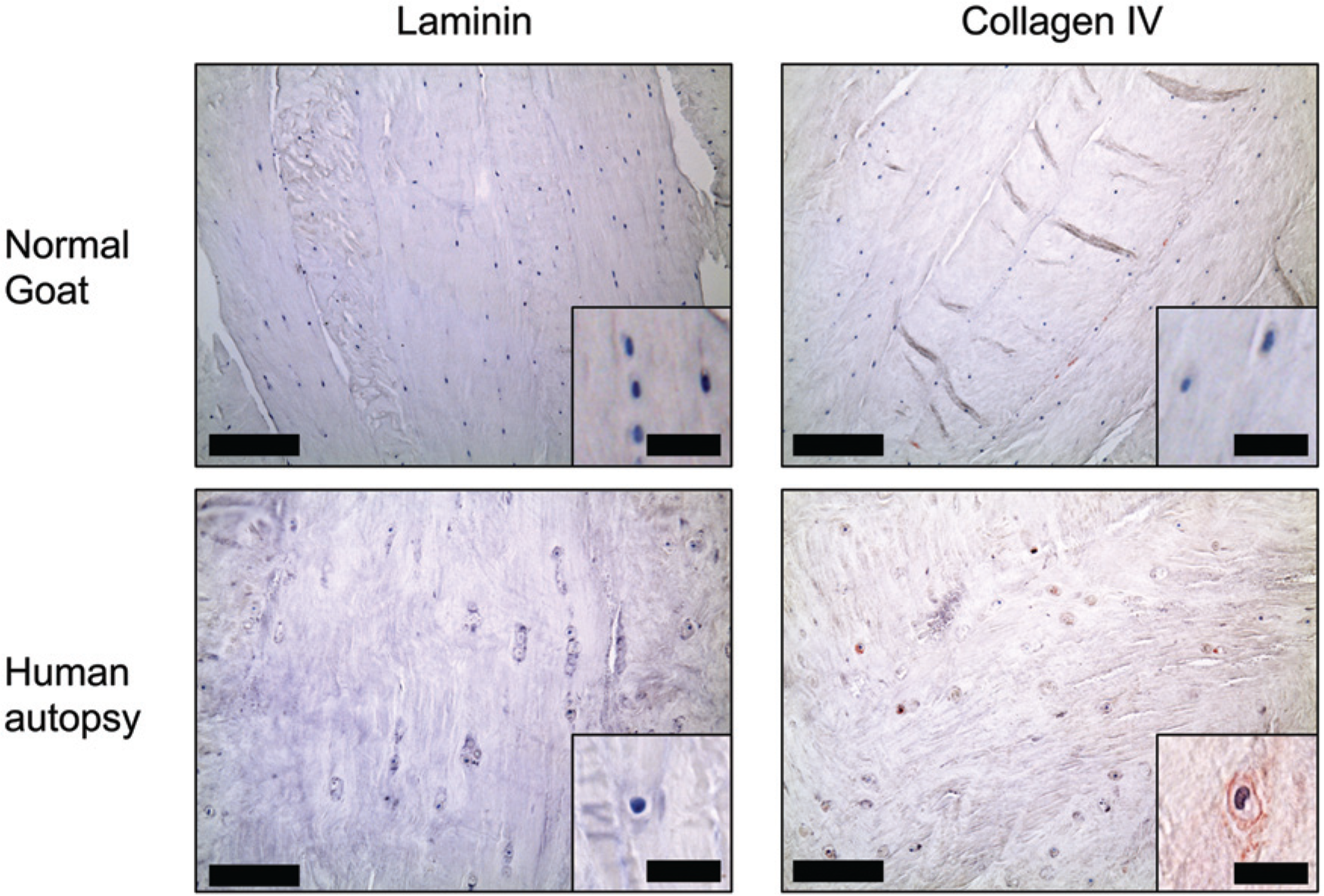
Annulus fibrosus region of the intervertebral disc. Collagen type IV was positive in the annulus fibrosus region in degenerated discs from human autopsy (shown on image) and from discectomy. Bars: Large image = 200 µm, small image = 20 µm.
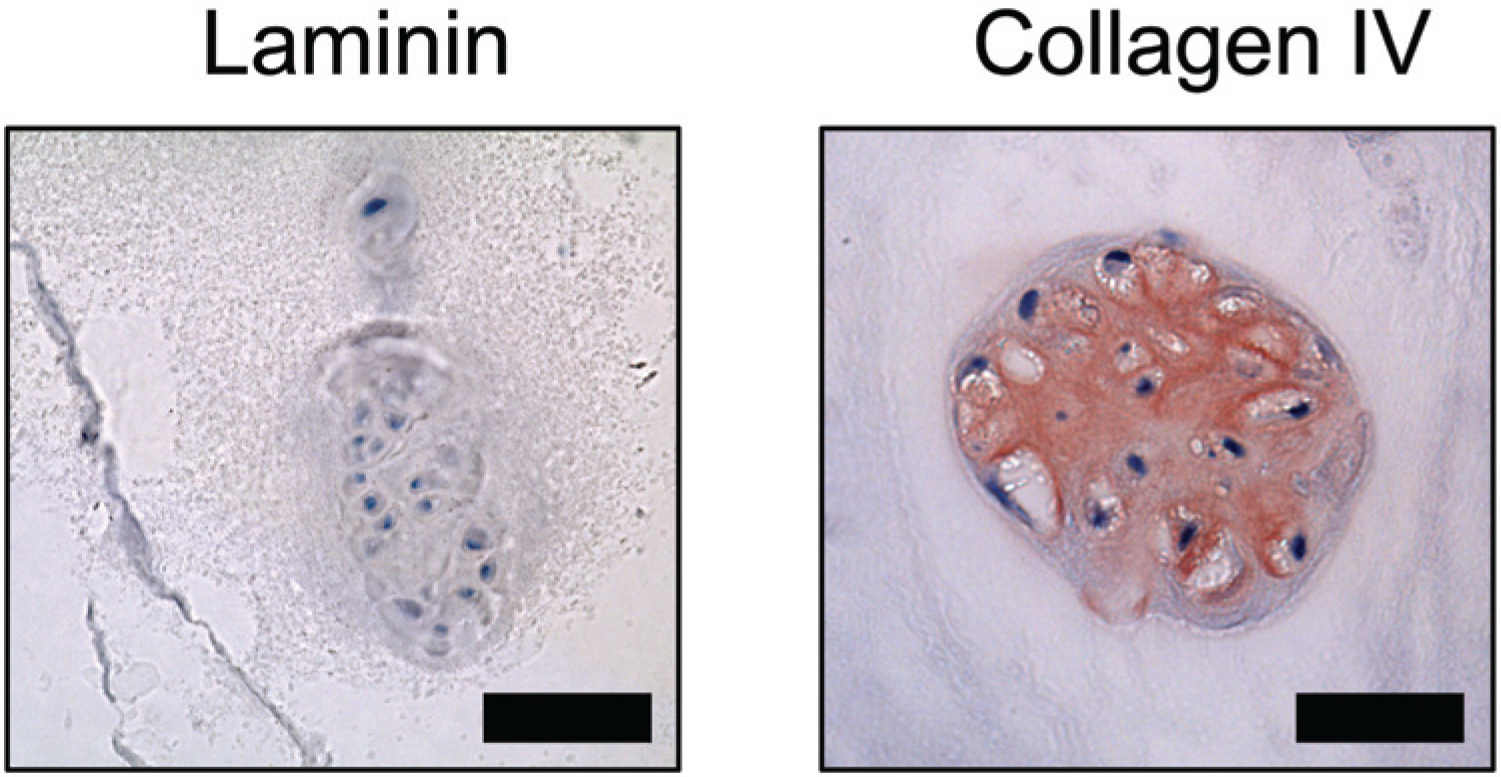
Chondrocyte clusters. Antibody stain of cell clusters in degenerative intervertebral disc cartilage. Clusters were found both in the near the endplate and in the more central regions. Collagen type IV but not laminin-positive stain was found. The collagen type IV stain was not only restricted to the pericellular matrix but was also found in the interterritorial matrix. Bar = 50 μm.
Enthesis
Chondrocytes in the calcified cartilage at the bone-tendon interface between the anterior cruciate ligament and tibia and at the patella-tibial junction stained positive for both laminin and collagen type IV pericellularly. While chondrocytes found in the fibrous part of the enthesis were collagen type IV–positive, a much lower amount of positive cells was found in the calcified zone. For laminin, the majority of positive cells were found in the calcified zone, while very few laminin positive cells were found in the fibrous zone ( Fig. 7 ; Table 1 ).
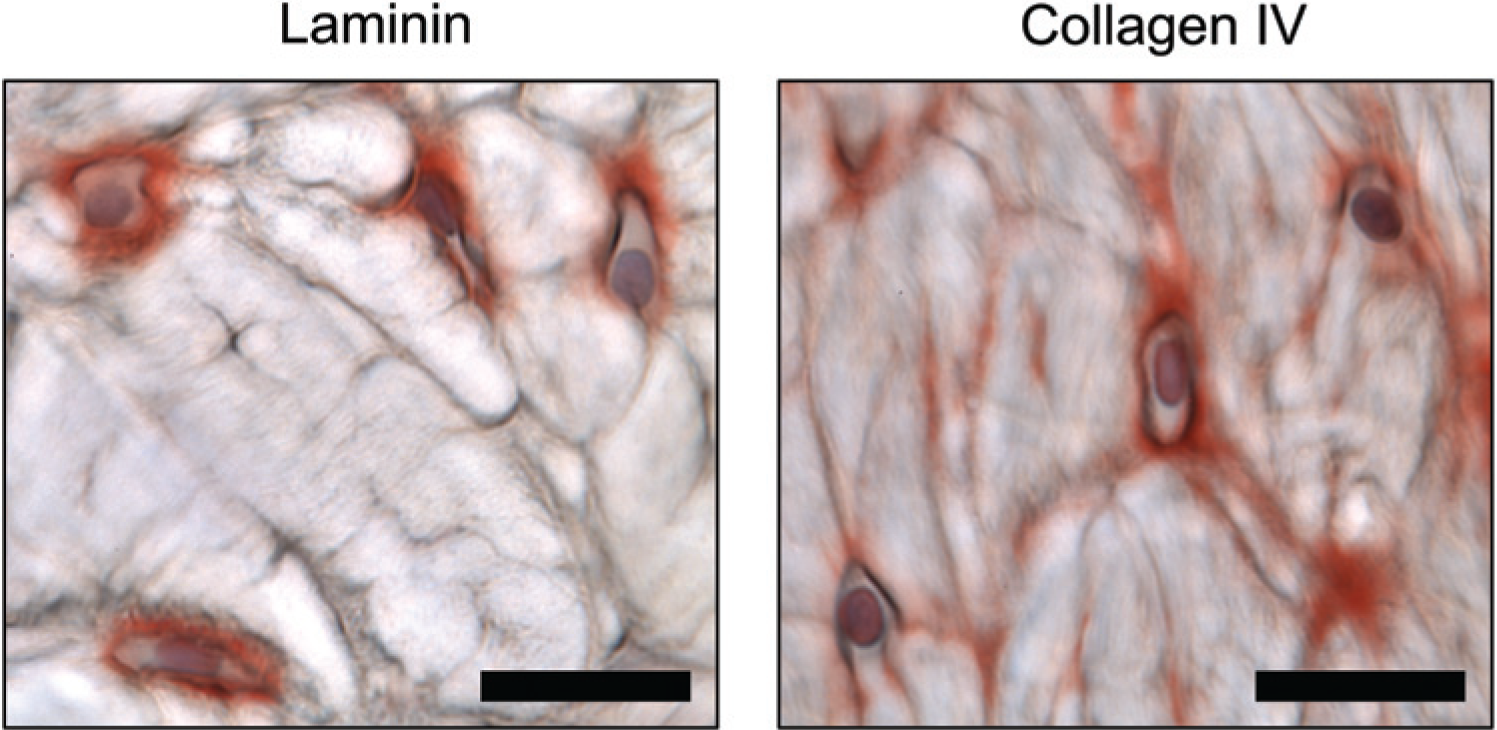
Patellotibial enthesis. Positive pericellular stain for both laminin and collagen type IV of chondrocytes. Laminin positive stain was also found in calcified cartilage, whereas only very few cells showed positive pericellular stain for collagen type IV. Bar = 50 µm.
Meniscus
The menisci from healthy caprine knees stained positive for both laminin and collagen type IV pericellularly ( Fig. 8 ). In contrast to the well-defined and sharp-edged ring localization of the 2 molecules in hyaline cartilage, the appearance in the menisci was a more diffuse pericellular stain. While the continuity of the pericellular ring was intact, we did not observe a defined outer edge of the ring. The menisci from human OA knees were negative for both molecules.
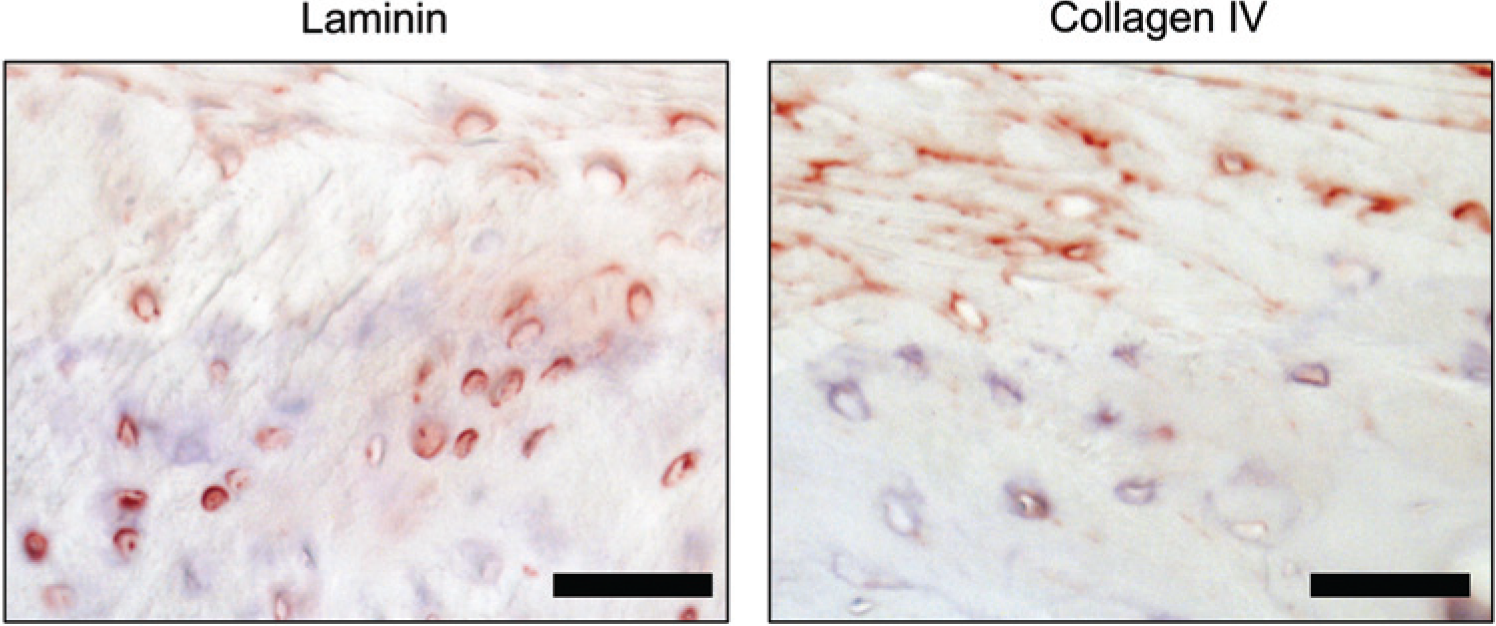
Meniscus. Positive pericellular stain for both collagen IV and laminin was found around chondrocytes in healthy menisci. Bar = 20 µm.
Discussion
This study investigated the spatial distribution of laminin and collagen type IV in normal and degenerated cartilage tissues. Notable findings of the present study dealing with degenerative human cartilaginous tissues and adult healthy cartilaginous tissues from goats, for comparison, include (1) the absence of laminin and diminished staining for collagen type IV in the PCM of degenerative human hyaline cartilage, in contrast to their pronounced presence in healthy (human and caprine) cartilage—articular cartilage and nucleus pulposus and (2) general absence of these 2 BM molecules in the PCM of chondrocytes in healthy and degenerative fibrocartilages—meniscus and annulus fibrosus. Also of note, comparing the appearance and disappearance the these 2 BM proteins, type IV is the first to be expressed as demonstrated in the matrix surrounding proliferating cells in the clusters and the last to be lost during the degenerative process. The expression of laminin and type IV collagen persisted as the cartilage underwent calcification in the formation of the enthesis.
Prior studies of adult tissues have identified laminin in the PCM of chondrocytes of normal superficial zone human articular cartilage 6 and type IV collagen in the PCM of human nucleus pulposus chondrocytes. 9 Other studies identified laminin and type IV collagen in the pericellular and interterritorial matrix of articular cartilage of newborn and adult mice. 7 The current work has expanded the knowledge on the presence of BM molecules in healthy cartilage by showing that both laminin and collagen type IV were prominently found in the PCM of chondrocytes in caprine articular cartilage and nucleus pulposus. In goat articular cartilage, the PCM of a greater percentage of chondrocytes in the deep zone contained laminin, compared to the superficial zone, while the reverse was true for collagen type IV. Of note was that there was virtually no laminin seen in degenerative articular cartilage from OA patients and only a relatively low percentage of cells in these samples displayed the presence of collagen type IV in their PCM. The same was true when comparing the staining for these BM molecules in healthy and degenerative nucleus pulposus.
Differences in the expression between healthy and degenerative cartilage tissue have not previously been described, although the lack of laminin in OA cartilage is supported by previous findings that laminin γ-1 chain mRNA level is decreased in human subjects with mild OA compared to the normal healthy controls. 15 In contrast to the findings in hyaline cartilage, there was little indication of the presence of laminin and collagen type IV in the PCM of chondrocytes in the fibrocartilages, perhaps reflecting the differences in the mechanical environment of hyaline cartilage and fibrocartilage, as well as in the differential organization of the collagen molecules in the ECM of these two cartilage tissues. The presence of laminin, but not collagen type IV, in the calcified cartilage samples, may indicate different roles of these two BM molecules in connective tissues. In the annulus fibrosus region of the IVD, we found a striking difference in the presence of collagen type IV, which was only present in degenerating fibrocartilage (i.e., from elderly patients). Collectively, these findings suggest that while laminin is consistently absent in degenerative tissue, collagen type IV may play a role in the adaptive response to the altered microenvironment and/or biomechanical changes in degenerative diseases in the musculoskeletal system.
The amount of collagen type IV gradually decreases from the superficial surface in articular cartilage, to almost completely disappear in the calcified zone (i.e., beneath the tidemark), while laminin is present in the calcified zone with lesser zonal variation in staining intensity throughout the cartilage ( Fig. 2 ). We also found that hypertrophic chondrocytes are not surrounded by the 2 BM molecules. These findings suggest that laminin and collagen type IV may be involved in regulating vascularization and the progression of chondrocyte hypertrophy toward the formation of calcified cartilage and subchondral bone. A previous study showed that collagen type IV is increased in experimentally induced OA in a dog model. 16 We found a ubiquitous presence of chondrocytes surrounded by collagen type IV in both normal and OA hyaline cartilage. Interestingly, in fibrocartilage tissue, collagen type IV surrounds chondrocytes in degenerative but not in normal fibrocartilage ( Figs. 4 and 5 ). Exceptions were the meniscus and enthesis, in which collagen type IV was found in the PCM in healthy tissue, although with a more diffuse appearance. In addition, only collagen type IV was found in chondrocyte clusters and was not restricted to the PCM ( Fig. 6 ). Two of the 3 collagen type IV heterodimers contain antiangiogenic domains17,18 and the avascular nature of hyaline cartilage and the apparent requirement of hypoxia to sustain the chondrogenic phenotype 19 make it reasonable to postulate that collagen type IV may play a role in exerting these anti-angiogenetic effects on the chondrocytes.
The laminin cell binding–receptor family consists of the integrins α3β1, α6β1, α6β4, and α7β1, 20 and chondrocytes have been shown to bind to laminin though α6β1 integrins.14,21 In fetal knees, Salter et al., 22 found that integrin α6β1 was expressed in chondrocytes in both articular, epiphyseal, and growth plate chondrocytes as well as in meniscal cells, and that both laminin and collagen type IV were found in all these tissues. A subsequent study found that that while the α6B receptor subunit promoted chondrogenesis, α6A expression was necessary for stabilizing the differentiated phenotype. 23 The loss of laminin in degenerated cartilage—regardless of location—in the present study supports this finding, and further proposes the laminin may be used as a marker for phenotypic changes in the chondrocyte in degenerating cartilage. Investigation of the developmental stage–specific location of laminin in articular cartilage in mice showed a widespread appearance in the ECM in the newborn mouse that shifted toward a pericellular appearance in the adult mouse. 7 Chondrocytes are able to bind to collagens through integrin α1β1, α2β1, α10β1, and α11β1 20 and chondrocytes have been shown to bind to collagen type IV through α1β1. 24 Collagen type II is by far the most abundant collagen in healthy hyaline cartilage. However, since the first descriptions of collagen type II–induced arthritis, 25 several studies have suggested that since collagen type II can induce OA, healthy chondrocytes are in fact not in direct contact with collagen type II but interact through other molecules. Thus, a possible function of the PCM may be protection of the chondrocyte from the surrounding ECM.
Based on immunohistochemical and flow cytometry analyses, Chen et al. 26 found that the nucleus pulposus region had higher expression of the laminin 5-α chain and laminin receptors than the annulus fibrosus region in human, porcine and rat IVDs. Investigation of collagen types in the IVD has previously shown the presence of collagen type IV in the adolescent nucleus pulposus region, while this was generally absent in the annulus fibrosus region. Age-related associations in the expression were found for both regions of the IVD, as only little collagen type IV was found in the adult/elderly patient group in the nucleus pulposus region, and minimal amounts of collagen type IV in the annulus fibrosus region in the adolescence group while none in the adult/elderly group. 9 Similarly, the latter was observed in our goat annulus fibrosus samples. Collectively, these findings suggest that while disappearance of laminin from the PCM may serve as an early indicator of degenerative cartilage changes, collagen type IV is more likely to be involved in the response to the degenerative changes.
The limitations in the present study involve the types of tissue collected, as we were only able to obtain human tissue with degenerated cartilage. Obtaining healthy cartilage tissue from patients is very challenging and autopsy tissues mostly obtained from elderly patients are subjected to cartilage degeneration. We were, however, able to validate the presence of laminin and collagen type IV in nondegenerated cartilage from patients with focal cartilage lesions. While there are differences between cartilage tissues from different anatomical locations, it is important to note that the degenerated and degenerating tissues in the present study were collected from patients presenting with different pathological changes. For instance, the articular cartilage tissues were from end-stage OA since these patients were undergoing total knee arthroplasty, while the degenerative changes in the IVD were only age related and might even be asymptomatic. Immunohistochemical analyses are always limited by the possibility for epitope-masking of the target sequences by other molecules. Thus, PCM degradation in degenerative tissues may reveal appearance of collagen type IV, which we find in degenerative but not in normal fibrocartilage.
In summary, we described the distribution of laminin and collagen type IV in the PCM in healthy and degenerative cartilaginous tissues. The molecules were widely expressed in the PCM of healthy, nondegenerative hyaline cartilage and generally absent in fibrocartilage (except menisci and enthesis). Collagen type IV could be found in the PCM of degenerative hyaline cartilage while laminin was consistently absent. Laminin, but not collagen type IV, was found in calcified cartilage from healthy and osteoarthritic joints, suggesting this molecule as an early marker for cartilage degeneration and/or chondrocyte dedifferentiation. Another argument for this is the loss of laminin in the PCM in OA cartilage and degenerated nucleus pulposus tissues.
Footnotes
Acknowledgments and Funding
The research reported here was supported, in part, by the U.S. Department of Veterans Affairs, Veterans Health Administration, Rehabilitation Research and Development Service. A postdoctoral research fellowship for CBF by an Elite Research Scholarship from the Danish Minister of Science, Technology and Innovation. WST is supported by A*STAR International Fellowship, Agency for Science, Technology and Research (A*STAR) Singapore. MS is supported by a Research Career Scientist Award from the Department of Veterans Affairs. The authors would like to acknowledge research fellow Thomas Cheriyan, MD, for his technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study was approved by our institutional review board.