
Abstract
Aging is a major risk factor for many chronic diseases, including cancer, cardiovascular disease, and diabetes. The basic causes of aging are not fully known; nonetheless, extensive research indicates that several key biological pathways play critical roles in controlling the process. These proteins include the sirtuin family, AMP-activated protein kinase (AMPK), insulin-like growth factor (IGF), autophagy processes, and nuclear factor erythroid 2-related factor 2 (Nrf2). Targeting these pathways with genetic therapies or dietary changes has been proven to increase longevity and slow aging-related decline. Seaweeds are high in nutrients, such as dietary fiber, polyunsaturated fatty acids, vitamins, minerals, and bioactive substances. They are known for their health benefits, such as antioxidant, anti-inflammatory, and anti-obesity activities. Notably, recent studies have indicated that extracts or isolated compounds like fucoidan, fucoxanthin, phlorotannin and carotenoids from seaweeds can positively influence these aging-regulating pathways. Certain seaweed-derived compounds have been shown to activate sirtuins, enhance AMPK activity, modulate IGF signaling, promote autophagy, and upregulate Nrf2 pathways, all of which are associated with extended lifespan and improved health span in various animal models. So, this review focuses on the potential anti-aging bioactive substances found in seaweeds, emphasizing studies conducted on cellular and animal models and their underlying anti-aging mechanisms.
Introduction to Cellular Senescence and Aging
Cellular senescence is a permanent state of cell cycle arrest that occurs when cells are subjected to various forms of stress. This process is characterized by the cessation of cellular proliferation, changes in cell morphology, alterations in gene expression, and the secretion of pro-inflammatory cytokines, growth factors, and proteases, collectively known as the senescence-associated secretory phenotype (SASP). Senescence serves as a critical mechanism to prevent the propagation of damaged cells, thereby acting as a tumor-suppressive measure. However, the accumulation of senescent cells over time contributes to tissue dysfunction and the progression of age-related diseases. 1
Aging is a complex, multifactorial process driven by various interconnected biological mechanisms and pathways. Key pathways involved in aging regulation include sirtuins, AMP-activated protein kinase (AMPK), insulin-like growth factor (IGF) pathway, autophagy, and nuclear factor erythroid 2-related factor 2 (Nrf2). Sirtuins are a family of NAD+ dependent deacetylases that regulate cellular processes such as metabolism, stress response, and aging, promoting longevity by enhancing DNA repair, reducing oxidative stress, and modulating mitochondrial function. 2 AMPK is an energy-sensing enzyme that maintains cellular energy homeostasis; its activation promotes autophagy, enhances mitochondrial biogenesis, and inhibits mTOR (mechanistic target of rapamycin) signaling, thereby extending lifespan. The IGF pathway, involved in growth and development, influences aging by modulating cellular growth, metabolism, and survival, with reduced IGF signaling being associated with increased lifespan and delayed aging. 3 Autophagy, a cellular degradation process, removes damaged organelles and proteins, maintaining cellular homeostasis and function, with enhanced autophagy linked to increased longevity and resistance to age-related diseases. Nrf2 is a transcription factor that regulates the expression of antioxidant proteins; its activation enhances cellular resistance to oxidative stress, mitigating aging and promoting longevity. 4
Aging significantly impacts health by increasing susceptibility to a variety of chronic diseases. The decline in cellular and tissue function associated with aging is a major risk factor for the development of cancer, cardiovascular disease, diabetes, and neurodegenerative diseases. 5 The accumulation of genetic mutations and a decline in immune surveillance contribute to increased cancer incidence with age. Age-related changes in the cardiovascular system, such as arterial stiffening and endothelial dysfunction, lead to a higher risk of hypertension, atherosclerosis, and heart failure. Impaired glucose metabolism and insulin resistance are more prevalent in older adults, increasing the risk of type 2 diabetes. 3 Additionally, aging is a major risk factor for neurodegenerative disorders such as Alzheimer's and Parkinson's disease, which are characterized by the accumulation of damaged proteins and neuronal loss. 6
Understanding the cellular and molecular mechanisms underlying aging and cellular senescence is crucial for developing therapeutic strategies to combat age-related diseases and promote healthy aging. This review will explore the potential of seaweed-derived bioactive compounds in modulating these aging-related pathways and their implications for extending lifespan and improving health span.
Seaweed Derived Bioactive Compounds with Anti-Aging Effects
Seaweed, also known as marine macroalgae, comprises a diverse group of photosynthetic organisms found in marine environments worldwide. Classified into three main types based on pigmentation brown algae (Phaeophyceae), red algae (Rhodophyta), and green algae (Chlorophyta). Brown algae like Laminaria (kelp), Macrocystis (giant kelp), and Sargassum are predominantly found in colder, temperate waters and often form extensive underwater forests. 7 They are typically found in intertidal and subtidal zones, attached to rocky substrates or floating freely in the water column. Brown algae are significant for coastal ecosystems as they provide habitat and food for various marine organisms. 8 They are also commercially valuable for their bioactive compounds and industrial uses. Red algae like Porphyra (nori), Gracilaria, Gelidium are characterized by their red to purplish coloration due to the presence of pigments like phycoerythrin and phycocyanin, which mask the green chlorophyl ll. They are typically found in deeper waters but can also thrive in intertidal zones. They are commonly found in tropical and subtropical regions, attached to rocks or other substrates. Green algae share characteristics with land plants, including chlorophylls a and b, which give them a green coloration. They can range in size from microscopic unicellular forms to large multicellular species 9 (Figure 1).

Seaweed Derived Bioactive Compounds with Anti-Aging Effects.
Fucoidan
Seaweed is renowned for its rich array of bioactive compounds that contribute to its nutritional, medicinal, and industrial value. Among these compounds, fucoidan stands out prominently. Primarily sourced from brown algae Phaeophyceae such as Fucus vesiculosus, Undaria pinnatifida (wakame), and various Sargassum species, fucoidan is a sulfated polysaccharide known for its diverse biological activities. 10 It exhibits potent antioxidant properties, scavenging free radicals and reducing oxidative stress. Moreover, fucoidan demonstrates anti-inflammatory effects by modulating inflammatory pathways and enhancing immune responses. These characteristics make fucoidan a promising candidate for various therapeutic applications, including cancer treatment, cardiovascular health promotion, and immune system support. In the cosmetic industry, fucoidan is utilized for its moisturizing and anti-aging properties, contributing to skincare products aimed at improving skin elasticity and reducing wrinkles.11-13
Phlorotannins
Another significant group of bioactive compounds found in seaweed is phlorotannins, exclusive to brown algae. Synthesized through the polymerization of phloroglucinol units, phlorotannins are potent antioxidants that surpass terrestrial polyphenols like catechins and flavonoids in radical-scavenging capacity.14,15 They are known for their anti-inflammatory properties, inhibiting inflammatory enzymes and reducing cytokine production. Additionally, phlorotannins exhibit antimicrobial activity against bacteria and fungi, suggesting potential applications in food preservation and pharmaceuticals. Their promising role in cancer prevention and treatment is under exploration, highlighting their potential as natural compounds for therapeutic interventions. 16
Carotenoids
Carotenoids, found in various seaweed types including brown algae (fucoxanthin), red algae (astaxanthin), and green algae (beta-carotene), contribute significantly to seaweed's bioactive profile. These pigments provide antioxidant protection against oxidative stress, enhancing cellular defense mechanisms and supporting overall health. 17 Fucoxanthin and astaxanthin offer photoprotective properties, shielding skin cells from UV-induced damage and contributing to skincare formulations aimed at sun protection and anti-aging effects. Carotenoids also exhibit anti-inflammatory properties, potentially benefiting inflammatory skin conditions and promoting immune function. Studies on fucoxanthin further explore its role in promoting fat metabolism and aiding in weight management, highlighting its potential as a functional ingredient in dietary supplements and nutritional products. 18
Polyphenols
Polyphenols, ubiquitous in all types of seaweed, encompass a diverse group of compounds with significant biological activities. These include antioxidant properties, which protect cells from oxidative damage and contribute to overall health maintenance. Seaweed polyphenols also possess anti-inflammatory effects, mitigating inflammation and supporting cardiovascular health. Their antimicrobial properties offer potential applications in food preservation and hygiene products.19,20 Polyphenols found in seaweed are increasingly studied for their therapeutic potential in chronic diseases such as cancer and metabolic disorders. In the food industry, these compounds serve as natural preservatives and enhancers of nutritional value, while in cosmetics, they contribute to skincare products aimed at anti-aging and skin protection. 21
Polysaccharides
Polysaccharides, such as agar, carrageenan, ulvan, and alginate, represent another vital group of bioactive compounds abundant in seaweed. These complex carbohydrates contribute to seaweed's gelling, thickening, and stabilizing properties, widely utilized in food, pharmaceuticals, and biotechnology. 19 Beyond their structural roles, seaweed polysaccharides exhibit diverse biological activities including antioxidant, immunomodulatory, and antiviral effects. They are integral to wound healing processes and are explored for their potential in drug delivery systems and tissue engineering. Polysaccharides like agar and carrageenan are essential in food products for their gelling properties, while ulvan and alginate find applications in biomedical research and industrial processes due to their biocompatibility and sustainability. 22
Mechanisms of Cellular Senescence and Aging
Cellular senescence is a state of irreversible growth arrest characterized by altered cellular morphology and function, playing a pivotal role in aging and age-related diseases. Understanding the mechanisms underlying cellular senescence is crucial for elucidating the biological processes driving aging phenotypes and identifying potential therapeutic interventions 23 (Figure 2).
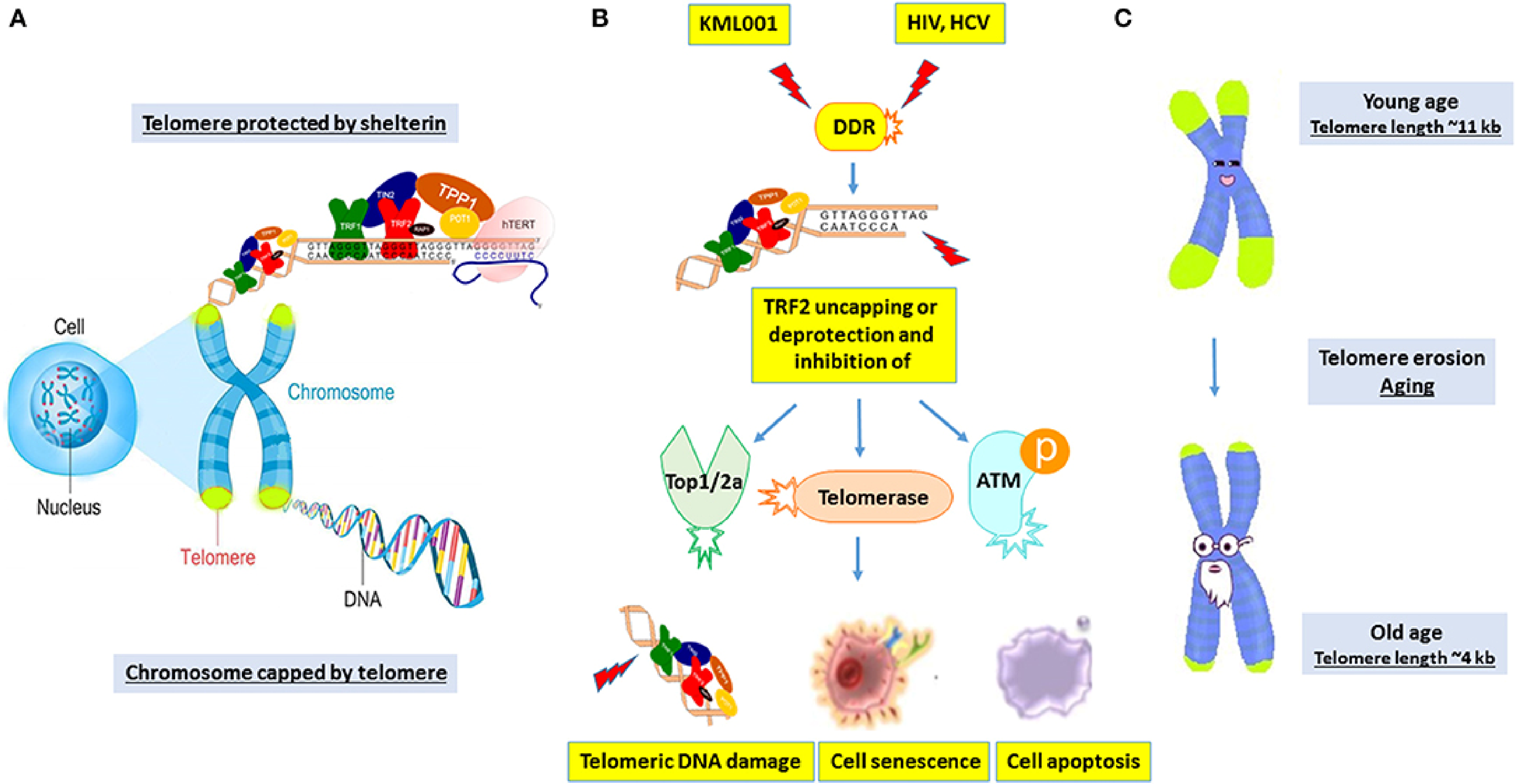
The Importance of Telomeres in Cellular Defense and Aging. (A) The Shelterin Complex Protects Telomeres, Which Are Found at the Ends of Chromosomes and Help Prevent DNA Damage. (B) TRF2 is Uncapped During Stress Conditions, Including as Exposure to DNA-Damaging Chemicals Like KML001 Or Viral Infections (HIV, HCV), Resulting in Telomeric DNA Damage, Senescence, or Apoptosis Via Telomerase Suppression and DNA Repair Pathway Activation. (C) Telomeres Shorten with Age, From Approximately 11 kb in Youth to 4 kb in Old Age, Causing Cellular Aging and Malfunction.

A Schematic Illustration of Cellular Senescence Processes and major Molecular Indicators. NOX2/4 (NADPH Oxidase 2/4) Produces Reactive Oxygen species (ROS), Resulting in Mitochondrial Malfunction (Q2) and Decreased Telomere Length. This Causes Overexpression of the Cell Cycle Inhibitors CDKN1A (Cyclin-Dependent Kinase Inhibitor 1A, p21) and CDKN2A (Cyclin-Dependent Kinase Inhibitor 2A, p16), Resulting in Cell Cycle Arrest. Increased β-Galactosidase (β-gal) Activity and Production of Senescence-Associated Substances (SA) Such as MMPs, Cytokines, and Chemokines are Also Observed. Inducible Nitric Oxide Synthase (iNOS) Increases Nitric Oxide (NO) Production, Exacerbating the Senescence-Associated Secretory Phenotype. Upregulation of the Urokinase Plasminogen Activator Receptor (uPAR) is Also Shown.
Telomere Shortening
Telomeres, protective DNA-protein complexes at chromosome ends, play a critical role in maintaining genomic stability. During cellular replication, telomeres progressively shorten due to the end replication problem and insufficient telomerase activity. Shortened telomeres trigger a DNA damage response, leading to cellular senescence or apoptosis. Studies have demonstrated that telomere dysfunction contributes significantly to replicative senescence and age-related pathologies24,25 (Figure 3).
Oxidative Stress
Reactive oxygen species (ROS), byproducts of cellular metabolism, induce oxidative damage to cellular components, including proteins, lipids, and DNA. Chronic oxidative stress overcome cellular antioxidant defenses, leading to persistent DNA damage and activation of senescence-associated pathways such as p53 and p16INK4a. Oxidative stress-mediated senescence is implicated in aging processes and age-related diseases, highlighting the role of redox homeostasis in cellular aging.26,27
DNA Damage
Accumulation of DNA damage, both from endogenous sources (replication errors, ROS) and exogenous insults (UV radiation, environmental toxins), triggers a DNA damage response.26,27 This response activates signaling pathways involving ATM, ATR, and p53, promoting cellular senescence or apoptosis to prevent the propagation of damaged DNA. Persistent DNA lesions and genomic instability in senescent cells contribute to aging phenotypes and age-related diseases.26,28,29
Epigenetic Alterations
Epigenetic modifications, including DNA methylation, histone modifications, and chromatin remodeling regulate gene expression patterns and cellular identity. 30 During aging, alterations in epigenetic marks disrupt normal gene regulation, contributing to cellular senescence and functional decline. Studies have implicated epigenetic dysregulation in the maintenance of senescent cell states and tissue aging, underscoring the role of chromatin remodeling complexes and histone modifying enzymes in aging processes.31,32
Inflammation and the Senescence-Associated Secretory Phenotype (SASP)
Senescent cells exhibit a senescence-associated secretory phenotype (SASP), characterized by the secretion of pro-inflammatory cytokines, chemokines, growth factors, and matrix metalloproteinases. 2 SASP components create a microenvironment of chronic low-grade inflammation, known as inflammaging, which contributes to tissue dysfunction and age-related pathologies such as cancer, cardiovascular diseases, and neurodegeneration. Understanding the regulation of SASP and its impact on neighboring cells is crucial for targeting senescent cell clearance and inflammation in aging interventions.2,33
Effects of Seaweed Derives on Lifespan Extension and Inhibition of Cellular Senescence
In animal models such as yeasts and flies, the lifetime extension effects of marine macroalgal chemicals have been reported. The carotenoid fucoxanthin, which is a significant photosynthetic pigment derived from brown algae, has been shown to extend the longevity of Caenorhabditis elegans and Drosophila melanogaster. A sulfated polysaccharide called porphyran is derived from the red algae Porphyra haitanensis and there have been reports of the lifespan extension effect of both natural and degraded porphyran in Drosophila. 34 Porphyrans have also been observed to boost the vitality of middle-aged flies in addition to their lifespan. These results imply that porphyran, at least in Drosophila, can lengthen life and improve health. In a similar way, polysaccharides derived from Saccharina japonica, a brown alga, have the ability to considerably increase the mean lifespan of Drosophila in both males and females. However, among females, the rise was more significant. 34
Conversely, an algal substance called fucoidan protects cells against stress-induced senescence as well as replicative senescence. According to some studies, fucoidan alleviated the senescence-induced senescence-associated beta-galactosidase (SA-β-Gal) activity in long-term cultivated endothelium colony-forming cells. Senescent cell identification is most commonly achieved with the use of the SA-β-Gal activity biomarker. 35 Additionally, fucoidan promotes the expression of the anti-senescence protein regucalcin while decreasing the expression of the pro-senescence protein p21. According to a different study, fucoidan therapy also lessens the SA-β-Gal activity in mesenchymal stem cells that is brought on by p-cresol, a significant uremic toxin. Furthermore, in stress-induced senescent fibroblasts, some chemicals derived from algae, such porphyran, demonstrated the anti-SA-β-Gal action. Furthermore, a class of red photosynthetic pigments found in red algae is called phycoerythrin. 36 The red alga Pyropia yezoensis synthesized phycoerythrin-derived peptide reduced the activity of SA-β-Gal in aged primary hippocampal neuron cells and lessened age-dependent neurite degeneration. 36
The anti-senescence capacity and lifespan extension effect of seaweed derivatives in lower organisms encourage studies on the anti-aging effect of algal compounds in higher animals. Although the effects of algal derivatives on major pathways that regulate lifespan have been reported, to the best of our knowledge, the lifespan extension effect of seaweed-derived bioactive compounds has not been evaluated using mammalian models. 35 This chapter investigates the anti-aging and regulatory effects of bioactive compounds detected in seaweeds using evidence from in vitro and in vivo studies. This chapter aims to explore the possible anti-aging advantages of these drugs and evaluate their application in future research. This study analyzes the regulatory roles of the anti-aging pathway using in vitro and in vivo studies on bioactive compounds derived from seaweeds. The goal is to understand the potential anti-aging impact of these compounds and explore their feasibility for future research.
Regulation of Anti-Aging Pathways by Seaweed Derived Bioactive Compounds
The SIRT (sirtuins), AMPK (5′ AMP-activated protein kinase), autophagy, and IGF (Insulin-like growth factor) signaling pathways have been identified as the main anti-aging-associated pathways. Other recognized regulators of aging include nuclear factor erythroid 2-related factor 2 (NRF2) and the antioxidant protein expression regulator. These routes are not, however, independent of one another. They typically collaborate in an integrative way, controlling the beginning or course of aging through mutual interactions.36,37
Activation of AMPK and SIRT Pathways by Seaweed Derived Bioactive Compounds
AMPK and SIRT Pathways
A class of protein deacetylases dependent on nicotinamide adenine dinucleotide (NAD+) is known as sirtuins (SIRT1–SIRT7). While SIRT2 is exclusively found in the cytoplasm, SIRT1 travels back and forth between the cytoplasm and the nucleus. While SIRT6 and SIRT7 are found in the nucleus and nucleolus, respectively, SIRT3, SIRT4, and SIRT5 are proteins found in the mitochondria. 38 Numerous transcription factors, histones, signaling molecules, and other enzymes are among their downstream deacetylation targets. The most closely related mammalian homologue to yeast SIR2, the most extensively researched sirtuin, is SIRT1. 37 In human fibroblasts, endogenous SIRT1 protein expression decreases with replicative aging. SIRT1 expression declined with ageing in the epidermis and kidneys of mice as well. Its additional genetic copies can replicate the effects of calorie restriction and have been demonstrated to increase the longevity of a variety of creatures, such as mice, flies, and yeasts. 38 Additionally, in yeasts and mice, SIRT1 small molecule activators like resveratrol and SRT1720 also demonstrate the ability to extend lifespans. SIRT1 substrates associated with cellular senescence include, but are not limited to, the autophagy negative regulator mammalian target of rapamycin (mTOR), the transcription factor forkhead box protein O (FOXO), which regulates cell development, and the tumor repressor p53. In addition to SIRT1, other sirtuins affect longevity and age-related illnesses. While resveratrol itself is not found in seaweeds, several seaweed-derived polyphenols and bioactive compounds, such as phlorotannins and fucoxanthin, have been shown to exhibit SIRT1-modulatory activity and similar anti-senescence effects. These compounds may mimic resveratrol's mechanism of action, positioning seaweeds as promising natural sources for anti-aging and cancer-preventive agents. 35 Mice with overexpressed SIRT6 have a much longer lifetime than wild-type mice. However, this lifespan extension effect was only seen in male mice. 35
In addition to SIRT, calorie restriction can activate AMPK, which has been shown to be crucial for the lifespan extension effect. Both SIRT1 and AMPK are affected by cellular energy status. Increased NAD+/NADH ratio activates SIRT1, whereas stressors that enhance the AMP/ATP ratio activate AMPK. SIRT1 partially inhibits senescence by activating AMPK via hepatic kinase B1. Simultaneously, AMPK promotes SIRT1 action by boosting cellular NAD + levels. 38 When AMPK is phosphorylated, its activity is at its highest. As a serine/threonine protein kinase, AMPK inhibits the synthesis of proteins and fatty acids while promoting the oxidation of fatty acids and glycolysis to produce ATP. It does this by phosphorylating specific metabolic enzymes and controlling the expression of relevant genes. Through an integrated signaling network, AMPK controls the aging process. It can improve cells tolerance to stress by activating signaling pathways like p53, Nrf2, and FOXO. 38
In the adipose tissue of older mice, the expression levels of both total AMPK and phospho-AMPK were lower than those of their younger counterparts. There is growing evidence that AMPK signaling becomes less sensitive as people age. 39 Numerous investigations have documented the lifespan extension function of AMPK. While lowered AMPK expression by RNA interference shortened the lifetime of Drosophila, transgenic expression of AMPK in adult muscle or fat bodies increased lifespan. Furthermore, mice with metformin, an AMPK-activating chemical, have longer lifespans and better health. More notably, male mice have shown similar health benefits when supplementing begins in middle life. 40
Adipose tissue ages more quickly in obese people. Genetically obese mice's adipose tissue displayed signs of early aging, including elevated p53 expression, proinflammatory cytokine secretion, and SA-β-Gal activity. In addition to persons with normal body weight, overweight subjects can also benefit from the anti-aging effects of the SIRT1 and AMPK pathways.39,40 In mice fed a high-fat diet, resveratrol administration increases the activity of both SIRT1 and AMPK, hence increasing the mice's survival. Overweight male individuals have also been shown to activate AMPK and SIRT1 in response to calorie restriction. 41
Seaweed-Based SIRT and AMPK Activators
The anti-aging properties of sirtuin-activating chemicals have been investigated. Resveratrol, a naturally occurring phenol found in berries and grapes, is the most well-known sirtuin-activating substance. Under a variety of circumstances, certain bioactive substances found in seaweeds have demonstrated the capacity to activate sirtuin. 42
The edible marine brown algae Ecklonia cava is found along the Korean and Japanese coasts. Its polyphenol extract has been shown to activate AMPK and SIRT1 in mice that have been made obese by high fat. The polyphenol extract supplementation dramatically raised the amount of hepatic phosphorylated AMPK and the expression of its downstream genes, while also mitigating the decline in hepatic SIRT1 protein level brought on by a high-fat diet. 43 According to a recent study, the methanol extract of Eucheuma cava, abundant in polyphenols, such as eckol, dieckol, and phloroglucinol derivatives, has shown significant biological activity. These phlorotannins are known for their potent antioxidant and anti-senescence properties, and have been reported to modulate cellular signaling pathways including SIRT1 activation, suppression of mTOR, and regulation of FOXO transcription factors, enhances AMPK phosphorylation in C2C12 mouse myoblasts after only one hour of treatment. Mice with streptozotocin-induced type I diabetes showed improved fasting glucose levels and recovered plasma insulin concentrations when they fed with methanol extract of E. cava. In mice fed a high-fat diet, it was discovered that an ethanol extract containing meroterpenoid-rich fraction from Sargassum serratifolium, another brown alga, raised the amount of phospho-AMPK in the liver. This fraction reduced obesity and non-alcoholic fatty liver disease brought on by a high-fat diet because it contains high quantities of sargahydroquinoic acid, sargachromenol, and sargaquinoic acid. 44
Additionally, conjunctival epithelial cells that have been treated with the brown alga A. nodosum extract have enhanced SIRT1 activity. Similarly, some red algae have demonstrated the ability to activate AMPK and/or SIRT1. For example, in differentiated 3T3-L1 mouse adipocytes, lipid formation was reduced by a phenol-rich extract of the red algae Gracilaria verrucosa. Additionally, during 3T3-L1 development, G. verrucosa extract enhanced glucose absorption and more significantly AMPK phosphorylation. Certain bioactive compounds from different algae such as phlorotannins (eg, dieckol), fucoxanthin, and algal sterols have also been found to demonstrate AMPK- and/or SIRT-activation capabilities, in addition to the ethanol and methanol extracts of algae, which contain a variety of diverse chemicals. 35
Phloroglucinol and dieckol, two substances that were isolated from E. cava, have demonstrated a notable capacity to increase AMPK. Cellular phospho-AMPK levels rose when immortalized human hepatocytes HepG2 were treated with phloroglucinol, a significant phenolic component in E. cava. Phloroglucinol given orally also markedly enhanced glucose tolerance in male mice given a high-fat diet.35,45 The brown algae Ecklonia stolonifera and E. cava contain the phlorotannin dieckol. Dieckol treatment of 3T3-L1 adipocytes during differentiation demonstrated the potential to activate AMPK, which resulted in the suppression of adipogenesis. Dieckol has also been shown to have an AMPK-activating action in db/db mice with type II diabetes. After 14 days of intraperitoneal dosing, the muscle tissues of the dieckol-administered group had a higher degree of AMPK phosphorylation than those of the saline-treated control group. Furthermore, their body weight, serum insulin level, and blood glucose level were all considerably lower than those of the control group.35,45
In addition to E. cava, bioactive substances from other brown algae can also activate SIRT and/or AMPK. For example, in db/db mice with type II diabetes, octaphlorethol A, another phlorotannin that was isolated from the brown algae Ishige foliacea, demonstrated effects on muscle AMPK-activation. Mice with diabetes who received an intraperitoneal injection of octaphlorethol A for two weeks experienced a reduction in hyperglycemia. 46 It dramatically lowered the postprandial blood glucose level, which continued to do so until week five, when the research came to an end. Two indole derivatives, indole-2-carboxaldehyde and indole-5-6-carboxyaldehyde, that were isolated from Sargassum thunbergii, another type of brown alga, demonstrated AMPK-phosphorylation induction effect in differentiated 3T3-L1 adipocytes in a dose-dependent manner, according to another study. It has been suggested that their suppression of adipogenesis and lipid buildup triggers the AMPK signaling pathway. 46
It has been shown that the sterol metabolite fucosterol, which was isolated from Ecklonia stolonifera, increases SIRT1 expression and inhibits FOXO1 phosphorylation in 3T3-L1. Reduced expression of phospho-FOXO1 suggested that SIRT1 was deacetylating it. Numerous types of brown algae are the primary source of fucoidan, which is primarily composed of fucose. Fucoidan can range in size from more than 100 kDa to less than 500 Da.47,48 Fucoidan shown the capacity to raise nuclear SIRT protein levels and phospho-AMPK in streptozotocin-treated β cells in addition to its influence on phospho-AMPK induction in human hepatoma cells. An SIRT1 inhibitor has been shown to limit fucoidan's ability to improve insulin synthesis and pancreatic β cell death caused by streptozotocin, suggesting that it works in a SIRT1-dependent manner. Fucoidan demonstrated strong SIRT6 activation effects in addition to SIRT1 activation effects. Since SIRT6 has also been linked to the control of cellular senescence, it would be interesting to look into fucoidan's anti-aging properties.47,48
In db/db mice with diabetes, it was shown that low-molecular-weight fucoidan (LMWF), derived from Saccharina japonica and has a molecular weight of 6500 Da, shielded the liver from harm. Hepatic triglycerides, cholesterol, and liver dysfunction indicators including plasma alanine aminotransferase and aspartate aminotransferase were reduced when LMWF was administered. Hepatic phospho-AMPK and SIRT1 levels were lower in db/db mice than in control C57BL-C mice. 49 Adding LWMF to db/db mice can raise their SIRT1 and phospho-AMPK levels. More significantly, the protective effect against inflammation and oxidative stress in HepG2 hepatic cells was nearly eliminated using an AMPK or SIRT1 inhibitor. These results imply that LMWF uses the SIRT1/AMPK pathways to shield the liver from harm. 49
Another study also reported on the influence of LMWF on AMPK activation. In the skeletal muscle of db/db diabetic mice, low-molecular-weight fucoidan produced by acid hydrolyzing fucoidan isolated from Undaria pinnatifida can raise the phospho-AMPK level.35,49 Additionally, it promoted phospho-AMPK expression in L6 myotubes, which resulted in the oxidation of fatty acids and the intake of glucose. LMWF is involved in the activation of SIRT3, a mitochondrial sirtuin, in addition to SIRT1. The long-term neurobehavioral results of treating aged mice with traumatic brain damage (TBI) with LMWF were improved. Compared to their younger counterparts, normal aged mice showed a drop in both the mRNA and protein levels of neuronal SIRT3. LMWF treatment increased SIRT3 expression in elderly mice following TBI. The neuroprotective effect of LMWF was partially inhibited by intracerebroventricular injection of siRNA to knockdown SIRT3.35,49
Another algal component with known nutraceutical benefits against cancer and obesity is fucoxanthin. In FL83B cells, which are oleic acid-induced hepatocytes, it can raise SIRT1 expression and AMPK phosphorylation. Fucoxanthin appears to modulate the AMPK signaling system in the skeletal muscle and liver of db/db mice with type II diabetes, as evidenced by the increased phosphorylation of AMPK in both tissues. 50 This may help to explain the health benefits of fucoxanthin in these db/db mice, which include a reduction in insulin resistance and body weight growth, as well as a drop in fasting glucose and epididymal fat weight. Another study found that fucoxanthin phosphorylated AMPK in HepG2 cells in response to oxidative stress triggered by iron and arachidonic acid, thereby reducing oxidative damage. 50
These findings point to the potential anti-aging effects of seaweed bioactive substances at the cellular and tissue levels. The majority of animal research on the SIRT or AMPK pathway and bioactive substances found in seaweeds has been done on obese or diabetic animals. It is unclear, particularly in animals, if these seaweed derivatives can increase the baseline levels of SIRT and phospho-AMPK expression as people age. Therefore, more research is required to determine whether bioactive substances like fucoidan and fucoxanthin, which are derived from seaweed, can be used to prolong the longevity of higher species like mice and primates.
The Bioactive Compounds from Seaweeds Initiate Autophagy
Aging Autophagy
Autophagy comes in three primary forms: chaperone-mediated autophagy, microautophagy, and macroautophagy. The primary form that will be covered in this article is macroautophagy, also known as autophagy. Autophagy is crucial for maintaining cellular homeostasis because it makes it easier for intracellular components that are defective or superfluous to be broken down and recycled. The phagophore's engulfment of material that needs to be broken down starts the autophagy process.51,52 After maturing into an autophagosome, this double-membrane structure elongates and sequesters the designated cellular components. It then moves through the cytoplasm to the lysosome and merges with it to produce an autolysosome. The lysosomal enzymes inside the autolysosome break down the inner membrane and its contents. Autophagy-related (ATG) proteins are essential for the development of autophagy machinery. For instance, ATG7 and ATG3 bind the lipid phosphatidylethanolamine to microtubule-associated protein 1 light chain 3 (LC3-I/ATG8) to create LC3-II/ATG8-PE. The converted LC3-II is therefore crucial for autophagosome biogenesis since it encourages the autophagosome's development and extension. Another autophagy effector is Beclin 1 (ATG6), which forms a complex with other elements to aid in the production of autophagosomes. In addition to ATG proteins, phosphorylation can activate mTOR, a crucial autophagy inhibitor.51,52
Autophagy becomes inadequate throughout both normal and pathological aging due to either a decreased autophagic flow or an excess of cargo brought on by long-term cellular damage. Cellular senescence may be triggered by the buildup of malfunctioning cellular components. Research has demonstrated that genetic suppression or disruption of autophagy shortens the lifespan of flies and causes age-related degenerative alterations in mammalian tissues. 51 Conversely, increased autophagy can postpone aging. Autophagy can be induced by calorie restriction; in mice, Drosophila melanogaster, and Caenorhabditis elegans, for example, autophagy suppression inhibits the anti-aging benefit of calorie restriction. Rapamycin is an autophagy inducer that can prolong the life of flies and mice when given over an extended period. Furthermore, overexpression of the autophagy gene LC3 in adult fly neurons has been linked to an increase in lifespan.35,38,51
The SIRT and AMPK pathways are not independent of the autophagy process. By blocking the mammalian target of rapamycin complex 1, a negative regulator of autophagy, the AMPK protein, a crucial upstream regulator of autophagy, promotes the autophagic processes. Autophagy is also necessary for the lifespan-prolonging impact of SIRT1, since the deletion or knockdown of Atg genes eliminates this effect.35,38,51
Autophagy Activators from Seaweeds
Numerous bioactive substances found in seaweeds have been investigated for their effects on autophagy. Nevertheless, research on how various substances either induce or inhibit autophagy has produced contradictory findings. Different experiment circumstances are probably the cause of the discrepancies. Study participants may react differently under stress than they would in a typical setting.53,54 Curiously, the majority of research has employed particular seaweed components rather than unrefined extracts.
The up-regulated expression of Beclin-1 and LC3 in gastric cancer SGC7901 cells suggests that the algal carotenoid fucoxanthin triggered autophagy. Fucoxanthin reduced the amount of mTOR and dose-dependently elevated the expression of LC3-II and Beclin-1 in human epithelial cervical carcinoma Hela cells. By promoting autophagy, as demonstrated by elevated expression of LC3-II and Beclin-1, fucoxanthin also demonstrated neuroprotection in a mouse model of TBI.53,54 Furthermore, fucoxanthin reduces mTOR phosphorylation and raises LC3-II and Beclin-1 levels in oxidatively stressed hepatocytes. The development of autophagosomes in DHA- or EPA-treated lung adenocarcinoma A549 cells suggests that other algal-derived substances, such as EPA and DHA, are also strong autophagy inducers. This review has already covered fucoidan's effects on AMPK and SIRT activation. Fucoidan's impact on autophagy has also been investigated. Human multiple myeloma U266 cells treated with fucoidan had more autophagosomes. In cells treated with fucoidan, phospho-mTOR expression dropped whereas LC3-II and Beclin-1 expression rose.53,54
In contrast, fucoidan has been shown to have an anti-autophagy impact on the liver in several clinical circumstances, including ischemia-reperfusion (I-R) and hepatic fibrosis. Fucosterol, another algal component, has also been shown to suppress autophagy in acutely wounded livers. By preventing autophagy, fucosterol pretreatment reduced hepatic damage in response to concanavalin A-induced acute liver damage. It's interesting to note that fucosterol had no effect on autophagy in healthy animals that did not have acute liver damage. 35
Because autophagy plays a role in liver illnesses, these anti-autophagy effects may not be entirely surprising. Liver fibrosis can be lessened by inhibiting autophagy since it is necessary for the activation of hepatic stellate cells, which generate an extracellular matrix in the liver during fibrosis. The function of liver autophagy in hepatic I-R is still unclear because it shows both positive and negative effects after hepatic I-R. Overall, it is more difficult to investigate and evaluate the effects of algal chemicals like fucoidan under normal circumstances due to the role of autophagy in liver disorders.35,46,53,54
Seaweed Bioactive Compounds Inhibit the Insulin/IGF-1 Signaling Pathway
The Pathway of Insulin/IGF-1
The pancreas secretes the hormone insulin, which controls blood glucose levels. A hormone that shares structural similarities with insulin is called insulin-like growth factor 1 (IGF-1). It has anabolic effects in adults and is crucial for childhood growth. In contrast to the AMPK, SIRT, and autophagy pathways discussed before, the increased insulin/IGF-1 pathway is a contributing factor to aging in a number of animals. Excessive insulin signaling speeds up aging and harms cellular function. In a number of model organisms, such as worms, flies, and mice, the insulin/IGF signaling system influences lifespan. Mice with a fat cell-specific homozygous deletion of the insulin receptor live longer in both sexes. 38
It has been determined that the primary receptors for insulin and IGF-1 in mammals are the insulin receptor (IR) and the IGF-1 receptor (IGF-1R), respectively. Insulin and IGF-1, the receptors’ ligands, activate them. Their intrinsic tyrosine kinase activity is triggered by ligand interaction, which causes receptor autophosphorylation and activation. The signal is transduced by the active receptors, which then initiate a series of signaling cascades that phosphorylate and retain the FOXO transcription factors in the cytoplasm. 55 FOXOs are essential for the insulin/IGF-1 signaling pathway's longevity regulating function. Additionally, FOXO promotes the transcription of genes involved in cellular differentiation, growth, metabolism, and stress resistance. Several ethnic groups have demonstrated a strong correlation between human longevity and genetic variations of FOXO. When insulin/IGF-1 signaling is suppressed, active FOXOs may have a lifespan-extending impact. 55
Humans have demonstrated the lifetime extension impact of inhibited insulin/IGF-1 signaling. However, insulin/IGF-1 signaling has a more significant sex difference in humans than other important lifespan extension pathways covered here. According to polymorphism research, women who have genetic variations that result in decreased insulin/IGF-1 signaling activity are more likely to survive into old life than men. Female participants in a different study of nonagenarians who had IGF-1 levels below the median had a noticeably longer survival time than those whose levels were above the median. Males, however, did not exhibit this survival benefit.35,55
Seaweed-Based Insulin/IGF-1 Inhibitors
Both isolated compounds and algae crude extracts of the insulin/IGF-1 signaling suppressors and FOXO activators have been employed. The green algae's polysaccharide that dissolves in hot water in AGS human gastric cancer cells, Capsosiphon fulvescens markedly reduced the phosphorylation of insulin receptor substrate 1 and IGF-1R in response to IGF-1. One important target of the insulin receptor is insulin receptor substrate 1, which the activated insulin receptor can phosphorylate. Additionally, the phosphorylation of Akt, an insulin and IGF-1 downstream target, was suppressed. 56
On the other hand, LMWF demonstrated the capacity to induce IGF-1 in mice receiving chemotherapy for bladder cancer. In comparison to mice given chemotherapy alone, mice treated with LMWF and chemotherapy medications showed increased IGF-1 expression and formation as well as decreased FOXO3 expression and activation. Since the mice in this study were in extremely pathological conditions, more research is necessary to determine how fucoidan affects IGF-1/insulin under normal conditions, much like autophagy regulation does in pathologic liver. 57
Additionally, Caenorhabditis elegans ability to withstand oxidative stress was improved by a methanolic extract of the edible red algae strain Chondrus crispus. The daf16 gene, the only ortholog of FOXO in the nematode C. elegans, showed a notable increase in transcription upon treatment with this methanolic extract. Furthermore, it has been discovered that kappa-carrageenan, a significant component of C. crispus water extract, causes C. elegans to activate daf16 in response to pathogen infection (Pseudomonas aeruginosa). 35 This is necessary for kappa-carrageenan to increase the immunological response. Furthermore, the lifetime of C. elegans was extended at 20, 30, and 35 °C by a fucose-containing polymer-rich fraction from the brown alga Ascophyllum nodosum. Under control settings (at 20 °C), fucose-containing polymer treatment had no effect on daf16 expression levels; however, under heat-stressed conditions (at 30 and 35 °C), it considerably increased. Studies on increased levels of FOXO3 and daf16 by seaweed derivatives have been conducted under stressful settings, including heat stress, pathogen infection, and oxidative stress, much as those on AMPK and/or SIRT activation. More research is necessary to determine whether these substances can block insulin/IGF-1 and activate FOXOs as people age. 35
Seaweed Bioactive Compounds Activate the NRF2 Pathway
Signaling Pathway of NRF2
In addition to the previously discussed important pathways, anti-inflammatory and antioxidant properties are also thought to help to prevent aging. Low-grade chronic inflammation and excessive oxidative stress may be factors in the beginning of aging. One of the main signs of aging is increased oxidative stress, which is linked to a number of age-related diseases. While the amount of different antioxidant enzymes falls with age, the generation of oxidants rises. 38
The overabundance of oxidative products damages cells. A transcription factor called NRF2 promotes cellular defenses against oxidative stress through its signaling pathway. The NRF2 repressor Kelch-like ECH-associated protein 1 (Keap1) is particular. NRF2 stays in the cytoplasm by interacting with Keap1. Keap1 conjugation also facilitates NRF2 degradation by functioning as an NRF2-specific E3 ligase adaptor protein. 58 Nrf2 separates from Keap1 and moves into the nucleus in a free and stable state when oxidative stress or other triggers are present. Nrf2 functions as a transcription factor inside the nucleus. The antioxidant response element (ARE), a DNA promoter found in the genes of many antioxidant proteins and detoxifying enzymes, is activated by NRF2 by dimerization with a small Maf protein. In addition to detoxifying enzymes like hemoxygenase1 (HO-1), glutathione S-transferase (GST), and NADPH: quinone oxidoreductase (NQO1), the Nrf2-ARE pathway can trigger antioxidant enzymes like superoxide dismutase (SOD) and catalase (CAT).38,58
Calorie restriction requires NRF2. NRF2 signaling guards against oxidative stress's after effects, including as aging and illnesses linked to aging. When exposed to acute stress, aging flies gradually lose their capacity to activate NRF2 targets. As people age, their ability to respond to stimuli like exercise by activating their NRF2-ARE downstream genes is compromised. Age-related functional loss can be countered by maintaining NRF2 signaling competence.38,58 It has been demonstrated that Keap1 loss-of-function mutations increase Drosophila lifespan, confirming NRF2 significance in regulating longevity. A comparison of mouse lifespans revealed a favorable correlation between lifespan and constitutive NRF2-signaling activity (ARE-binding activity). Reactivation of NRF2 can repair cellular HGPS abnormalities, while impaired activity of the NRF2 antioxidant pathway is a driving mechanism of Hutchinson-Gilford progeria syndrome (HGPS), a rare and always fatal premature aging disorder. Additionally, NRF2 exhibits tissue-specific anti-aging properties. For instance, compared to wild-type mice, the outer retina of Nrf2-deficient mice was more susceptible to age-related macular degeneration.38,58
Seaweed-Based NRF2 Activators
It has been observed that certain chemicals extracted from seaweeds displayed an NRF2 activation effect. IMR-32 neuroblastma and LNCaP prostate cancer cells’ NRF2-ARE pathway was strongly activated by extracts and a number of fractions made from cultivated green algae. The chosen fractions caused transcription of NQO-1, an NRF2 target gene, and nuclear translocation of NRF2. Furthermore, RAW 264.7 macrophages expressed the NRF2 protein in response to an ethanol extract of the marine brown algae S. serratifolium. In contrast to the ethanol extract's dose-dependent suppression of the Keap1 protein level, a downstream target, HO-1, boosted its protein expression. Furthermore, in the presence and absence of lipopolysaccharide, a phlorotannin-rich extract of another brown alga, E. cava, also stimulated the production of HO-1 and NRF2 in macrophages. Lipopolysaccharides encourage oxidative stress by activating macrophages’ membrane-bound NADPH oxidase.59,60
It has been demonstrated that a variety of algal substances, including polysaccharides, unsaturated fatty acids, sargaquinoic acid, and carotenoids, have the ability to activate NRF2. Isolated unsaturated fatty acid (C18:1(n-11)) from the green alga Ulva lactuca, which in human neuroblastoma IMR-32 cells activated cytoprotective genes controlled by the NRF2/ARE pathway, such as NQO1 and HO1. The ARE-activation activity of Ulva extract enriched with C18:1(n–11) was further confirmed by evaluation of several mouse organs, including the brain, heart, lung, liver, and stomach. 35
By encouraging NRF2-dependent cytoprotection, polysaccharides from the brown algae Sargassum fusiforme enhanced antioxidant defense and reduced stress insult in aging mice. As male mice aged, their livers’ levels of the Nrf-2 protein decreased in both the cytoplasm and the nucleus. Two months after middle-aged mice (9 months old) were given polysaccharide, the mice's livers showed enhanced nuclear accumulation of NRF2 and total protein expression. At the same time, the level of the NQO1 protein rose as well. 35
By activating the Nrf-2 signaling pathway, indole-6-carboxaldehyde (I6CA), which was extracted from Sargassum thunbergii, reversed oxidatively induced cell cycle arrest in Chinese Hamster lung fibroblasts. When cells were treated with both H2O2 and I6CA, the phosphorylation of Nrf-2 and the protein expressions of HO-1 and Nrf-2 were much higher than when cells were treated with H2O2 alone. Mycosporine-like amino acids (MAAs) are water-soluble metabolites that absorb ultraviolet light and are generated by seaweed. The NRF2-ARE pathway may be activated by two MAAs, shinorine and porphyra-334, which are competitive inhibitors of Keap1-NRF2 binding. Furthermore, when LPS was stimulated in RAW 264.7 cells, sargaquinoic acid, which was extracted from the brown algae Myagropsis myagroides, had anti-inflammatory properties. Treatment with sargaquinoic acid raised the levels of the nucleus NRF2 and total HO-1 protein in response to LPS. 61
Using the NRF2/ARE pathway, dieckol, which was isolated from Ecklonia stolonifera, activated NRF2 and raised the expression of NRF2 target proteins in HepG2 cells, such as HO-1, NQO-1, and GST. It was also discovered that eckol, another phlorotannin isolated from E. stolonifera, stimulated the nuclear translocation of NRF2 and caused HepG2 cells to produce HO-1. The brown algae Dictyopteris undulata contains zonarol, a para-hydroquinone-type pro-electrophilic chemical activated the NRF2/ARE pathway and induced NRF2 target genes like HO-1 and NQO1 in HT22 cells, which are neurons in the hippocampus.35,61 Furthermore, in reaction to glutamate, which causes cell death, zonarol improved the survival of HT22 cells. The activated NRF2/ARE pathway is at least partially responsible for zonarol's neuroprotective effects. In mouse hepatocytes BNL CL.2, fucoxanthin also enhanced the transcription of HO-1 and NQO1 and the accumulation of nuclear NRF2. The NRF2 activation effects of several classes of seaweed derivatives provide compelling evidence for the antioxidant properties of algal substances. Along with other aging-regulating mechanisms, such an NRF2-stimulating action may also have anti-aging effects.35,61
Conclusions and Future Prospects
The necessity to develop safe, dependable natural products with health-promoting qualities has increased due to the growing demand for nutraceuticals. Despite the fact that many macroalgae species have been used for millennia as food and natural remedies, there is no many research examining their potential anti-aging benefits. The possible effects of seaweed extracts or chemicals on pathways linked to aging are compiled in this review. At the cellular and tissue levels, the regulation of these pathways points to a potential function for macroalgal bioactive chemicals in anti-aging, occasionally, they also show neuroprotective, anti-obesity, and anti-diabetic actions. Senescence has been linked to a number of illnesses, including type II diabetes, obesity, and cognitive decline. The potential for using seaweed-derived bioactive chemicals in anti-aging applications is further supported by their ability to protect against these disorders. To learn more about the roles of these substances, research on the lifespan and health span extension effects of seaweed derivatives on elderly animals is required. In the meanwhile, it is important to investigate how sex affects the anti-aging properties of these compounds generated from seaweed. Several substances, including fucoxanthin and meroterpenoid, have demonstrated the capacity to control several anti-aging pathways. More research is needed on these substances to determine whether they have anti-aging properties, whether they have the ability to protect against age-related illnesses like cardiovascular and neurological disorders, and prolong life expectancy, particularly in higher animal models.
Footnotes
Acknowledgements
We would like to acknowledge SAGE journals for providing this opportunity.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Grant Number
Not Applicable
Informed Consent/ Patient Consent
Not Applicable
Trial Registration Number/Date
Not Applicable