
Abstract
Objective:
Describe the local morphological response of the articular–epiphyseal cartilage complex to surgical stab incision in the distal femur of foals, with emphasis on the relationship between growth cartilage injury, enchondral ossification, and repair.
Design:
Nine foals were induced into general anesthesia at the age of 13 to 15 days. Four full-thickness stab incision defects were created in the cartilage on the lateral aspect of the lateral trochlear ridge of the left distal femur. Follow-up examination was carried out from 1 to 49 days postoperatively, including examination of intact bones, sawed slabs, and histological sections.
Results:
Incision defects filled with cells displaying fibroblast-, chondrocyte-, and osteoblast-like characteristics, potentially validating the rationale behind the drilling of stable juvenile osteochondritis dissecans lesions in children. Incisions induced necrosis within the cartilage on the margins at all depths of the defects. Sharp dissection may therefore be contraindicated in cartilage repair in young individuals. Incisions caused a focal delay in enchondral ossification in 2 foals, apparently related to the orientation of the incision defect relative to the direction of ossification. Defects became progressively surrounded by subchondral bone, in which granulation tissue containing clasts and foci of osteoblast-like cells was observed. Continued enchondral ossification was therefore likely to result in healing of uncomplicated defects to morphologically normal bone.
Conclusions:
Epiphyseal growth cartilage injury had the potential to exert a negative effect on enchondral ossification. Enchondral ossification exerted a beneficial effect on repair. This relationship warrants consideration in future studies of cartilage injury and repair within the articular–epiphyseal cartilage complex of all species.
Keywords
Introduction
Articular cartilage injury and repair is a major topic in orthopedic research. There may, however, still be gaps to bridge in the historical and current knowledge.1-4
A number of experimental studies have been carried out in skeletally mature individuals, where the epiphyseal ends of appendicular long bones are covered simply by a layer of avascular, hyaline articular cartilage.1,2 This is different from skeletally immature individuals, where the covering of bone ends also includes a layer of epiphyseal growth cartilage, located between the articular cartilage and the subchondral bone of the secondary, epiphyseal center of ossification.5-7 The articular and epiphyseal growth cartilages are collectively known as the articular–epiphyseal cartilage complex. 6 The function of growth cartilage is to achieve growth by a process of enchondral ossification.5-7 Injury to growth cartilage may therefore have ramifications beyond the repair of the local cartilage because of the risk of such injury affecting the process of enchondral ossification. In the metaphyseal growth plate or physis, the potential for injury to lead to premature cessation of enchondral ossification, with consequential relative limb shortness, is well recognized.8,9 A similar relationship between injury and the process of enchondral ossification does not appear to have been considered for the articular–epiphyseal cartilage complex.
The articular–epiphyseal cartilage complex is thicker than the adult articular cartilage, and temporarily receives a blood supply during the early phases of growth via so-called cartilage canals.10-13 In animals, it has been hypothesized that failure of the blood supply to epiphyseal growth cartilage leads to ischemic chondronecrosis, an associated focal delay in enchondral ossification (articular osteochondrosis, OC) and pathological cartilage fracture (osteochondrosis dissecans).14,15 The report that follows represents part of a larger study to test this hypothesis experimentally by transecting the blood supply using surgical stab incisions. Following transection, ischemic chondronecrosis was predicted to occur within the epiphyseal growth cartilage distant from the incision (in press). 16 It was not known whether the stab incisions would induce a morphological response within the cartilage locally at the transection sites, or indeed whether the incisions themselves would induce a focal delay in enchondral ossification or cartilage fracture. During the planning of the experiment, the existing literature was searched extensively. Although a number of studies in skeletally immature animals were discovered, they did not answer the questions at hand in a satisfactory manner. It was therefore considered important to examine and report the local morphological changes at the site of transecting incisions, and the relationship between incisions and the process of enchondral ossification, in their own right.
The current stab incisions represent a procedure for the purpose of modeling a disease process and are, thankfully, not identical to any surgical procedure that is currently used in skeletally immature individuals. By virtue of being carried out in skeletally immature foals, however, systematic study of the current stab incisions can nevertheless generate information that may be relevant to certain aspects of surgical procedures in children. Histologically validated study of cartilage injury and spontaneous or surgical repair is not likely to be ethically defensible in children. In adult individuals, the bone in which cartilage injury and repair can be modeled most accurately is the distal femur of the horse.17,18 When histologically validated studies are carried out in animals, and perhaps particularly in foals, the results should be reported and maximally exploited in terms of their potential translational value to cartilage injury and repair in children.
The aim of the current study was to describe the local morphological response of the articular–epiphyseal cartilage complex to 4 full-thickness surgical stab incisions in the distal femur of young foals, with a particular emphasis on the relationship between incisions and the process of enchondral ossification, and on the potential translational value of the results to cartilage injury and repair in children.
Methods
Animals
Stallions (n = 4) and mares (n = 9) of the Norwegian Fjord Pony breed were recruited and confirmed free of radiographic signs of femoropatellar joint disease, to minimize disease predisposition among the offspring. Foals of both genders born to the screened parents were eligible for inclusion. Diagnosis of any form of orthopedic, medical, or surgical disorder was a criterion for exclusion prior to the start of the study and for veterinary intervention during the study.
Creation of Incision Defects
Creation of incision defects was scheduled for as close to the age of 14 days as possible, while still within weekday operating hours. Each foal received flunixin meglumine (1.1 mg/kg body weight intravenously [bwt i.v.]), sodium benzylpenicillin (22,000 IU/kg bwt i.v.), and gentamicin (6.6 mg/kg bwt i.v.) before being induced into general anesthesia. An arthroscopic approach to the lateral pouch of the left femoropatellar joint was made. The lateral trochlear ridge was selected because it is one of the most common sites for the development of spontaneous lesions of OC. The blood supply on the lateral aspect of the lateral trochlear ridge was inspected, and 2 adjacent, radially oriented vessels in the mid to proximal third of the ridge were selected and designated distal and proximal, respectively. An instrument portal was created at the level of the distal selected vessel and a 3-mm wide miniature edged blade, mounted on a holder, was introduced into the joint ( Fig. 1A , Ref. No. EL-6400, Eagle Labs, Rancho Cucamonga, CA). Stab incisions were positioned so as to transect the distal selected vessel in 2 separate locations: a cranial location, near the crest of the lateral trochlear ridge, and a lateral location, near the capsule attachment of the lateral pouch of the joint. In each case, the scalpel blade was advanced until it made contact with the subchondral bone, and lengthened until it measured approximately 6 mm. A minimal amount of bleeding was observed from the incision site. A second instrument portal was created and the procedure repeated for the proximal selected vessel, until a total of 4 separate stab incisions had been made. The incisions were referred to by their relative anatomical locations as the proximal cranial, proximal lateral, distal cranial, and distal lateral incision, respectively ( Fig. 1B ). Morphine, expanded to a total volume of 5 mL in 0.5% bupivacaine was injected into the joint after skin closure, before recovery from anesthesia. Foals received flunixin meglumine (1.1 mg/kg bwt i.v. or p.o. once daily) for 3 days and trimethoprim/sulphadiazine (5/25 mg/kg bwt p.o. twice daily) for 7 days postoperatively.
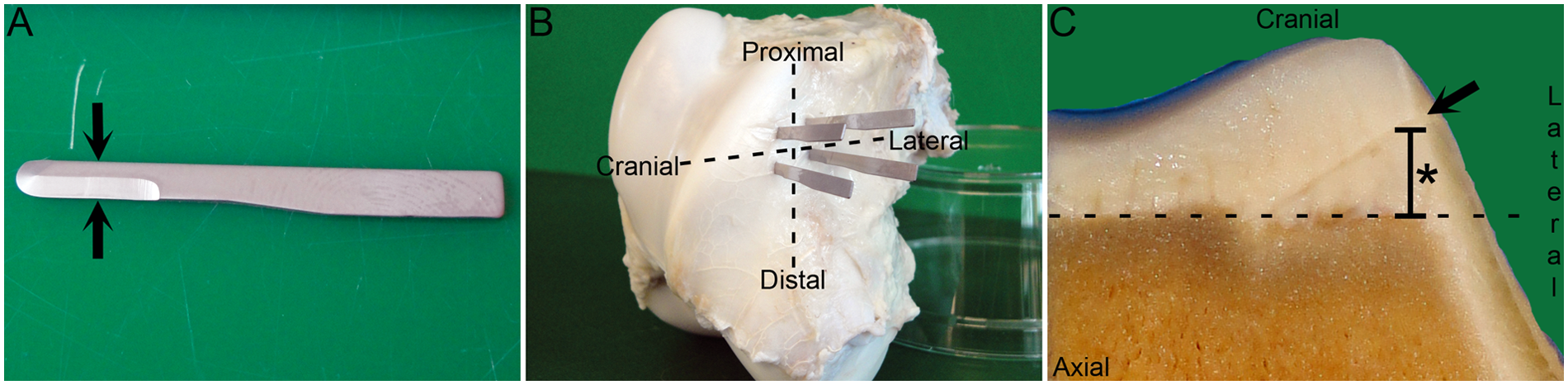
Methods. (
Follow-up Examination
Foals were arbitrarily assigned a time for follow-up examination from 1 to 49 days postoperatively. Each foal was euthanized with an overdose of pentobarbital. The operated left distal femur was harvested and fixed in 4% phosphate-buffered formaldehyde for 48 hours. The incised lateral trochlear ridge was sawed free from the femur with a band saw and decalcified in 10% EDTA. The decalcified block was cut into 5-mm thick slabs in a transverse plane parallel with the articular weight-bearing surface of the distal femoral condyles ( Fig. 1C ). The slabs were trimmed to fit into cassettes measuring 32 × 25 × 5 mm, which were then paraffin embedded, sectioned, and stained with hematoxylin and eosin for histological examination.
The number of animals and time to follow-up examination was based on previous experimental studies in piglets.19,20
Results
Exclusions did not occur. One foal was diagnosed with pneumonia on the first postoperative day and euthanized on humane grounds. The foal was included, representing the examination interval of 1 day postoperatively. The study group comprised 9 foals: 5 males and 4 females. Age at surgery was 13 to 15 days (mean = 13.6 days) and weight at surgery was 50 to 72 kg (mean = 63.3 kg). One foal was examined at each of the intervals of 1, 4, 10, 14, 21, 28, 35, 42, and 49 days postoperatively (weight = 63-114 kg).
Seven to 12 slabs were examined per foal (mean = 8.4). Four incision defects had been created in 9 foals, resulting in a maximum of 36 defects being available for examination. However, trimming caused a single lateral incision defect to be omitted from sections in 3 foals at 1, 42, and 49 days. Additional measures to section the omitted lateral incision defects were not taken. Thus, a total of 33 individual incision defects were examined histologically.
Changes are defined and described in the text at the first interval when they were observed and are summarized for all observation intervals in Table 1 . Descriptions refer to the proximal and distal cranial incision defects unless otherwise stated. The adjectives “superficial,” “intermediate,” and “deep” are used to indicate the depth level at which the described changes were located, both within the incision defect and within the articular–epiphyseal cartilage complex.
Local Morphological Changes at the Site of Articular–Epiphyseal Cartilage Stab Incision.
Observations in Histological Sections: Incision Defects
A portion of all incisions entered the subchondral bone and marrow cavities, however; because they were made with a round tip blade ( Fig. 1A ) in a curved region of the distal femur, incisions were not necessarily full thickness throughout their entire length of 6 mm.
Incision defects in the foal examined at 1 day were empty, or contained an acellular, eosinophilic material at all depths, possibly representing fibrin. Defects were located in a nonarticular region of the distal femur where the surface was covered by thin, sparsely cellular loose connective tissue. In the foal examined at 4 days, the superficial portion of the distal incision defect contained loose connective tissue that resembled the tissue on the bone surface. The deep portion of the proximal incision defect contained tissue that was identical to subchondral bone marrow.
Cells within incision defects of the foals examined at 10 and 14 days were more differentiated than cells in earlier examined foals ( Table 1 ). The superficial portion of incision defects contained capillaries and fibroblast-like cells ( Fig. 2A ). The deep portion contained cells interpreted as chondrocytes, in lacunae surrounded by a modest amount of matrix ( Fig. 2B ).
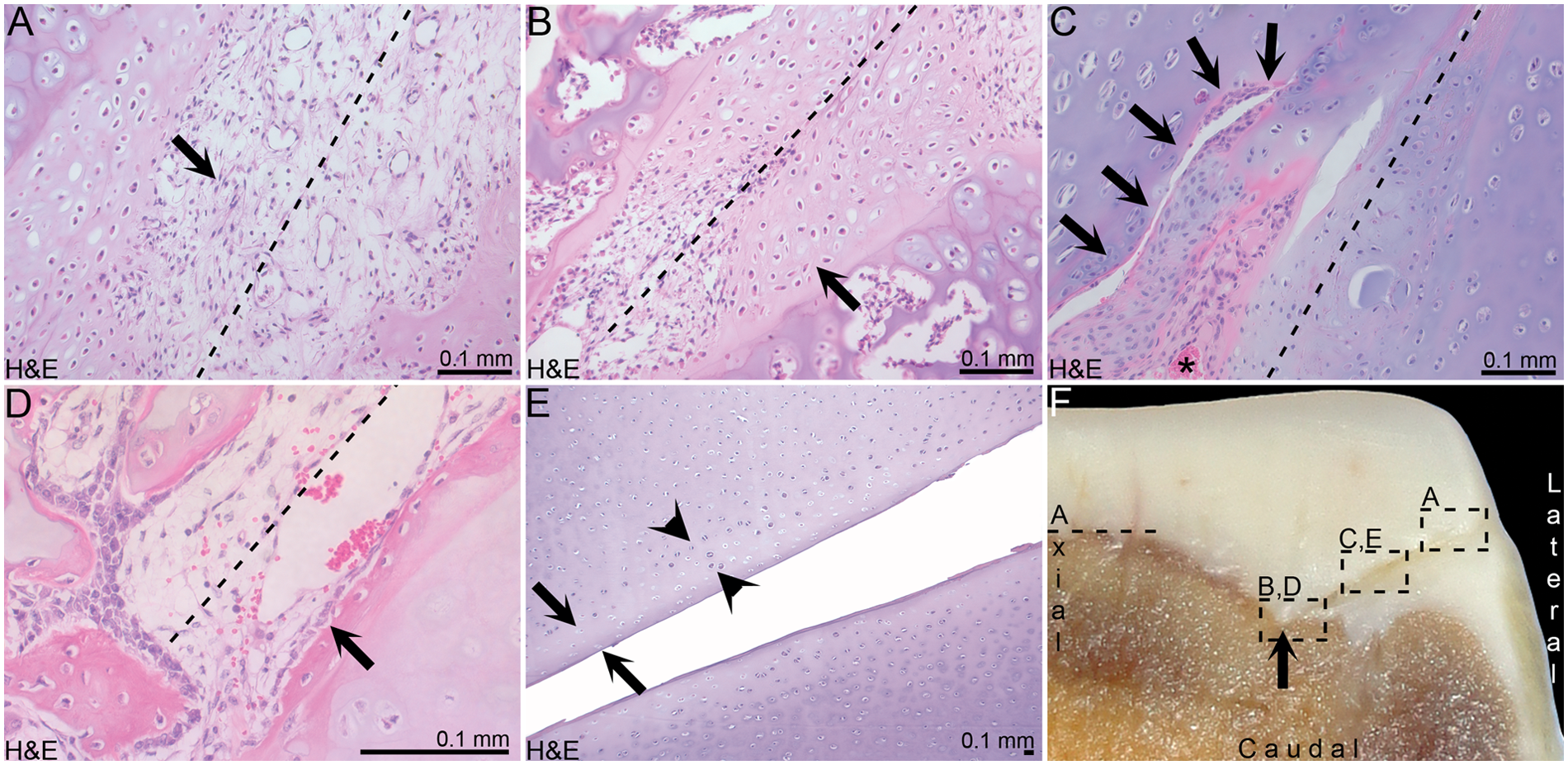
Results. (
Examples of incision defects traversing cartilage canal luminae were occasionally captured in sections from all foals. At points where incisions and luminae met at an intermediate depth of the epiphyseal growth cartilage, there was a continuum between cells within incision defects and perivascular cells within incised, but viable portions of cartilage canals ( Fig. 2C ). Perivascular cells within canals therefore contributed to the filling of incision defects with cells.
In the foal examined at 21 days, the deep portion of the proximal lateral incision defect contained palisading, osteoblast-like cells along its lateral margin ( Fig. 2D ). A narrow zone of matrix adjacent to these cells stained intensely eosinophilic and was interpreted as osteoid, based on its close similarity to osteoid in adjacent, nonincised tissue.
Cranial incision defects were oriented obliquely with respect to the underlying ossification front. In the earliest examined foal, incision defects were separated from the underlying ossification front on the cranial aspect of the lateral trochlear ridge by a maximum distance of 8 mm ( Fig. 1C ). This distance decreased with increasing post-operative time until, from 35 days, the superficial portion of incision defects was located at or deep to the cranial ossification front ( Table 1 ).
In the foal examined at 42 days, the subchondral bone contained 4 individual, linear areas of soft tissue, oriented either approximately parallel or perpendicular with respect to the cranial ossification front. The parallel areas were interpreted as granulation tissue within the deep portion of incision defects. The perpendicular areas were interpreted as granulation tissue within incised, but viable portions of cartilage canals. The granulation tissue was generally heterogeneous, but contained smaller foci of cells displaying homogeneous fibroblast-, chondrocyte-, or osteoblast-like characteristics within each focus. The granulation tissue also contained occasional multinucleated, giant cells, interpreted as clasts.
In the foal examined at 49 days, superficial portions of incision defects located within the cartilage on the lateral aspect of the lateral trochlear ridge were almost completely filled with acellular material. Ossification was progressing around portions of incision defects that were in contact with the ossification front, as described for the foal examined at 21 days.
Observations in Histological Sections: Cartilage on the Margins of Incision Defects
Changes on the margins of incision defects were uniform along the entire length of the defect unless otherwise indicated.
In the foals examined at 1 and 4 days, occasional chondrocytes within the cartilage on the margins of incision defects displayed cell shrinkage and cytoplasmic eosinophilia, interpreted as necrosis. Pyknotic nuclei were still visible within these cells. In the foals examined at 10 and 14 days, there was a continuous zone of chondrocyte necrosis within the cartilage on the margins of incision defects ( Fig. 2E ). Nuclear remnants were not visible in these cells, characterized as karyolysis, but eosinophilic cytoplasm remained, interpreted as coagulative necrosis. The zone of necrosis was an average of 2 cell layers thick, but a maximum 6 layers of necrotic chondrocytes were counted in exceptional, highly focal portions of the zone. The zone remained an average of 2 cell layers thick with increasing time ( Table 1 ). The matrix surrounding necrotic chondrocytes stained mildly paler than matrix surrounding morphologically normal chondrocytes ( Table 1 ).
In the foal examined at 4 days, occasional chondrocytes peripherally adjacent to necrotic chondrocytes showed proliferation, characterized by the presence of more than one chondrocyte within a lacuna, outside the proliferative zone of epiphyseal growth cartilage. In the foals examined at 10 and 14 days, proliferating chondrocytes formed a continuous zone adjacent to the zone of chondrocyte necrosis ( Fig. 2E ). The number of chondrocytes within individual lacunae tended to increase with increasing postoperative time ( Table 1 ).
Observations in Slabs
In the foals examined at 35 and 49 days, the proximal cranial incision defects were associated with a visible notch in the ossification front, representing a focal delay in enchondral ossification ( Fig. 2F ). In histological sections, ossification had progressed up to, but not beyond, the thin zone of chondrocyte necrosis adjacent to the incision defect.
Observations in Intact Bones
In the foal examined at 28 days, the distal incision defect was confluent with a cartilage fracture ( Fig. 3A ). The fracture margins were characterized histologically by a zone of chondrocyte necrosis that was 2 cell layers thick. The fracture was therefore interpreted as direct propagation of the incision defect.
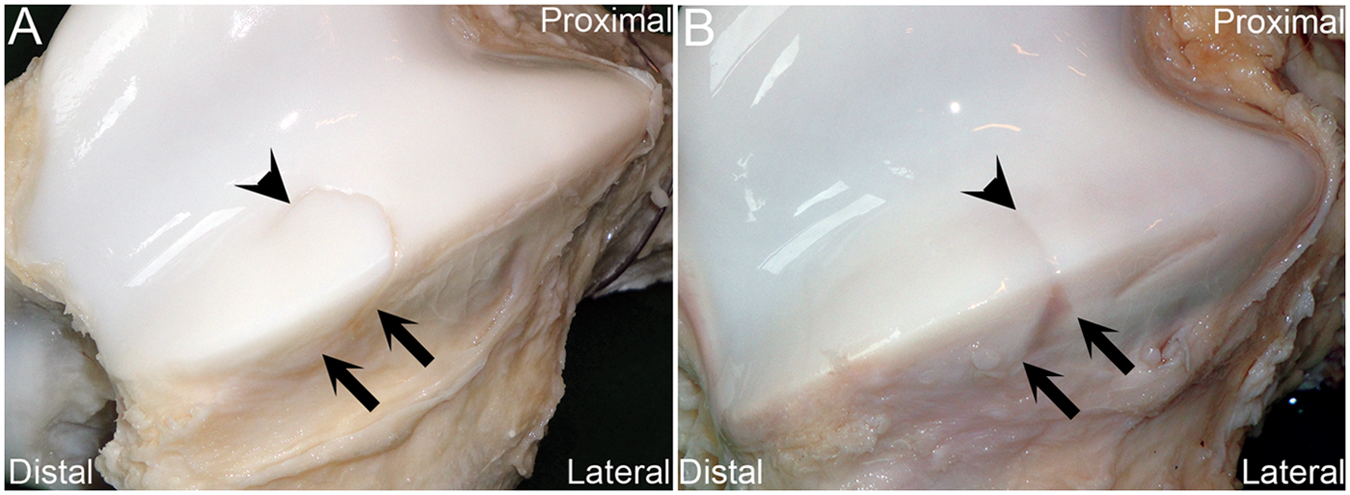
Intact bones. (
In the foal examined at 35 days, the distal incision defect was associated with a horizontal, linear depression in the cartilage surface on the cranial and lateral aspects of the lateral trochlear ridge ( Fig. 3B ). The cause of the depression was not clear in histological sections.
In the foal examined at 42 days, there was an irregular, vertical cartilage fracture on the axial aspect of the lateral trochlear ridge. In histological sections, the fracture space was empty. A major portion of the fracture traversed through an extensive area of cartilage canal and chondrocyte necrosis. It was therefore interpreted as pathological fracture through an area of ischemic chondronecrosis induced by, but occurring within the cartilage distant from the stab incision.
Discussion
The current report is not the first study of cartilage injury and repair in skeletally immature animals. It may, however, be the first in which a potential relationship between cartilage injury and the process of enchondral ossification has been studied systematically within the articular–epiphyseal cartilage complex. The results indicate that additional factors need to be considered in cartilage injury and repair in skeletally immature versus adult individuals.
Incision defects filled with amorphous material and cells. By comparing different individuals at increasing time intervals following the creation of defects, cells demonstrated an apparent ability to differentiate toward loose connective tissue, cartilage or bone. Further characterization of the observed material and cells will be pursued using immunohistochemical staining in the future. The observation that small cartilage defects fill with cells is in agreement with a number of previous studies. In these studies, the most frequent identified sources of cells are the subchondral bone marrow,21-25 the adjacent cartilage,22,23,26) and the synovium. 27 Defects were previously created in fetal ovine cartilage that was observed to contain patent cartilage canals, but the depth of the defects was controlled, and canals were not incised. 26 Calandruccio and Gilmer 23 created a variety of defects in the articular–epiphyseal cartilage complex of puppies and considered that “perivascular connective tissue related to intracartilaginous blood vessels might have contributed to repair.” The current rare observation that perivascular cells within incised, but viable portions of cartilage canals contributed to the filling of defects with cells therefore supports Calandruccio and Gilmer’s early suspicion that this might be the case. 23 Experimental defects are preferentially created within cartilage that is presumed normal, that is, free from macroscopically visible signs of local or generalized disease. During the drilling of stable juvenile osteochondritis dissecans lesions in children, tracts are created between normal and diseased tissue.28-30 Although the current defects were created in presumed normal tissue, the results can potentially be seen as a validation of the rationale behind the drilling of stable juvenile osteochondritis dissecans lesions, which is to induce repair by recruitment of stem cells into the diseased tissue.28-30
Stab incisions induced necrosis of a narrow zone of chondrocytes within the cartilage on the margins of all incision defects in the current examined young foals. Bupivacaine is toxic to equine adult articular chondrocytes in in vitro cell culture. 30 The observed chondrocyte necrosis was, however, of seemingly identical character and extent to the necrosis observed in previous experimental studies where bupivacaine was not used.22,23,25,32 Thus, the observed chondrocyte necrosis was unlikely to have been caused by bupivacaine toxicity. In previous studies, it was suggested that chondrocyte necrosis occurred because of mechanical forces,23,26,32,33 loss of a survival factor, or influx of an inhibitory substance.23,34 The current observed chondrocyte necrosis was morphologically compatible with coagulative necrosis, which is often associated with ischemia. 35 The ischemic chondronecrosis that is observed in equine articular OC is centered on necrotic vessels within zones of the epiphyseal growth cartilage that are not accessible by diffusion, typically comprises more than 250 cells and induces vigorous proliferation of vessels in adjacent, viable growth cartilage.36-38 In contrast, the current observed chondrocyte necrosis was centered on incision defects, was equally present in all cartilage zones, was an average of 2 cell layers thick, was not associated with vascular proliferation and therefore poorly compatible with necrosis caused by ischemia. The current results indicate that chondrocyte necrosis was unlikely to have been caused by either bupivacaine toxicity or ischemia, but the cause of the necrosis must ultimately be determined through studies that are designed specifically for this purpose.
In the current study, chondrocyte necrosis persisted and became associated with a change in matrix staining for the duration of the observation interval from 1 to 49 days after defect creation. In a previous study of OC lesions in pigs, similar altered matrix staining was demonstrated to reflect proteoglycan loss. 39 Sharp dissection is sometimes used in the preparation of grafts and recipient beds in arthroplasty procedures for the purpose of cartilage repair.40,41 Although major advances have been made in articular hyaline cartilage repair, the challenge of optimal, long-term integration of the repair tissue into the native cartilage remains. 25 Chondrocyte necrosis and altered matrix composition may impair, rather than enhance integration. Thus, the current results suggest that the use of sharp dissection may be contraindicated in cartilage repair procedures. A similar suggestion was previously made for the use of sutures to affix cartilage repair constructs, as suturing was observed to induce osteoarthritis-like changes. 33 The principal alternative is to consider biochemical rather than mechanical options, such as the use of glue instead of sutures to affix repair constructs 33 and the use of enzyme washing instead of sharp dissection to prepare defects for repair. 27
Stab incisions were associated with complications in three foals at 28, 35, and 49 days, respectively. A single cranial incision defect was associated with a linear depression in the cartilage surface of the foal examined at 35 days ( Fig. 3B ). Regrettably, the reason for the association could not be determined in histological sections, which in retrospect should have been prepared in a sagittal, rather than a transverse plane of section, as this would have demonstrated the association more clearly.
A single cranial stab incision defect was associated with a focal delay in enchondral ossification in each of the foals examined at 35 ( Fig. 2F ) and 49 days. Enchondral ossification usually takes place within intact epiphyseal growth cartilage, that is, cartilage without the cleavage planes created by the current stab incisions, and in some ways it was perhaps more surprising that ossification apparently was able to progress at the site of 31/33 studied incision defects, than that it was delayed in association with only 2/33 incision defects. Focal delays in enchondral ossification have, when representative of OC, been associated with clinically significant joint effusion and lameness even in the presence of an intact, overlying articular cartilage surface in horses. 36 The focal delay in enchondral ossification that occurs in equine OC is, however, caused by locally extensive areas of ischemic chondronecrosis. 37 The current observed chondrocyte necrosis was limited and approximately equal for all incision defects and therefore does not in itself explain why only 2/33 defects were associated with a delay in ossification. The location and orientation of the incision defects was tailored to the individual pattern of the blood supply, rather than standardized. Cranial incision defects were oriented obliquely with respect to the underlying ossification front. The current lateral defects and previous experimental defects in piglets were oriented perpendicular to the ossification front,19,20 and an associated focal delay in enchondral ossification was not observed. Thus, it may be that the orientation of the defect relative to the ossification front determines whether an associated focal delay in enchondral ossification will develop or not. It has been suggested that the incised region of the lateral trochlear ridge in foals forms by ossification of cone-shaped, microscopic subunit volumes of growth cartilage. 36 The current stab incisions may have cleaved the cartilage variably between or across conical subunit volumes, and this could further explain why a focal delay in enchondral ossification developed in association with only 2/33 incision defects. Similarly, a single cranial incision defect appeared to have propagated through adjacent, morphologically normal, nonincised tissue in the foal examined at 28 days ( Fig. 3A ). Again, this is different from the pathological fracture that occurs through an area of ischemic chondronecrosis in equine osteochondrosis dissecans.36,37 It may be that the propagation was due to a particularly unfortunate orientation of this, compared with that of other incision defects. The implication is that the potential for defects to cause a focal delay in enchondral ossification or cartilage fracture can be minimized by optimal alignment of the defect to the direction of ossification. A similar suggestion has previously been made for the sometimes unavoidable use of implants that traverse the physis in the fixation of Salter–Harris type fractures. 8
In the current study, the net effect of continued enchondral ossification was that the incision defects and associated cartilage changes became progressively surrounded by bone. Within bone, defects were associated with a vascularized granulation tissue that contained clasts with the potential to remove necrotic chondrocytes and abnormal matrix by phagocytosis. Foci of osteoblast-like cells were observed within the granulation tissue, and similar granulation tissue was previously observed to undergo transformation to bone by secondary, intramembranous ossification in naturally occurring lesions of equine tarsal OC. 42 In pony foals at the current stage of maturation, it is considered likely that uncomplicated defects would have undergone spontaneous healing to morphologically normal bone. Continued enchondral ossification therefore exerted a profoundly beneficial effect on the repair of surgical stab incisions into the articular–epiphyseal cartilage complex. Many of the challenges of adult articular cartilage repair are circumvented in the repair of immature, epiphyseal growth cartilage by the fact that the tissue is temporary and destined for complete eventual removal and replacement by subchondral bone.6,7
Experimental injury exerted a negative effect on the process of enchondral ossification by causing a focal delay in ossification in 2 foals, apparently related to the orientation of the respective incision defects. Continued enchondral ossification exerted a beneficial effect on repair, by causing defects to become surrounded by subchondral bone within which complete healing was likely to occur. The current observed relationship between cartilage injury, enchondral ossification, and repair warrants consideration in future studies of cartilage injury and repair within the articular–epiphyseal cartilage complex of all species.
Footnotes
Authors’ Note
The reported work was carried out at the Norwegian School of Veterinary Science.
Acknowledgments and Funding
This work was supported by the Norwegian Agricultural Purchasing and Marketing Co-Op and the Astri and Birger Torsted Foundation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The experimental protocol and animal use were approved by the National Animal Research Authority. Animals were kept in accordance with national animal welfare regulations.