
Abstract
Objective:
Little is known about how the biochemical properties of collagen change during tissue regeneration following cartilage damage. In the current study, temporal changes in cartilage repair tissue biochemistry were assessed in a rabbit osteochondral defect.
Design:
Bilateral full-thickness 3-mm osteochondral trochlear groove defects were created in 54 adult male skeletally mature New Zealand white rabbits, and tissue repair was monitored over 16 weeks. Collagen content, cross-links, lysyl hydroxylation, gene expression, histological grading, and Fourier transform infrared analyses were performed at 2, 4, 6, 8, 12, and 16 weeks.
Results:
Defect fill occurred at ~4 weeks postinjury; however, histological grading showed that the repair tissue never became normal, primarily due to the presence of fibrocartilage. Gene expression levels of Col1a1 and Col IIaI were higher in the defect compared with adjacent regions. Collagen content in the repair tissue reached the level of normal cartilage at 6 weeks, but it took 12 weeks for the extent of lysine hydroxylation to return to normal. Divalent immature cross-links markedly increased in the early stages of repair. Though the levels gradually diminished thereafter, they never returned to the normal levels. The mature cross-link, pyridinoline, gradually increased with time and nearly reached normal levels by week 16. Infrared imaging data of protein content paralleled the biochemical data. However, collagen maturity, a parameter previously shown to reflect collagen cross-link ratios in bone, did not correlate with the biochemical determination of cross-links in the repair tissue.
Conclusion:
Collagen biochemical data could provide markers for clinical monitoring in a healing defect.
Introduction
When articular cartilage is damaged as a result of osteoarthritis (OA) or injury, the tissue has limited capacity for repair and regeneration. There has been extensive research into techniques to repair damaged cartilage, including the use of microfracture techniques, autograft and allograft transplants, and autologous chondrocyte implantation procedures.1-3 However, a thorough evaluation of these strategies is limited in part by the lack of techniques for measuring the response of articular cartilage to tissue repair procedures. Knowledge of specific ultrastructural changes in early or later stages of repair could be extremely important during arthroscopic procedures, where crucial decisions are made regarding tissue quality. It is estimated that at least 65,000 such procedures are performed each year in the United States. 4
The progression and maturation of collagen cross-links are critical steps in the development and stability of connective tissues.5-7 Cross-link type is predominantly determined by the extent of hydroxylation of specific lysine residues catalyzed by lysyl hydroxylases (LHs),8-11 whereas cross-link quantity is determined primarily by the extent of oxidative deamination of the specific lysine or hydroxylysine residues in the telopeptides of collagen catalyzed by lysyl oxidase(s) (LOX), 12 respectively. There have been a limited number of studies that evaluated type II collagen cross-links in hyaline cartilage, primarily directed toward the goal of understanding how properties of the collagen network contribute to mechanical and functional properties in native and engineered cartilage13-19 but also assessing their contribution to MRI-derived cartilage parameters. 20
Collagen cross-links have also been evaluated in fibrocartilage, including in meniscus and intervertebral disc. Meniscal studies addressed whether cross-link changes were found with interventions, such as hyaluron treatment of a rabbit medial meniscectomy model 21 and a chicken model of strenuous exercise, where pyridinoline cross-links were greatly reduced with exercise. 22 The collagen cross-link profile of harvested human menisci with aging was also investigated, and both immature and mature cross-links were found to increase with age. 23 In studies of aging intervertebral discs, pyridinoline decreased and pentosidine increased with aging, 24 suggesting a correlation with collagen and disc degeneration. Kaapa et al. 25 found that mature collagen cross-links decreased in a porcine disc injury model.
However, to date, cross-link maturation in cartilage repair tissue has not been systematically evaluated. Based on the earlier studies of both hyaline and fibrocartilage, knowledge of the natural history of cross-link progression in repair tissue could provide biochemical markers for clinical monitoring in a healing defect.
Although surgical repair of cartilage defects is increasingly performed worldwide, “second-look biopsies” to assess repair tissue growth biochemically or histologically are not typically performed given the invasive nature of this procedure. Therefore, biochemical assessment of cross-links in cartilage defects is not possible in a clinical scenario. Vibrational spectroscopic techniques in the cartilage field are currently used predominantly as research tools, but advances are being made toward clinical use. The evaluation of cartilage degradation in harvested human osteoarthritic tissues was carried out in our laboratory by mid-infrared fiber optic probe, a modality that has the potential to be used arthroscopically.26,27 As infrared spectroscopy is sensitive to changes in collagen molecular structure,28,29 if changes in collagen cross-links during the cartilage repair process could be monitored spectroscopically, it may be possible to use a minimally invasive fiber optic probe for assessment in a clinical environment.
In the current study, we evaluated the extent of lysine hydroxylation in collagen and the profile and maturation pattern of collagen cross-links in a rabbit osteochondral defect model combined with histological grading of the tissue and gene expression studies. Spontaneous repair of osteochondral defects has been shown to occur in rabbits,30-32 lending support to the use of this model to follow the natural process of repair. In addition, we used Fourier transform infrared imaging spectroscopy (FT-IRIS) to assess correlations between spectral and biochemical assessment of cross-links to evaluate the potential for infrared assessment of cartilage repair tissue cross-links in situ.
Methods
Rabbit Osteochondral Defect Creation and Tissue Extraction
Bilateral full-thickness osteochondral trochlear groove defects 3 mm in diameter were created in 54 adult male skeletally mature, New Zealand white rabbits under IACUC approval. Animals (n = 9 per time point, 18 defects per time point) were sacrificed using barbiturate overdose at 2, 4, 6, 8, 12, and 16 weeks postdefect creation. At each time point, tissues were harvested for histology and FT-IRIS (n = 5), gene expression (n = 8), and collagen amino acid and cross-link analyses (n = 5). For histology, the defect and surrounding area were removed with a saw. For gene expression and collagen cross-link analyses, tissues were sampled from 3 regions: Region A, within the healing defect; Region B, which included cartilage around the healing defect (peri-lesion or adjacent cartilage); and Region C, control tissue harvested from cartilage on either of the femoral condyles, which was grossly normal in appearance at all time points. Initially, the decision was made to evaluate normal control tissue from the same limb to account for interanimal variability. However, gene expression in cartilage from mature rabbits that did not undergo any procedure was also added as a control. For all samples, care was taken to harvest only the cartilage and cartilage-like tissue, excluding any gross bone or calcified tissue. Samples for gene expression analysis were homogenized with a Polytron homogenizer 1200 E (Kinematica, Bohemia, NY) in 1 ml of Tri Reagent on ice and stored at −80 °C until RNA isolation. Samples for collagen cross-link analysis were frozen at -20 °C until ready for processing.
Histological Grading
Harvested samples were fixed in formalin, decalcified in EDTA, and paraffin embedded, and 7-µm sections were cut and stained with Alcian blue or hematoxylin and eosin (H&E). Using the O’Driscoll semiquantitative scoring system,33,34 two blinded observers scored one Alcian blue and one H&E-stained section based on the nature of the predominant tissue, structural characteristics, surface regularity, freedom from cellular changes of degeneration, freedom from degenerative changes in adjacent cartilage, and bonding to adjacent cartilage. The possible range of scores is 0 to 24, with 24 reflecting pristine hyaline cartilage. To assess consistency between scorers, a Kappa statistic for interobserver variability was calculated 35 and found to be 0.81, which is considered excellent agreement.
Gene Expression
Total RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA), and the first-strand cDNA was synthesized using iScript cDNA Synthesis kit (Bio-Rad Life Science, Hercules, CA). Real-time PCR was performed in triplicate using the rabbit-specific primers for glyceraldehydes-3-phosphate dehydrogenase (GAPDH) (used as a reference gene, as in other studies of rabbit cartilage degeneration and repair36,37), Col IaI, Col IIaI, aggrecan, and SOX9. The detailed forward and reverse primer sequences, and resultant products are summarized in
Table 1
. They were analyzed by the DNA Engine Opticon 2 Real Time PCR Detector (Bio-Rad, Hercules, CA). The mean fold changes in gene expression relative to GAPDH were calculated using the values obtained from Region C as a calibrator by means of 2−Δ
Sequence of Primers Used for Gene Expression Analysis and Resulting Polymerase Chain Reaction Product Size in Base Pairs
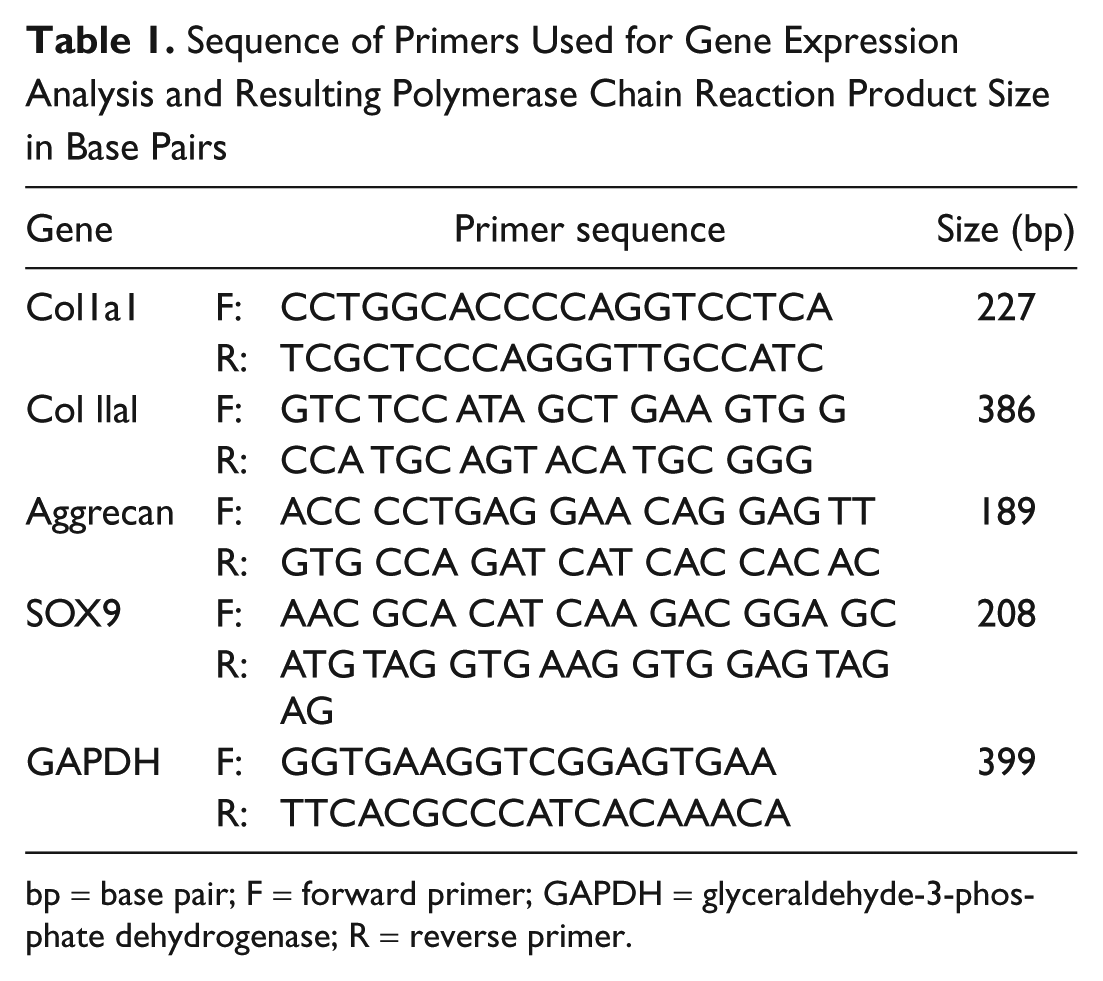
bp = base pair; F = forward primer; GAPDH = glyceraldehyde-3-phosphate dehydrogenase; R = reverse primer.
Amino Acid and Cross-Link Analyses
For amino acid analysis, tissues were collected from 3 regions in the same manner as described, pulverized to a fine powder under liquid N2 using a Spex Freezer Mill (Spex, Inc., Metuchen, NJ). Pulverized samples were washed with cold PBS and then with cold distilled water several times by centrifugation (4000g) and lyophilized. Five samples at each time point were then pooled together to obtain sufficient amounts of collagen for the following analyses. The analyses were done in duplicate to confirm the results. Dried samples were then reduced with standardized NaB 3 H4 and hydrolyzed with 6N HCl in vacuo at 110 °C for 24 hours, and an aliquot of each hydrolysate was subjected to amino acid analysis on a Varian high-performance liquid chromatography (HPLC) system (Prostar 240/310, Varian, Walnut Creek, CA) with a strong cation exchange column (AA-911, Transgenomic, San Jose, CA). The collagen content per protein was evaluated by the level of hydroxyproline (Hyp) per 1,000 total amino acids. The hydroxylysine (Hyl) content was calculated as moles per mole of collagen based on a value of 300 residues of Hyp per collagen molecule.
The hydrolysates with known amounts of Hyp were then analyzed for cross-links on a Varian HPLC with an AA911 column (see above) linked to a fluorescence detector (FP1520; Jasco Spectroscopic, Tokyo, Japan) and a liquid scintillation analyzer (500TR series; Packard Instrument, Meriden, CT) as described. 38 The major reducible cross-links (i.e., dehydrodihydroxylysinonorleucine/its ketoamine [deH-DHLNL], dehydrohydroxylysinonorleucine/its ketoamine [deH-HLNL]) were analyzed as their reduced forms, that is, dihydroxylysinonorleucine (DHLNL) and hydroxylysinonorleucine (HLNL), respectively. The nonreducible cross-link, that is, pyridinoline (Pyr) and deoxypyridinoline (d-Pyr), was also analyzed simultaneously as previously reported. 38 All cross-links were quantified as moles per mole of collagen.
Fourier Transform Infrared Imaging Spectroscopy (FT-IRIS)
Tissues from the paraffin histology blocks were sectioned at 7-µm thickness and mounted onto low-e slides (Kevley, Chesterfield, OH) for data collection in transflectance mode. FT-IRIS was performed using a Perkin Elmer Spectrum Spotlight 400 imaging spectrometer (Perkin Elmer, Shelton, CT). A rectangular region of interest that contained the defect and adjacent normal cartilage was selected, and data were acquired at a spatial resolution of 25 µm and spectral resolution of 8 cm−1.
FT-IRIS Analysis
Infrared data were analyzed using ISys 5.0 software (Malvern, UK). The spectral region from 900 to 2000 cm−1 contains molecular information associated with collagen and proteoglycans, the primary components of cartilage. 28 Images were masked to 3 different regions as repair tissue, adjacent to defect, and away from the defect. Quantitative data are presented for the defect region and tissue away from the defect region. Total protein content was monitored by the integrated area of the Amide I absorbance peak, centered at 1660 cm−1. The ratio of the 1660:1690 cm−1 intensities has previously been correlated to cross-link maturity in type I collagen in bone, 28 specifically to the ratio of Pyr to DHLNL, and was evaluated here to assess the potential correlation to mature and immature cross-links in repair tissue.
Statistical Methods
A paired Student t test was used to compare normal and repair tissue parametric values within each group. Significant differences among the repair tissue parameters for the 6 time points were determined by one-way ANOVA followed by a post hoc Tukey test to compare group by group differences, with significance at the P < 0.05 level. An ANOVA on ranks was used to assess significant differences in the O’Driscoll score among the 6 time points (SigmaStat 3.1, Systat Software Inc., San Jose, CA). Statistical analyses could not be performed on the biochemical data, as tissues from all 5 samples for each time point had to be pooled to obtain sufficient tissue for analysis. Thus, trends are reported for these data.
Results
Histology
Histological analysis revealed the defect was filled by the 6 or 8 week time point in all samples, and generally, an overall improvement in tissue quality was observed over the 16-week period ( Figure 1A - D ). However, the scores never reached the level of normal hyaline cartilage ( Figure 1E ). The nature of the predominant tissue was fibrous in ~28% of the samples, fibrocartilage in ~60% of the samples, and hyaline in only ~12% of the samples.
Gene Expression
Collagen II expression increased ~25-fold over time in the repair and adjacent tissue, peaking at approximately 4- to 6-fold over un-operated control between 8 and 12 weeks postoperatively (P < 0.05, all comparisons, Figure 2A). In contrast, the tissue away from the defect had collagen type II expression that remained at a constant low level through the time course. Collagen I expression in each of the tissues analyzed was present and remained constant throughout the 16 week period ( Figure 2B ). The aggrecan and SOX9 gene expression paralleled each other with increases over time in repair, adjacent, and tissue away from the defect. Compared with 2-week controls, expression levels rose approximately 4- to 6-fold peaking between 8 and 12 weeks postoperatively (P < 0.05, all comparisons, Figures 2C and 2D ); however, the absolute level of expression (normalized to GAPDH) were modest compared with the collagens.
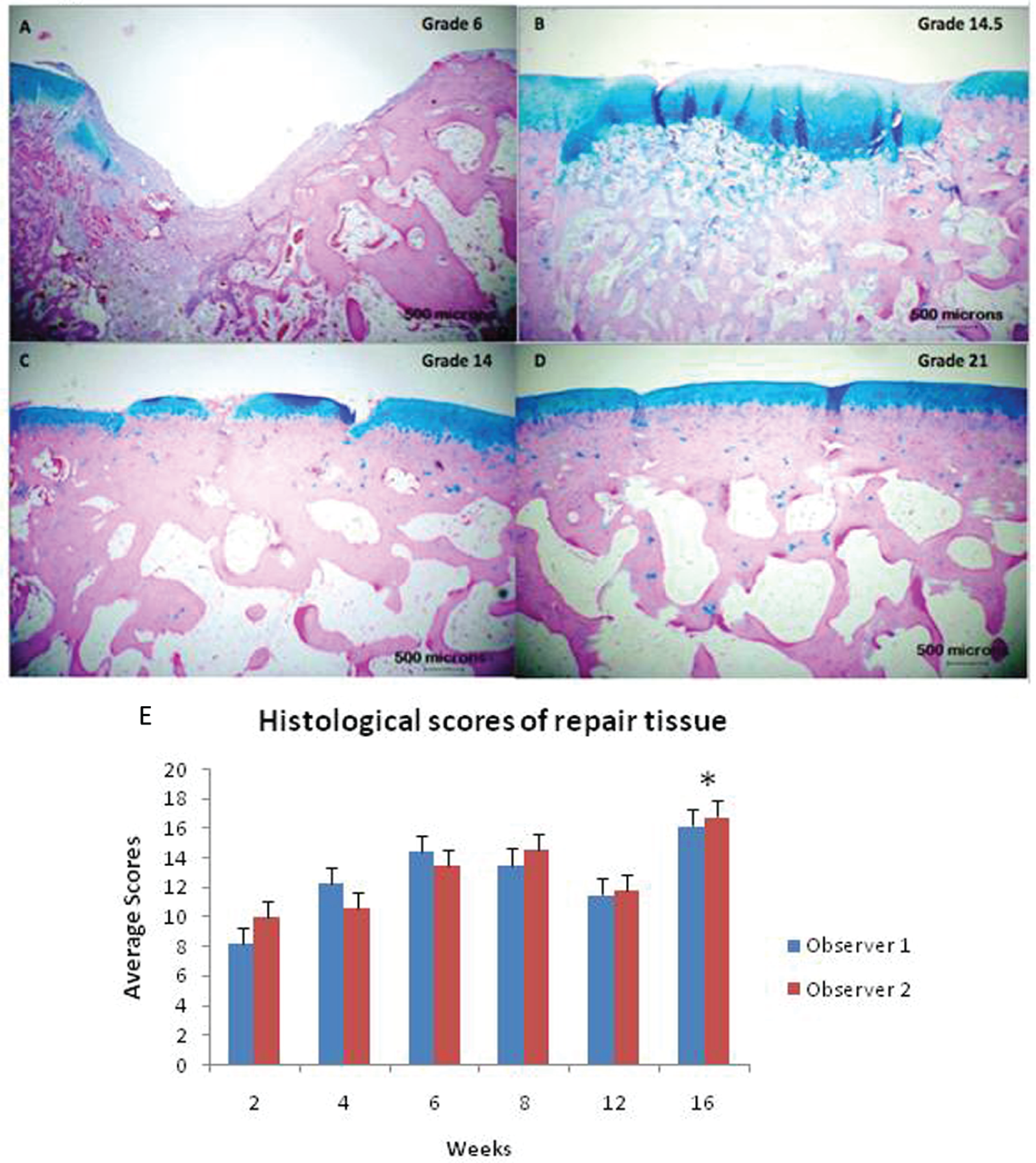
Alcian blue-stained histological sections of repair and adjacent cartilage at (
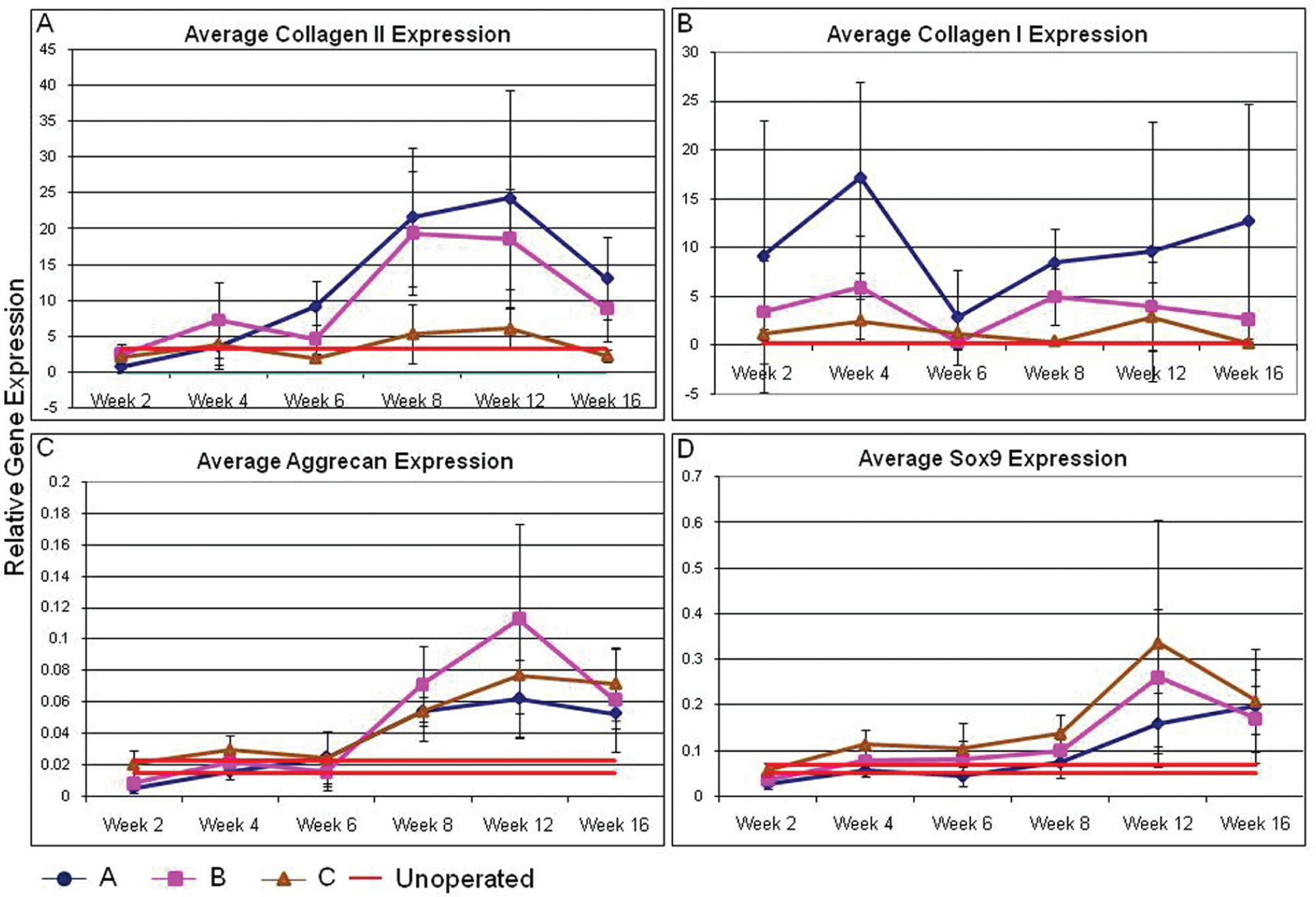
Gene expression levels of (
Collagen and Hyl Content
The collagen content (relative to total protein) in region A was low in the early stages (2-4 weeks) of healing but reached the control level at 6 weeks and remained normal thereafter ( Figure 3A ). The Hyl content in collagen in the region A was ~30% lower than those of region B and C at 2 weeks. However, the level gradually increased with time and reached near the control level at 16 weeks ( Figure 3B ).
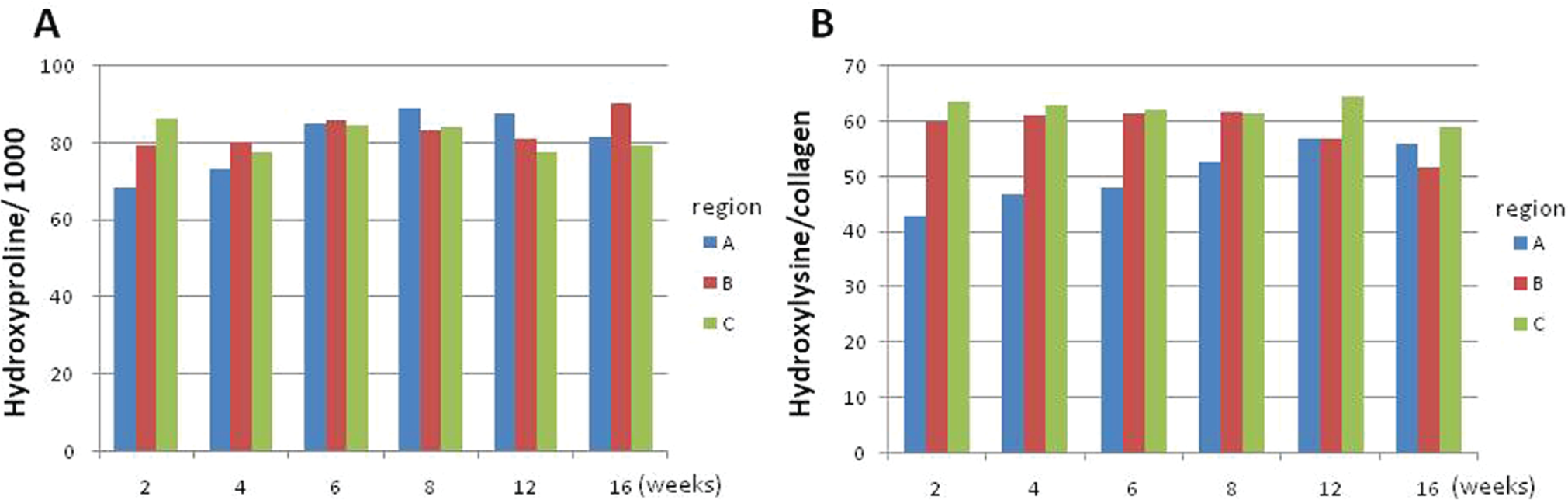
(
Collagen Cross-Link Analysis
Two reducible, immature cross-links, DHLNL and HLNL, and a nonreducible, mature cross-link, Pyr, were identified in all regions and the levels were compared among the regions ( Figure 4 ). No d-Pyr or histidine-based cross-links were detected in any of the regions at any time point. Cross-link precursor aldehydes were also below detection levels at any time point. In all 3 regions (A, B, and C), the major cross-links were DHLNL (immature) and Pyr (mature). Significant levels of reducible cross-links were found in region A during the course of experiment. Compared with region C, the level was 9.2-fold higher up to 6 weeks. Even at week 16, it was 4.7-fold higher than that of region C. In region A, DHLNL diminished from 0.6-0.7 mol/mol collagen in the early stages of healing (6 weeks) to 0.1 to 0.2 mol/mol collagen at late stage of healing (16 weeks). In region B and C, DHLNL stayed very low during the course of the experiment. HLNL, another reducible immature cross-link, was also identified as a minor cross-link in all of the samples. However, like DHLNL, the amount of this cross-link was always higher in region A (1.5- to 4-fold) compared with the other 2 regions. The levels of HLNL in the regions B and C were constantly very low (<0.03 mol/mol of collagen). In region A, Pyr was low in the early stages of healing (i.e., 2, 4 weeks); however, it increased from 0.2 mol/mol collagen at early stages to 0.5 to 0.6 mol/mol collagen at late stage. In region C, Pyr showed a slight increase at late stage.
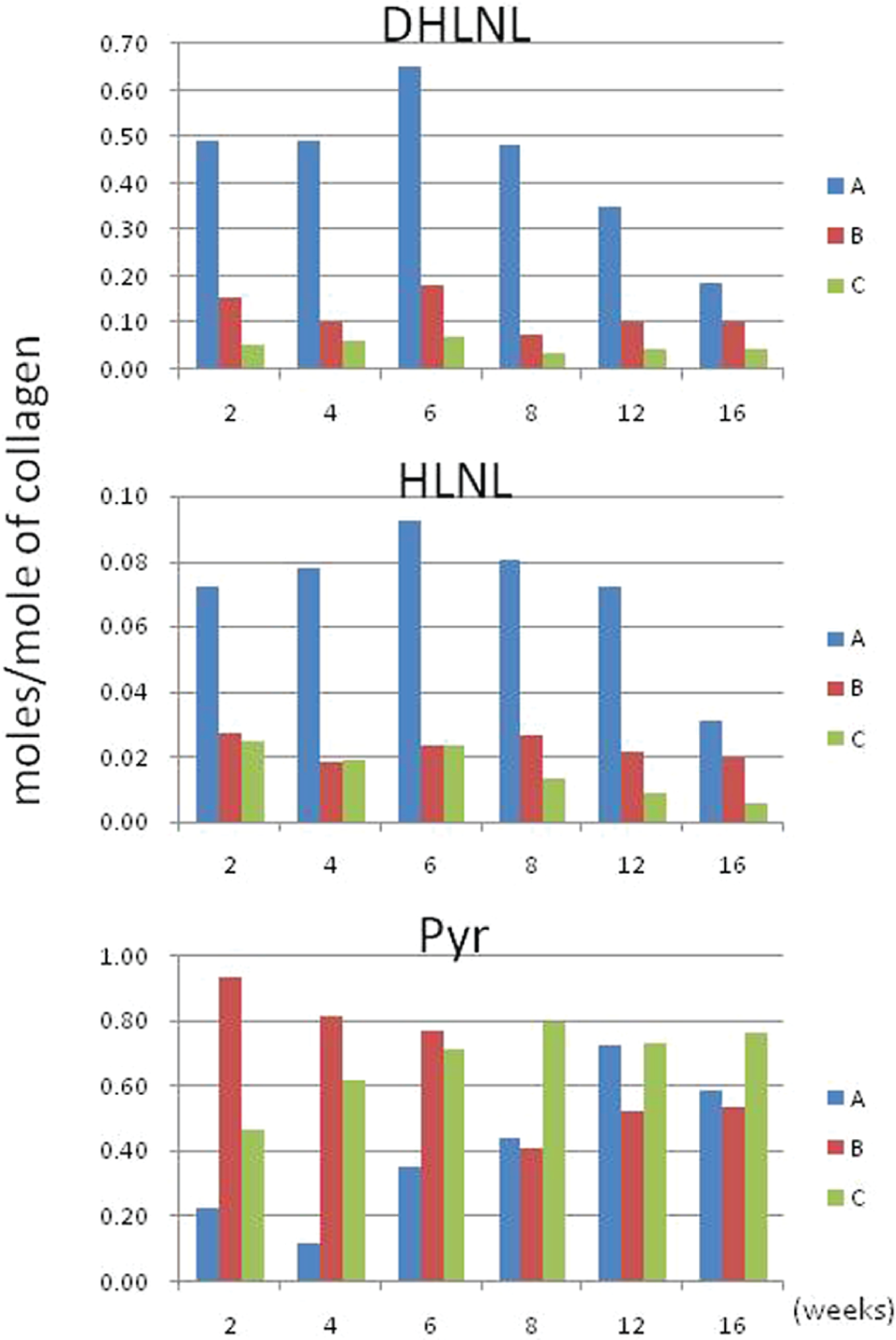
Collagen cross-links expressed as moles of cross-links per mole of collagen for all repair time points (2-16 weeks). DHLNL = dihydroxylysinonorleucine (immature cross-link); HLNL = hydroxylysinonorleucine (immature cross-link); Pyr = pyridinoline (mature cross-link). Region A, within the healing defect; Region B, which included cartilage around the healing defect (perilesion or adjacent cartilage); and Region C, control tissue harvested from cartilage on either of the femoral condyles, which was normal in appearance at all time points. Tissues from 5 samples per time point were pooled to obtain enough tissue for analysis, and therefore statistical analysis could not be performed. Immature cross-links were higher in repair tissue versus control and tended to diminish over time. Pyr was lower in repair compared with controls and appeared to normalize over time.
FT-IRIS
The repair tissue had a lower Fourier transform infrared (FTIR)-determined protein content compared with tissue away from the defect in the initial weeks post-injury but reached values comparable to normal tissue by 8 weeks ( Figure 5 ), consistent with the biochemical data. The previously reported infrared (IR) measure of collagen cross-link maturity, related to Pyr/DHLNL, was shown to be greater in the repair tissue in the initial weeks and then reached levels comparable to control tissue away from the defect by ~8 weeks, a result inconsistent with biochemical determination of cross-links.
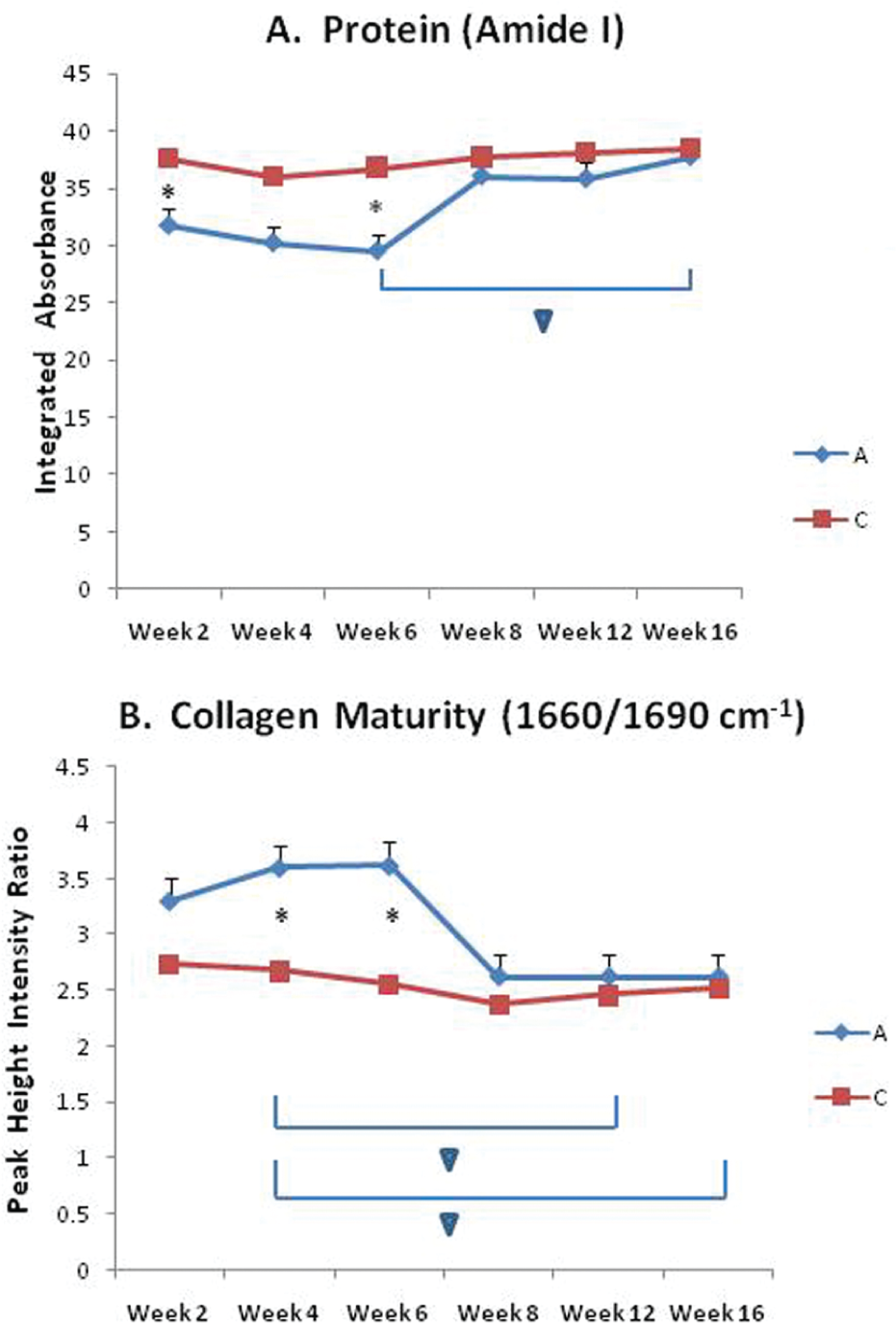
FT-IRIS-determined protein content (
Discussion
We have used a well-established rabbit osteochondral defect model to show the feasibility of quantifying specific measures of healing using collagen biochemical cross-link analysis and FTIR imaging with comparison to traditional measures of histological scoring and gene expression. As expected, the quality of the repair tissue in this rabbit model improved over the course of the 16-week period but did not reach that of normal, hyaline cartilage. This is in agreement with earlier studies in which normal hyaline cartilage was not observed even at 48 weeks postdefect creation in mature rabbits. 32 The collagen amino acid and cross-link analyses in the current study provided quantitative data to supplement the traditional repair measures and raised interesting questions.
Enzymatic cross-link formation is regulated by the action of 2 groups of enzymes: the lysyl hydroxylases (LHs) and lysyl oxidases (LOXs).8,9,12 In normal cartilage, Hyl aldehyde-derived cross-links, both immature and mature cross-links, are the predominant type of cross-link. It has been proposed that 2 residues of the bifunctional reducible cross-link, DHLNL, give rise to 1 tri-functional Pyr residue. 8 We found that although it takes 6 weeks to deposit a comparable amount of collagen matrix in the defects to controls, it took 12 weeks for the extent of Lys hydroxylation (posttranslational modification important for glycosylation and cross-linking) to reach the control level. This may explain the persistent presence of HLNL (a less hydroxylated cross-link) in Region A (defect) up to 12 weeks. Overall, the cross-link analysis indicates that it can take up to 16 weeks for the defects to become biochemically comparable to those of control.
Significant amounts of DHLNL and HLNL were present throughout the course of tissue healing in the defects, whereas their levels were constantly low in the control groups. Though they diminished at later stage of healing, both were still significantly higher than those of controls. Pyr, a mature cross-link, was initially low in the defect but increased during the repair period. These results indicate that divalent cross-links were rich in the repair tissue at early stage through active collagen synthesis and matured into a trivalent cross-link, forming mature collagen fibrils. Furthermore, Pyr in the peri-lesion of defects was high at the beginning but diminished during 2 to 8 weeks. The reason for this is not clear at this point, but it could be due to the anatomical location where differential mechanical properties are required. Pyr in the control also showed a slight increase with time. This is consistent with a previous report that demonstrated a greater Pyr content in articular cartilage associated with mechanical instability of the tissue. 39
There were some limitations in the biochemical analyses. Due to the small amounts of tissue, we had to pool the samples to perform biochemical analysis at each time point, and thus we could not evaluate the changes statistically. Furthermore, as the repair tissue was generally a combination of fibro and hyaline cartilage, it was not possible to discern total amount of type I versus type II collagen or cross-linking in type I versus type II collagen. In recent studies of human cartilage repair, Dickinson et al. 40 and Hollander et al. 41 developed a novel ELISA method to quantitatively assess type I and type II collagen in small tissue volumes from second-look biopsies, in addition to demonstrating the ability to quantitate mature and immature cross-links in these tissues. Although they did not explicitly differentiate cross-links in type I versus type II collagen from repair tissue, they did demonstrate differences in cross-link profiles (mature to immature ratio) of histologically classified fibrocartilage, with greater type I collagen content, compared with those of hyaline cartilage, with primarily type II collagen. Hyaline cartilage generally had cross-link ratios within the range for natural cartilage, and fibrocartilaginous tissues had a significantly lower ratio of mature/immature cross-links than natural cartilage or hyaline biopsies. Thus, we can draw a parallel to our data in the current study, where the biochemically derived cross-link ratios increased over the 16-week study period, as did the hyaline-like nature of the repair tissue. Therefore, in spite of the lack of explicit data on collagen type, the trend during the course of repair is clear, and the data obtained from monitoring collagen cross-linking provide insight into collagen phenotypic changes during healing. It suggests that this type of monitoring could become a useful biomarker to evaluate therapeutics in cartilage repair quantitatively.
The disparity in the biochemical cross-link data and the IR-derived cross-link maturity data were surprising, as we have previously shown a correlation between IR and chromatography analysis of cross-links in bone. 42 There are several possibilities that may explain the discrepancy: (1) FTIR-derived values likely reflect differences in type I as well as type II collagen and thus are influenced by type-specific sequence around the cross-links, whereas the biochemical data reflect changes without differentiating the two types. (2) FTIR data could be influenced by the presence of collagen-binding molecules that may mask the cross-linking sites of collagen, whereas biochemical data are not affected by such molecules. (3) FTIR data could also be influenced by the degree of glycosylation of the cross-links, whereas the biochemical analysis used in this study reflects the total values, that is, glycosylated and nonglycosylated forms. 43 (4) Finally, the tissues examined by FT-IRIS were formalin fixed, a process that cross-links tissues, and this could have affected results as well. Further studies are planned to establish the correlation between FTIR-derived parameters and biochemical cross-link data in cartilage repair tissue.
In summary, we have shown region- and time-related changes in tissue morphology and biochemistry during the spontaneous healing of osteochondral defects in a well-established rabbit model. This study demonstrated that the repair tissue lacked normal cartilage integrity and typical cross-linking patterns, despite the fact that it did fill the defect region. Accordingly, although the repair tissue may appear “hyaline” in some regions, it may be unable to support normal cartilage-like mechanical function. Thus, assessment of collagen biochemistry can provide important data to aid in understanding preclinical and clinical outcomes of tissue repair.
Footnotes
Acknowledgments and Funding
This work was supported by NIH R01 EB000744.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study was approved by our animal care committee.