
Abstract
Introduction
Skin ageing, a natural process of becoming old, is the most conspicuous sign of appearance. Given the widespread interest in retarding this process, 1 as research into anti-aging cosmetics is burgeoning, along with a heightened interest in natural extract-based materials suitable for cosmetic applications. 2 Molecules preventing changes in skin appearance encompass a spectrum of properties, including antioxidants, anti-wrinkle agents, and anti-inflammatory compounds.3–5 Extracts possessing these characteristics serve as essential ingredients in cosmetic formulations that reduce skin age. 3
Anti-oxidants work through enzymatic systems that activate enzymes to remove free oxygen, whereas non-enzymatic systems remove free radicals. Particularly, the non-enzymatic system scavenges free radicals and provides extra electrons for anti-oxidant molecules to form electron pairs in the skin. 6 Two common methods are used to assess oxidative stress protection: the 2,2′-azino-bis-3-ethylbenzthiazoline-6-sulphonic acid (ABTS) analysis, which measures the reduction of blue/green ABTS by anti-oxidants, and the 1,1-diphenyl-2-picrylhydrazyl (DPPH) assay, which evaluates the reduction of purple DPPH to 1,1-diphenyl-2-picryl hydrazine. 7
Wrinkle formation is caused by the excessive production of reactive oxygen species (ROS) under the influence of ultraviolet (UV) radiation, an increase in matrix metalloproteinases (MMPs), and the subsequent degradation of collagen and elastin. MMPs have different roles depending on their type. Enzymes responsible for collagen degradation encompass MMP-1 and MMP-3, whereas those targeting elastin degradation include MMP-7 and MMP-12. 8
Skin inflammation is a reaction that can be caused by agents, such as lipopolysaccharides (LPS) and UV radiation. During the inflammatory reaction, macrophages secrete inflammatory mediators, proinflammatory cytokines, and inflammatory enzymes, such as nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1), interleukin-6 (IL-6), inducible NO synthases (iNOS), and cyclooxygenase-2 (COX-2). 9 In particular, IL-6 is secreted by NF-κB and other factors in macrophages to promote immune responses. It functions alongside TNF-α and IL-1 to induce acute inflammatory responses and contribute to the progression of chronic inflammatory diseases. 10 Moreover, among the NO synthases, iNOS synthesizes large amounts of NO over extended periods, making it the most potent. 11 High concentrations of NO play a crucial role in suppressing inflammation, owing to their implications in the pathogenesis of cell death, immune response, and inflammatory diseases. 12
Spiraea salicifolia L. is a small ornamental shrub belonging to the Rosaceae family, characterized by its resistance to cold and tolerance to salinity.13,14 This plant is indigenous to Asian countries, including Korea, 15 China, 14 Russia, 16 as well as certain European countries. 17 It typically grows to 1–1.5 m, with its light red flowers blooming from June to July. 13 The young leaves of the plant are edible, and both the young leaves and roots are utilized for various medicinal purposes.14,15 Studies have reported that S. salicifolia possesses anti-oxidant, 16 anti-radical, anti-viral, 18 and α-glucosidase inhibitory activities. 19 Previous chemical studies on this species have reported the isolation of neolignan glycosides, 13 phenolic compounds, 15 and many types of flavonoids, such as vitexin, vicenin, luteolin, kaempferol, astragalin, trifolin, nicotiflorin, robinin, and hesperidin, as well as secondary metabolites. 20 However, S. salicifolia has not been well characterized for its effects and safety on the skin, and research on its chemical constituents remains limited.
Thus, in this study, we aimed to explore the anti-aging effects of S. salicifolia through assessments of its anti-oxidant, anti-wrinkle, anti-inflammatory properties, and local toxicity effects, particularly on the skin and eyes.
Materials and Methods
Plant Material and Sample Preparation
The stems of Spiraea salicifolia L. were collected from Gurye, Jeollanam-do Province, Republic of Korea, in November 2021. A voucher specimen (no. KKM-SS-21y) was maintained at the Skin & Natural Products Laboratory of Kolmar Korea, South Korea, for future reference. The dried and cut stem parts of S. salicifolia (1.69 kg) were extracted three times with methanol (20 L) at room temperature for 72 h to obtain S. salicifolia methanolic stem extracts (66.0 g). Samples for in vitro efficacy evaluation were used after dissolving the extracts in MeOH at 5% stock concentration.
ABTS Radical Scavenging Assay
A working solution (WS) of 7.4 mM ABTS was prepared by mixing 15 mM ABTS and 4.9 mM potassium persulfate in a 1:1 ratio. This mixture was reacted at room temperature, and light-shielded for 12–16 h. After adjusting the absorbance to 0.7 ± 0.02, the WS of 7.4 mM ABTS was used, and S. salicifolia stem extracts prepared at each concentration were transferred to a 96-well plate at an appropriate ratio, reacted at room temperature, and light-shielded for 7 min. Methanol was then added to the ABTS non-reaction port and allowed to react. Absorbance was measured at 734 nm using a microplate reader (Thermo, UV/Vis). The ABTS radical scavenging ability was calculated by the following formula:

DPPH Radical Scavenging Assay
Solutions of 0.25 mM DPPH and S. salicifolia stem extracts prepared at each concentration were mixed at a 1:1 ratio and transferred to a 96-well plate, followed by a 30 min reaction at room temperature in a light-shielded state. Then, ethanol was added to the DPPH non-reaction port and allowed to react. The absorbance was measured at 540 nm using a microplate reader (Thermo, UV/Vis). The DPPH radical scavenging ability was calculated using the following formula:

Cell Culture
CCD-986sk cells (ATCC, CRL-1947) were purchased from American Type Culture Collection (ATCC; Manassa, VA, USA) to verify wrinkle and elasticity efficacy. The cells were cultured in IMDM medium containing 10% fetal bovine serum (FBS) and 1% antibiotics-antimycotics (100X) under 5% CO2 at 37°C. RAW 264.7 cells (ATCC, TIB-71) were purchased from ATCC to verify their anti-inflammatory efficacy. The cells were cultured in DMEM/HIGH GLUCOSE medium containing 10% FBS and 1% antibiotics-antimycotics (100X) under 5% CO2 at 37°C.
Cell Viability Assay
The MTT assay was performed to evaluate the cytotoxicity of S. salicifolia stem extracts on CCD-986sk cells. The cells were seeded in a 96-well plate at a concentration of 5 × 104 cells/mL and incubated for 24 h at 37 °C under 5% CO2. After removing the culture medium, the cells were treated with various concentrations of S. salicifolia stem extracts and cultured for an additional 24 h. 20 μL of MTT solution (5 mg/mL) was added to each well, and the cells were incubated for 3 h before completing the culture. After removing the medium, 150 μL of DMSO was added, and the mixture was stirred at room temperature for 10 min. Absorbance was measured at 570 nm using a microplate reader (Thermo, UV/Vis).
The CCK-8 assay was performed to assess the cytotoxicity of S. salicifolia stem extracts on RAW 264.7 cells. The cells were seeded in a 96-well plate at a concentration of 5 × 105 cells/mL and incubated for 24 h at 37 °C under 5% CO2. After removing the culture medium, the cells were treated with various concentrations of S. salicifolia stem extracts and cultured for another 24 h. Then, 10 μL of CCK-8 solution was added to each well, and the cells were incubated for 2 h. After incubation, absorbance was measured at 450 nm using a microplate reader (Thermo, UV/Vis).
To confirm the LPS-induced cytotoxicity in RAW 264.7 cells treated with S. salicifolia stem extracts, the cells were treated with the extracts, and after 30 min, an appropriate amount of 1 μg/mL LPS was added and incubated for 24 h under the same conditions. Cell viability (%) was calculated using the following formula:
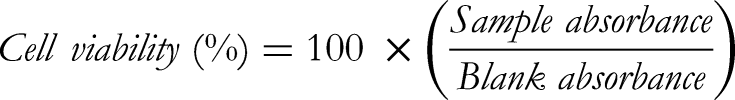
NO Assay
RAW 264.7 cells were seeded on a 96-well plate at a concentration of 5 × 105 cells/mL and incubated for 24 h at 37 °C under 5% CO2. After removing the medium, the cells were treated with various concentrations of S. salicifolia stem extracts and cultured for 30 min. Then, an appropriate amount of 1 μg/mL LPS was added and cultured for 24 h. After cultivation, the supernatant was transferred to a 96-well plate and reacted with a Griess receptor system kit (Promega) at room temperature in a light-shielded state. The absorbance was measured at 540 nm using a microplate reader (Thermo, UV/Vis). The standard curve was expressed by diluting the 0.1 M sodium nitrate standard. The amount of NO produced was calculated by the following formulas using the standard curve:

Real-Time Reverse Transcription PCR (RT-qPCR) (MMP-1, MMP-12 and IL-6)
To assess the effects of S. salicifolia stem extract on the mRNA expression of MMP-1 and MMP-12 in CCD-986sk cells, the cells were seeded in a six-well plate at a concentration of 3 × 105 cells/mL and incubated for 24 h at 37 °C under 5% CO2. For cell starvation, the medium was replaced with FBS-free medium, and the cells were incubated for 24 h. After removing the medium, the cells were treated with various concentrations of S. salicifolia stem extract and cultured for 24 h.
To evaluate the mRNA expression of IL-6 in RAW 264.7 cells treated with the S. salicifolia stem extract, the cells were seeded in a 6-well plate at a concentration of 5 × 105 cells/mL and incubated for 24 h at 37 °C under 5% CO2. After removing the medium, the cells were treated with various concentrations of the S. salicifolia stem extract. Subsequently, an appropriate amount of 1 μg/mL LPS was added, and the cells were cultured for an additional 24 h.
After the culture was complete, the cells were washed with HBSS, and RNA was isolated using TRIzol. An appropriate amount of nuclease-free water was added to the isolated RNA, which was then quantified. The quantified RNA was synthesized under conditions of 42 °C for 30 min, 70 °C for 10 min, and 4 °C for 1 min. RT-qPCR was performed by mixing cDNA, primers, SYBR green, and nuclease free water in an appropriate ratio and then reacting at 95 °C for 1 min using Realtime PCR (Applied Biosystems, Waltham, MA, USA; Quantstudio 5), followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. The following primers were used: Human GAPDH, forward 5′- GTC TCC TCT GAC TTC AAC AGC-3′ and reverse 5′-ACC ACC CTG TTG CTG TAG CCA A-3′; Human MMP-1, forward 5′-CCA GAT TTG CCA AGA GCA GA-3′ and reverse 5′-GAT GGG CTG GAC AGG ATT TT-3′; Human MMP-12, forward 5′-AGT TTT GAT GCT GTC ACT ACC G-3′ and reverse 5′-CAC TGG TCT TTG GTC TCT CAG AA-3′; Mouse GAPDH, Forward 5′- TTG GCA TTG TGG AAG GGC TC -3′ and Reverse 5′- ACC AGT GGA TGC AGG GAT GA -3′; Mouse IL-6, Forward 5′- CTG CAA GAG ACT TCC ATC CAG -3′ and Reverse 5′- AGT GGT ATA GAC AGG TCT GTT GG -3′. GAPDH was used as an internal control, and PCR result analysis yielded ΔΔCt values to compare the expression of the genes.
Western Blotting (iNOS)
RAW 264.7 cells were seeded on 60 mm dish at a concentration of 5 × 105 cells/mL and incubated for 24 h at 37 °C under 5% CO2. After removing the culture medium, the cells were treated with various concentrations of the S. salicifolia stem extract. Subsequently, an appropriate amount of 1 μg/mL LPS was added and cultured for 16 h. After culturing, the cells were washed with HBSS and separated. Then, 400 μL of the PRO-PREP protein extraction solution was added and incubated at −20 °C for 20 min. Proteins were obtained by centrifugation at 4 °C, 13,000 rpm, and 10 min. Bradford assay was performed, and the protein was quantified to be the same amount. The quantified proteins were electrophoresed on a 10% sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis gel and transferred onto a polyvinylidene fluoride membrane using a Trans-Blot Turbo Transfer System (Bio-Rad Laboratories, Hercules, CA, USA). The membrane was blocked at room temperature for 1 h with 5% bovine serum albumin and incubated for 20 h at 4 °C with a primary antibody. Subsequently, the membrane was washed with 1× Tris-buffered saline containing Tween (TBST; 0.3% Tween 20, w/v) and incubated with a secondary antibody at room temperature for 2 h. The protein bands were washed with 1× TBST, reacted with Western Chemical horseradish peroxidase Substrate (Merck Millipore) for a certain period, and visualized using a Chemical Documentation System (ATTO, Tokyo, Japan). Protein expression (%) was calculated using the following formula:

Skin Irritation Test Using RHE Model
KeraSkin (BioSolution, Seoul, Korea) was purchased from the manufacturer and used to evaluate skin irritation caused by the S. salicifolia stem extract. The test was conducted according to the OECD test guideline (TG) 439
21
and the manufacturer's instructions. Dulbecco's phosphate-buffered saline (DPBS) (Welgene, Gyeongsangbuk-do, Korea) and 5% SDS (Sigma-Aldrich) were used as negative and positive controls, respectively. The S. salicifolia stem extract was diluted to a final concentration of 1% in butylene glycol (Daicel Corporation, Osaka, Japan). Controls and test materials were exposed to 40 μL for 30 s at 37 °C in a 5% CO2 incubator. The tissues were incubated after rinsing with DPBS. MTT (Sigma) in Dulbecco's modified Eagle's medium (DMEM) (Welgene) was used to determine cell viability. Formazan was extracted with isopropanol (Sigma) and quantified using a spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Isopropanol was used as a blank for absorbance measurements. Tissue cell viability was calculated using the following equation:

Eye Irritation Test Using the RHCE Model
MCTT HCE™ (BioSolution) was purchased from the manufacturer and used for evaluating eye irritancy of the S. salicifolia stem extract. The test was conducted according to OECD TG 492 22 and the manufacturer's instructions. DPBS and methyl acetate (Sigma) were used as the negative and positive controls, respectively. The extract was diluted to a final concentration of 1% in butylene glycol (Daicel Corporation), as in the skin irritation test. Controls and test materials were exposed to 40 μL for 60 s at the same conditions of the skin irritation test. The tissues were incubated overnight after rinsing with DPBS. To determine cell viability, (3-4,5-dimethyl thiazole 2-yl) 2,5-diphenyltetrazolium bromide (WST-1) (Roche, Basel, Switzerland) in DPBS was used. Formazan was extracted by centrifugation at 200 g (Hanil Science, Kyonggi-do, Korea) and quantified using a spectrophotometer (Thermo Fisher Scientific). The 25X WST solution was used as a blank for absorbance measurements. The cell viability of the tissue was calculated, and the quality control of the RHCE model was examined in the same manner as the skin irritation test.
Extraction and Compound Isolation
The stem parts of S. salicifolia were dried and cut into pieces. The dried stems (1.69 kg) were extracted three times with 20 L of methanol (MeOH) for 72 h at room temperature to yield an extract (66.0 g). The MeOH extract was dissolved in double-distilled water and sequentially partitioned with n-hexane (n-Hex), methylene chloride (MC), ethyl acetate (EA), and n-butanol (n-BuOH) (each 750 mL×3) to produce layers of 5.18, 1.99, 3.75, and 6.30 g, respectively. The EA-soluble layer (3.22 g) was subjected to silica gel column chromatography (70-230 mesh) and eluted using an MC-MeOH stepwise gradient (50:1 → 20:1 → 10:1 → 5:1 → 3:1 → 2:1 → 1:1 → 1:5 → 1:10, v/v, each 1.2 L) to obtain nine fractions (Fr. E1–Fr. E9). Based on the HPLC results, further separation and purification of Fr. E5 and Fr. E7, the main fractions of the EA-soluble layer, were conducted. Fr. E7 (454.9 mg) was separated by preparative reverse-phase LC using a YMC-Actus Triart C18 column (250 × 20 mm i.d., 5 μm, 12 nm); flow rate, 11 mL/min; gradient conditions 20 → 48% acetonitrile (CH3CN) in water for 50 min; λmax 254 nm to obtain
HPLC Analysis
HPLC analysis was performed on the methanolic extract of the stems of S. salicifolia and compounds
Statistical Analysis
Each set of experiments was independently repeated three times, and the results are expressed as means ± standard deviations. Statistical analyses were performed using GraphPad Prism software (version 6.0; La Jolla, CA, USA), and differences between multiple groups were compared using one-way ANOVA and Tukey's test. A p-value of < 0.05 was considered statistically significant when comparing with the control group.
Results
Effect of Spiraea salicifolia Stem Extracts on Free Radical Scavenging Capacity
The ABTS and DPPH free radical scavenging capacities were measured to determine the anti-oxidant activity of S. salicifolia stem extracts. S. salicifolia stem extract showed increased anti-oxidant efficacy in a concentration-dependent manner. The ABTS and DPPH free radical scavenging abilities were 92% and 19%, respectively, at a concentration of 100 μg/mL (Figure 1). Vitamin C (Vit-C) eliminated 99% and 97% of ABTS and DPPH free radicals, respectively, at a concentration of 100 μg/mL.

Effect of S. salicifolia stem extracts on free radical scavenging capacity. (A) ABTS radical scavenging capacity, (B) DPPH radical scavenging capacity. Data are presented as means ± standard deviations (n = 3). p-values between the control and the S. salicifolia stem extracts groups are indicated (*** p < 0.001). Vitamin C (Vit-C) was used as the positive control.
Effect of Spiraea salicifolia Stem Extracts on Cell Viability in CCD-986sk Cells
We performed a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay to confirm the viability of CCD-986sk cells treated with S. salicifolia stem extracts. As a result of MTT assay, the S. salicifolia stem extracts showed no cytotoxicity to the cells at a concentration ≤100 µg/mL (Figure 2). In addition, since precipitation of the extract occurs at concentrations above 250 µg/mL, we judged the concentrations ≤ 100 µg/mL are suitable for cell experiments (data not shown). Therefore, subsequent experiments were conducted at concentrations of 25, 50, 75, and 100 μg/mL.

Effect of S. salicifolia stem extracts on CCD-986Sk cell viability. CCD-986sk cells were treated with multiple concentrations of the S. salicifolia stem extracts. The S. salicifolia stem extracts showed no toxicity at concentrations of 25, 50, 75, and 100 μg/mL. Data are presented as means ± standard deviations (n = 3). The p-values between the control and the S. salicifolia stem extracts groups are indicated (*p < 0.05).
Effect of Spiraea salicifolia Stem Extracts on Inhibiting MMP-1 and MMP-12 in CCD-986sk Cells
The expression levels of MMP-1 and MMP-12 mRNA were assessed via real time polymerase chain reaction (PCR) at various concentrations (25, 50, 75, and 100 μg/mL) of S. salicifolia stem extracts (Figure 3). The results showed that the S. salicifolia stem extracts significantly reduced MMP-1 expression at concentrations of 75 and 100 μg/mL, along with a notable decrease in MMP-12 expression at a concentration of 100 μg/mL. Specifically, the S. salicifolia stem extracts inhibited MMP-1 mRNA expression by −4, 24, 31, and 43% at concentrations of 25, 50, 75, and 100 μg/mL, respectively, while suppressing MMP-12 mRNA expression by 12%, 16%, 27%, and 34% at the same concentrations. Meanwhile, treatment with 10 μM Epigallocatechin gallate (EGCG) inhibited MMP-1 and MMP-12 mRNA expression by 90% and 64%, respectively.

Effect of S. salicifolia stem extracts on inhibiting mRNA expression of matrix metalloproteinases (MMPs) in CCD-986sk cells. CCD-986sk cells were treated with various concentration (25, 50, 75, and 100 μg/mL) of S. salicifolia stem extracts for 24 h. After treatment completion, mRNA expression of (A) MMP-1 and (B) MMP-12 was analyzed through real time polymerase chain reaction (PCR). Data are presented as means ± standard deviations (n = 3). The significance level was denoted as follows: * p < 0.05, ** p < 0.01, and *** p < 0.001, indicating the difference between the control and S. salicifolia stem extracts groups. Epigallocatechin gallate (EGCG) was used as the positive control.
Effect of Spiraea salicifolia Stem Extracts on Cell Viability in RAW 264.7 Cells
We performed a CCK-8 assay to confirm the viability of S. salicifolia stem extracts in RAW 264.7 cells. Consequently, the S. salicifolia stem extracts showed no cytotoxicity to the cells at concentrations ≤100 µg/mL in both LPS-free and stimulated RAW 264.7 cells (Figure 4). Therefore, subsequent experiments were conducted at concentrations of 25, 50, 75, and 100 μg/mL.

Effect of S. salicifolia stem extracts on RAW 264.7 cell viability. RAW 264.7 cells were treated with multiple concentrations of the S. salicifolia stem extracts. The S. salicifolia stem extracts showed no toxicity at concentrations of 25, 50, 75, and 100 μg/mL in both (A) unstimulated and (B) stimulated RAW 264.7 cells with 1 μg/mL LPS. Data are presented as means ± standard deviations (n = 3). The significance level denoted as *** p < 0.001 indicated the difference between the blank and negative control (LPS treated control).
Inhibiting Effect of Spiraea salicifolia Stem Extracts on NO, IL-6 and iNOS in RAW 264.7 Cells Stimulated with LPS
The Griess agent system kit was employed to measure NO production of S. salicifolia stem extracts at concentrations of 25, 50, 75, and 100 μg/mL post-stimulation with 1 μg/mL LPS (Figure 5A). Our findings revealed a significant reduction in NO production with S. salicifolia stem extracts across the tested concentrations. Relative to the negative control, NO production inhibition rates were 13%, 21%, 26%, and 46% at concentrations of 25, 50, 75, and 100 μg/mL, respectively. Moreover, treatment with 10 μM curcumin and 10 μg/mL celecoxib resulted in NO production inhibition rates of 74% and 60%, respectively, compared to the negative control.

Inhibitory effects of S. salicifolia stem extracts on RAW 264.7 cells exposed to lipoopysaccharides (LPS). RAW 264.7 cells were treated with 1 μg/mL LPS and 25, 50, 75, and 100 μg/mL of S. salicifolia stem extracts. (A) Nitric oxide (NO) production was analyzed by the Griess agent system kit after 24 h of sample treatment. (B) mRNA expression level of InterLeukin-6 (IL-6) was analyzed through real time polymerase chain reaction (PCR) after 24 h of treatment. (C) Protein expression level of nitric oxide synthase (iNOS) was analyzed by western blotting after 16 h of sample treatment. Data are expressed as means ± standard deviations (n = 3). * p-value (** p < 0.01 and *** p < 0.001) versus the negative control (LPS treated group). Positive controls: Curcumin and Celecoxib.
The mRNA expression levels of IL-6 were evaluated using real time PCR after stimulation with 1 μg/mL LPS and treatment with various concentrations of the S. salicifolia stem extract (25, 50, 75, and 100 μg/mL; Figure 5B). The results demonstrated a concentration-dependent decrease in IL-6 expression in response to treatment with the S. salicifolia stem extracts. Compared to the negative control, the inhibition rates for IL-6 expression were 18, 39, 39, and 42% at concentrations of 25, 50, 75, and 100 μg/mL of S. salicifolia stem extracts, respectively. Treatment with 10 μM curcumin and 10 μg/mL celecoxib as positive controls decreased IL-6 mRNA expression by 52% and 81%, respectively. However, no detectable changes in the mRNA expression of proinflammatory cytokines such as TNF-α and IL-1β were observed compared to the negative control (data not shown).
In addition, iNOS expression was assessed via western blotting after stimulation with 1 μg/mL LPS and treatment with concentrations of 25, 50, 75, and 100 μg/mL of S. salicifolia stem extracts (Figure 5C). Our results demonstrated a concentration-dependent reduction in iNOS expression with S. salicifolia stem extracts. Compared to the negative control, inhibition rates for iNOS expression were 22%, 56%, 62%, and 72% at concentrations of 25, 50, 75, and 100 μg/mL, respectively. Notably, treatment with 10 μM curcumin and 10 μg/mL celecoxib as positive controls led to 93% and 97% reduction in iNOS expression, respectively, compared to the negative control. Additionally, no significant change was observed in COX-2 protein expression relative to the negative control (data not shown).
Local Toxicity Test of Spiraea salicifolia Stem Extract
Reconstructed human-like models are often used for cosmetic safety assessments in accordance with alternative animal testing approaches. 23 Reconstructed human epidermis (RHE) is composed of a multilayered structure that mimics human skin properties and proliferates in primary normal human keratinocytes. Similarly, reconstructed human cornea-like epithelium (RHCE) is differentiated from human cornea. To evaluate the local toxicity of S. salicifolia stem extract, in vitro three-dimensional RHE and RHCE models were used according to the OECD test guidelines.21,22
The skin irritation test was implemented using the KeraSkin™ (Seoul, Korea) RHE model. The tested chemical was predicted to be non-irritative and showed a mean relative cell viability greater than 50%. Based on the prediction criteria, S. salicifolia stem extracts were defined as non-irritant to the skin (Table 1). An eye irritation test was performed using the MCTT HCE and RHCE models. The liquid tested chemical was predicted to be non-irritative, with a mean relative cell viability of approximately 35%. According to the prediction model, S. salicifolia stem extracts were defined as non-irritant (Table 2). None of the test materials showed color development, MTT, or WST reactivity in the pretest. This study met all acceptance criteria, including the absorbance value of the controls and the standard deviation between replicates.
Relative Cell Viability of the RHE Model in the Skin Irritation Test.

All data are presented as means ± standard deviations (n = 3).
NC, negative control; PC, positive control; SC, solvent control.
Relative Cell Viability of the RHCE Model in the eye Irritation Test.

All data are presented as means ± standard deviations (n = 2).
NC, negative control; PC, positive control; SC, solvent control.
The extract was classified under the UN Globally Harmonized System of Classification and Labelling of Chemicals (GHS) no-category in skin and eye irritation tests. Generally, extracts used as ingredients in cosmetics are combined with solvents or preservatives. 24 Butylene glycol was used as the extraction solvent. Considering the cell viability of the solvent control, S. salicifolia stem extracts did not cause skin or eye irritation.
Characterization of Purified Compounds
Chromatography was used to determine the main constituents present in the stems of S. salicifolia. Five known compounds, a terpene glycoside, three isoflavonoids, and a phenolic glucoside, were isolated from the stems of S. salicifolia. Based on nuclear magnetic resonance (NMR) (1H and 13C) and mass spectrometry (MS) data, and comparison with previously published paper,

Chemical structures of compounds
Spectroscopic Data for Compounds 1–5
Luteolin 5-O-β-D-glucopyranoside (
Yellow powder, C21H20O11, ESI-MS m/z = 447.0969 [M-H]−, 1H-NMR (400 MHz, DMSO-d6) and 13C-NMR (100 MHz, DMSO-d6): see Supplemental Material. Trans-resveratrol 3-O-β-D-glucopyranoside (
Yellow powder, C20H22O8, ESI-MS m/z = 389.1282 [M-H]−, 1H-NMR (700 MHz, DMSO-d6) and 13C-NMR (175 MHz, DMSO-d6): see Supplemental Material.
Hyperoside (
Yellow powder, C21H20O12, ESI-MS m/z = 463.0927 [M-H]−, 1H-NMR (400 MHz, DMSO-d6) and 13C-NMR (100 MHz, DMSO-d6): see Supplemental Material.
Apigenin 5-O-β-D-glucopyranoside (
Yellow powder, C21H20O10, ESI-MS m/z = 431.1016 [M-H]−, 1H-NMR (400 MHz, DMSO-d6) and 13C-NMR (100 MHz, DMSO-d6): see Supplemental Material.
Prunioside A (
White oily gum, C25H30O11, ESI-MS m/z = 505.1750 [M-H]−, 1H-NMR (400 MHz, CD3OD) and 13C-NMR (100 MHz, CD3OD): see Supplemental Material.
High-Performance Liquid Chromatography (HPLC) Results
As shown in Figure 7, the methanolic extract of the stems of S. salicifolia showed major peaks at retention times of 16.202, 17.242, 17.684, 18.214, and 24.364 min, corresponding to luteolin 5-O-β-D-glucopyranoside (
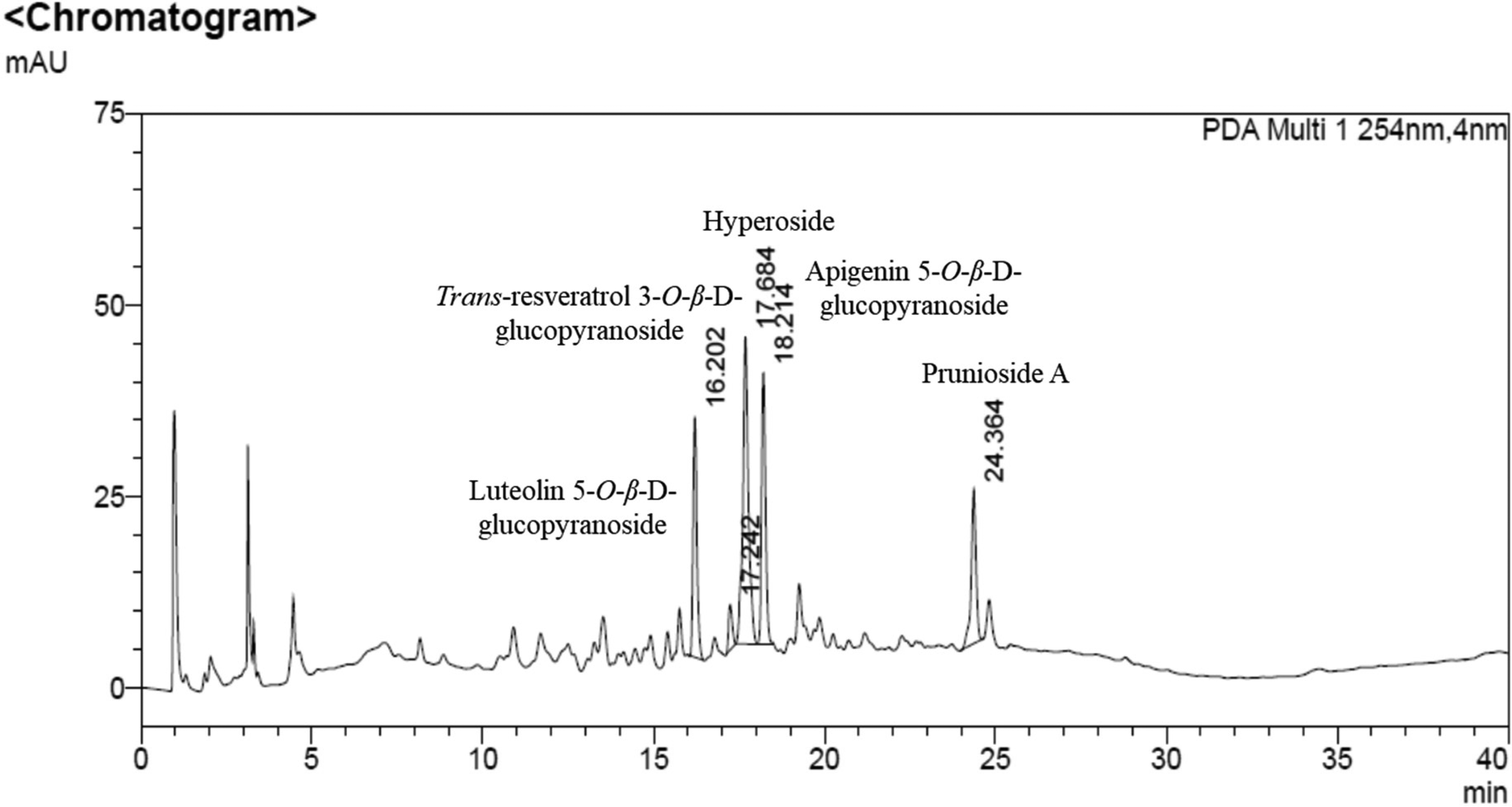
High-performance liquid chromatography (HPLC) chromatogram of S. salicifolia methanolic stem extracts at 254 nm.
Discussion
The purpose of this study was to elucidate the anti-aging effects of S. salicifolia stem extract and investigate its main chemical constituents. To achieve this, the extract was dissolved in methanol and evaluated for its antioxidant, anti-wrinkle, and anti-inflammatory properties through chemical and in vitro tests.
To assess the antioxidant effects of S. salicifolia stem extract, we evaluated its ABTS and DPPH radical scavenging activities. ABTS and DPPH contain a proton free radical with a characteristic absorption that is significantly reduced upon exposure to proton radical scavengers. Some dihydrochalcones and flavanones have been reported to exhibit higher reactivity with ABTS radicals compared to DPPH. These compounds demonstrate a strong ability to scavenge ABTS radicals, particularly against the pyrogallol structure, as observed in ABTS analysis for phenolic acids. 30 Vit-C, a potent reducing agent known as ascorbic acid, was used as the positive control. 31 As a result of the test, S. salicifolia stem extracts possess anti-oxidant properties akin to dihydrocalcones and flavanones, given their superior ABTS scavenging ability compared to that of DPPH.
To determine whether S. salicifolia stem extract improves wrinkles, we measured the inhibitory activities of MMP-1 and MMP-12 after performing an MTT assay using skin fibroblasts. MTT is a yellow tetrazolium salt that is converted into an insoluble purple formazan product by mitochondrial dehydrogenase of a metabolically active cell and, then, dissolved into dimethylsulfoxide. 32 MMPs can be divided into collagenases (MMP-1, MMP-8, and MMP-13), gelatinases (MMP-2 and MMP-9), stromelysins (MMP-3, MMP-10, and MMP-11), matrilysins (MMP-7 and MMP-26), membrane-type MMPs (MMP-14, MMP-15, and MMP-16), and MMPs that affect skin wrinkle and elasticity, which are divided by role as follows: MMP-1 destroys collagen types 1 and 3; MMP-3 and MMP-9 destroy collagen type 4; MMP-7 and MMP-12 destroy elastin. 33 Therefore, we measured MMP-1, a representative collagenase, and MMP-12, a representative elastase, to confirm the anti-wrinkle and elasticity effects of the S. salicifolia stem extracts. EGCG was used as a positive control, known for its anti-aging effects attributed to its inhibition of MMPs. 34 Thus, S. salicifolia stem extracts may contribute to anti-wrinkle and skin anti-aging effects through the inhibition of MMP-1 and MMP-12.
Furthermore, we evaluated the inhibitory activities of NO, IL-6, and iNOS based on the results of the CCK-8 assay in macrophages. CCK-8 is involved in the reduction of water-soluble orange-yellow formazan by dehydrogenase present in the cells. 35 NO is a representative inflammatory mediator in immune cells that plays a major role in inflammation. LPS promotes an inflammatory response by increasing NO production with increased iNOS expression in macrophages.12,36 To confirm the anti-inflammatory effects of S. salicifolia stem extracts, we assessed NO production and iNOS protein expression in RAW 264.7 cells stimulated with LPS. In addition, we investigated proinflammatory cytokines that are secreted from macrophages through the NF-κB and MAPK signaling and promote inflammatory responses by measuring IL-6 mRNA expression levels induced by LPS in Raw 264.7 cells. 37 We also utilized as positive controls curcumin, which regulates cytokines by inhibiting Toll-like receptor 4 (TLR-4) signaling, 38 and celecoxib, which exhibits anti-inflammatory effects as a COX-2-specific inhibitor. 33 As a result of the test, our findings suggest that S. salicifolia stem extracts exert anti-inflammatory effects by inhibiting NO production through iNOS downregulation and suppressing IL-6, a proinflammatory cytokine.
Therefore, our findings revealed that S. salicifolia stem extract exerts anti-aging effects by reducing oxidative stress, improving wrinkles, and alleviating inflammation. In addition, S. salicifolia stem extract has been identified as non-irritating and free of local toxicity in skin and eye irritation tests. Thus, it seems to be a topically safe ingredient suitable for use in cosmetics. The chemical investigation of S. salicifolia stem extract has led to five known compounds. Their structures were determined through NMR and MS to be luteolin 5-O-β-D-glucopyranoside (
Luteolin 5-O-β-D-glucopyranoside (
Apigenin 5-O-β-D-glucopyranoside (
The anti-aging effects of S. salicifolia stem extracts may be attributed to these five compounds identified as major components through HPLC analysis. These compounds should be further studied for their anti-aging activity. Furthermore, additional research on the cosmetic applications of the extract and clinical trials of creams formulated with it should be pursued to further validate its efficacy and potential.
Conclusions
In this study, the anti-aging capacity of S. salicifolia stem extracts was evaluated via their antioxidant, anti-wrinkle, and anti-inflammatory effects, and their chemical constituents were identified. The extracts exhibited antioxidant effects against ABTS and DPPH radicals. Moreover, they demonstrated anti-wrinkle effects by inhibiting collagenase MMP-1 and elastase MMP-12. Additionally, the extracts exerted anti-inflammatory effects by inhibiting NO, IL-6 and iNOS. They were also defined as non-irritants in skin and eye irritation tests.
Five known compounds—luteolin 5-O-β-D-glucopyranoside (
HPLC analysis confirmed that these molecules were the main compounds present in S. salicifolia stem extracts. Compounds
In conclusion, S. salicifolia stems may be valuable in cosmetic applications as safe, natural ingredients. It could be applied in various skincare formulations, such as serums, lotions, and creams, particularly in anti-aging cosmetics. Further studies on S. salicifolia could reveal its potential as an ingredient in cosmetics, food, and pharmaceuticals.
Supplemental Material
sj-docx-1-npx-10.1177_1934578X251322711 - Supplemental material for Anti-aging Effects and Main Chemical Constituents of
Spiraea salicifolia
Stem Extracts
Supplemental material, sj-docx-1-npx-10.1177_1934578X251322711 for Anti-aging Effects and Main Chemical Constituents of
Footnotes
Acknowledgments
The authors thank the editor and the reviewers for their useful feedback that improved this paper.
Authors’ Contributions
Gwee Kyo Park designed the research; Gwee Kyo Park, Jin Yeong Song, Hye-Young Yoo, and Minseo Kwon wrote the manuscript; Gwee Kyo Park, Jin Yeong Song, and Minseo Kwon performed the research; Gwee Kyo Park, Jin Yeong Song, Hye-Young Yoo, Minseo Kwon, and Ki Yong Lee analyzed the data; You Ah Kim and Sung-Ha Park supervised the research; Byoung Jun Park revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Availability of Data and Materials
All data and databases used and/or analysed during the current study are available from the corresponding author on reasonable request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical Approval is not applicable for this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Note
Statement of Human and Animal Rights: This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no human subjects in this article, and informed consent is not applicable.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.