
Abstract
Introduction
The genus Acronychia has been discovered to consist of over 50 species, primarily shrubs or trees taller than 30 meters, belonging to the Rutaceae family. They are widely distributed in India, Sri Lanka, China, Vietnam, Malesia, Australia, and islands in the Western Pacific Ocean.1–4 Similar to other members of the Rutaceae family, their leaves release a fragrant scent when crushed.4,5 Most species of the Acronychia genus have long been used in traditional folk medicine of indigenous Asian and Australian populations to treat bacterial and fungal infections, as well as for antispasmodic, gastrointestinal, fever-reducing, and hemostatic purposes. Additionally, the fruits and aerial parts are used as food in salads and spices, while essential oils (EOs) extracted from the flowers and leaves are utilized in cosmetics production. 6 Various groups of compounds have been discovered in species of this genus, including phenolics, flavonoids, terpenoids, alkaloids, etc Particularly, this genus contains characteristic acetophloroglucinol compounds with significant pharmacological values. Evidence from previous studies indicates that plant extracts from some Acronychia species and their secondary metabolites have the potential to induce cell toxicity, exhibit antioxidant, antibacterial, and anti-inflammatory properties.3,7–12
Three species belonging to the Acronychia genus have been discovered in Vietnam. Acronychia pedunculata (Vietnamese name: Bưởi bung, bí bái) is one of these species and is common in mountainous regions across the country. A. pedunculata is familiar to many local people and has been widely used in traditional medicine. Specifically, the stems, leaves, and fruits of this species are used to treat various ailments including boils, dermatitis, rashes, pain, arthritis, diarrhea, coughs, asthma, and bleeding disorders.4,13,14 Many volatile and non-volatile compounds have been identified in this species, such as polyphenols, terpenoids, and alkaloids,4,10 as well as volatile compounds from its EOs, including α-pinene, (E)-β-caryophyllene, caryophyllene oxide, and linalool.15–17 Studies on the chemical composition of the EOs in the central and northern regions of Vietnam have shown the influence of geographical factors and growing environments on the composition of the plant's EOs, with different parts of the plant containing different oil compositions. In addition to their chemical composition, the EOs from this plant species have been shown to have larvicidal activity, antimicrobial activity against various bacteria, particularly Salmonella enterica and Staphylococcus epidermidis, antifungal properties, and cytotoxic effects.15,16,18 In this study, we aimed to identify the chemical components present in the root, leaf, and flower EOs of this plant species collected from the south of Vietnam. Our goal was to determine whether there are any differences compared to specimens from the central and northern regions. Currently, the use of EOs for prevention and treatment of diseases has received attention. Evidence has shown the potential of EOs in supporting treatment of diabetes. For example, EOs from parsley, savory, and rosemary demonstrated prebiotic properties by promoting the growth or restoration of beneficial gut bacteria in humanized mice receiving fecal transplants from patients with ischemic heart disease and type 2 diabetes mellitus. 19 The nutraceutical and cosmeceutical industries have made efforts to incorporate EOs with tyrosinase inhibitory properties to deter tanning and promote skin lightening. Hence, there is a demand to investigate natural anti-tanning agents that both are safe for utilization in these sectors and have minimal adverse effects. In this study, we evaluated the inhibitory effects of EOs on α-amylase and tyrosinase. This suggests the potential use of A. pedunculata EOs in managing diabetes and addressing skin pigmentation disorders, thereby creating opportunities for their utilization in the nutraceutical and cosmeceutical sectors.
Results and Discussion
Chemical Composition
The chemical compositions of the EOs from the roots, leaves, and flowers of A. pedunculata were analyzed and identified by GC/MS, and their data are presented in Table 1. The chromatographic peaks representing the volatile organic compounds in the EOs are shown in Figures S1, S2, and S3 (Supplemental Material).
Chemical Compositions of the EOs from the Roots, Leaves and Flowers of Acronychia pedunculata.
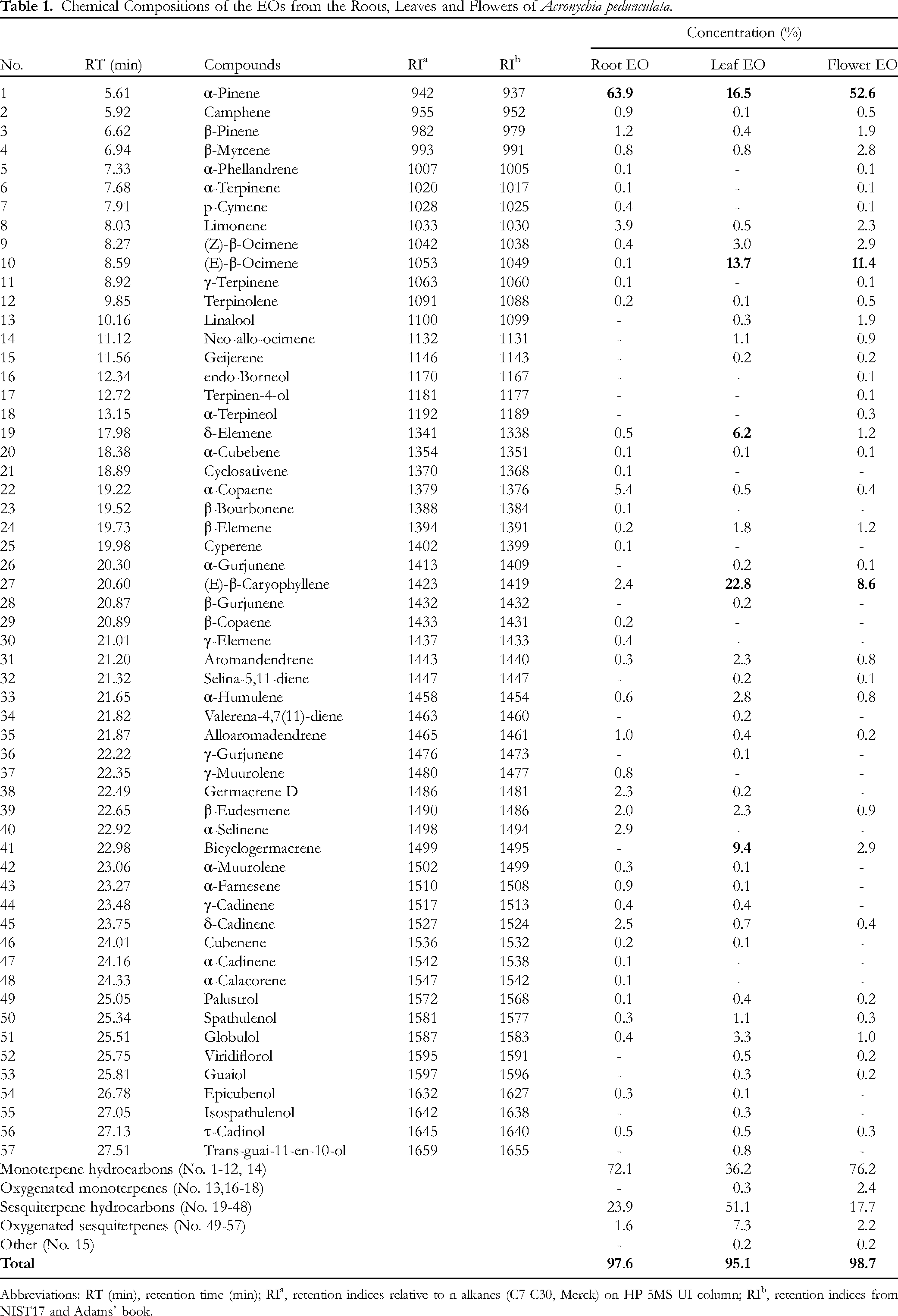
Abbreviations: RT (min), retention time (min); RIa, retention indices relative to n-alkanes (C7-C30, Merck) on HP-5MS UI column; RIb, retention indices from NIST17 and Adams’ book.
The results demonstrated the presence of 57 volatile compounds in the EOs, accounting for 97.6% (roots), 95.1% (leaves), and 98.7% (flowers) of the total oil content. Among them, the most abundant class of compounds was monoterpene hydrocarbons, making up 36.2–76.2% of the total oil content. This class comprised 13 compounds, with α-pinene being the most abundant compound, ranging from 16.5–63.9% of the total oil content. It was also the predominant compound in the composition of the examined EOs. Sesquiterpene hydrocarbons were the second most abundant class of compounds, comprising 17.7–51.1% of the total oil content. As seen in Table 1, this class is constituted of 30 compounds in total, with the highest number of compounds detected in the root EO (24 compounds), followed by the leaf EO (21 compounds). The most dominant compound in this class was (E)-β-caryophyllene, accounting for 22.8% and 8.6% of the total oil content of the leaf EO and flower EO, respectively. Bicyclogermacrene was another major sesquiterpene hydrocarbon, accounting for 9.4% and 2.9% of the EOs from the leaves and flowers. The results reported oxygenated monoterpenes and oxygenated sesquiterpenes were the minor chemical classes in the EOs. The former class is made up of four compounds, which were detected only in the leaf and flower EOs. The latter class consists of nine compounds, which were detected in all the studied EOs.
As seen in Figure 1, the distribution of the volatile compounds in the EOs from A. pedunculata is displayed using a Venn diagram. The root and leaf EOs were similarly found to contain more compounds (41) than their flower counterpart (37). The two EOs also possessed 28 constituents in common. The findings also revealed that the root EO shared 26 constituents with the flower EO, whereas the leaf and flower EOs shared 30 constituents. All the three EOs possessed 22 compounds in common, with pinenes being one of these shared constituents.

A Venn diagram for the numbers of volatile organic compounds present in the A. pedunculata EOs.
Previously, EO composition of leaves, branches, and fruits of this plant species collected in Duc Trong district, Lam Dong province, the Central Highlands region in Vietnam was shown that the content of oxygenated sesquiterpenes predominated, forming up to 85.1% of the total oil contents. The most abundant compounds were caryophyllene oxide (35.3%) and spathulenol (31.9%). 17 In the current study, caryophyllene oxide was not detected while spathulenol was found at very low levels (0.3-1.1%). In another study, an EO from aerial parts (ie, branches, leaves, and fruits) of A. pedunculata harvested in Me Linh district, Vinh Phuc province, Northern Vietnam, was shown to contain α-pinene (57.4%) and (E)-β-caryophyllene (13.6%) as the main components. 16 The concentration of α-pinene was higher while the concentration of (E)-β-caryophyllene was lower compared to the leaf EO analyzed in the current study. Altogether, these investigations into the EOs from A. pedunculata collected in different regions aforementioned provide evidence of geographical and climatic influences on the variations in the composition of this plant's EOs.
Inhibition of α-Amylase
Table 2 presents the α-amylase inhibitory activity of EOs extracted from different parts (flowers, leaves, and roots) of A. pedunculata, along with the reference standard acarbose. The inhibitory activity is reported as the percentage of enzyme inhibition at various concentrations of the EOs and acarbose, ranging from 31.25 to 2000 μg/mL. Additionally, the table provides the IC50 values (μg/mL), which represent the concentration required for 50% inhibition of α-amylase activity. For the flower EO, the inhibition percentage against α-amylase exhibited a gradual increase as the concentration of the EO was raised within the range of 0 to 1000 μg/mL, suggesting a linear relationship within this concentration range. However, the inhibition appeared to plateau at concentrations than 1000 μg/mL, indicating potential saturation or limitations in the inhibitory effect. Regarding the root and leaf EOs, the percentage inhibition increased steadily with increasing concentrations up to 125 and 500 μg/mL, respectively. All the EOs displayed a dose-dependent inhibitory effect on α-amylase, with higher concentrations leading to increased enzyme inhibition. The α-amylase inhibitory activity varied significantly among the EOs extracted from different plant parts. The root EO exhibited the strongest inhibitory activity, with an IC50 value of 53.73 ± 1.94 μg/mL, followed by the leaf EO (IC50 = 283.01 ± 36.59 μg/mL) and the flower EO (IC50 = 578.47 ± 21.62 μg/mL). As seen in Table 2, from 125 to 500 μg/mL, the inhibition percentage of the root EO was about 1.6–2 and 2–7.7 times as high as those of the leaf and flower counterparts, respectively. Compared to the reference standard acarbose (IC50 = 88.80 ± 1.87 μg/mL), the root EO exerted a significantly stronger inhibitory activity against α-amylase. It is also noted that the anti-α-amylase of the A. pedunculata EOs may generally be stronger than those from species in the Zingiberaceae family, such as Zingiber officinale (IC50 = 684 μg/mL), Zingiber atroporphyreus (IC50 = 300 μg/mL), and Wurfbainia schmidtii (1850 μg/mL).20,21
α-Amylase Inhibitory Activity of the Acronychia pedunculata EOs and Acarbose.

*: The percentage inhibition was calculated at the different EO concentrations (31.25-2000μg/mL).
Data are presented as mean ± standard deviation (n = 3).
n.a.: not available
Different letters (a, b, c, d) denote significant differences in the α-amylase activity among the samples (p < 0.05).
There is evidence to suggest that several Acronychia species, such as A. pedunculata, A. oligophylebia, A. laurifolia, and A. baueri, have a potential to inhibit α-amylase due to skimmianine in their composition. This compound was shown to strongly inhibit carbohydrate-digestion enzymes. 22 Other than this, limited information about anti-α-amylase activity of EOs and extracts from Acronychia species has been documented. Previous research into α-amylase inhibitory activity of EOs from species belonging to the Rutaceae family revealed that they may exert powerful influences on carbohydrate hydrolyzing enzymes. For example, EO from Toddalia asiatica aerial parts showed a modest inhibition of α-amylase. 23 Clausena excavata leaf EO displayed an anti-α-amylase activity which was comparable to that of acarbose. 24 EO from Citrus aurantium, a commonly found tree species in Southern and Eastern Asia, were shown to be potent inhibitors of α-amylase. 25
In our investigation, the predominant compounds identified in the A. pedunculata EOs were α-pinene, β-(E)-ocimene, (E)-β-caryophyllene, α-copaene, and bicyclogermacrene. Prior studies have revealed that inhibition of α-amylase demonstrated by EOs extracted from plants in the Rutaceae family may be linked to the presence of monoterpenes and sesquiterpenes in their composition. For instance, it is suggested that caryophyllene could play an important role in α-amylase inhibitory activity of Paramignya scandens EOs. 8 Pinenes as major constituents in EOs of Citrus plants were shown to be effective α-amylase inhibitors.26,27 In addition to these compounds, α-copaene was reported to contribute to inhibitory effect on α-amylase. 28 Furthermore, the anti-α-amylase activity observed in these plant species could result from synergistic or additive effects arising from the diverse constituents present in the complex mixture of EOs, rather than being solely attributed to individual compounds.
Inhibition of Tyrosinase
As shown in Table 3, within the tested concentration range (125-1000 μg/mL), there was a consistent increase in the percentage of enzyme inhibition as the concentrations increased. This suggests that the EOs’ inhibitory activity against tyrosinase followed a concentration-dependent pattern. The root EO presented a more pronounced concentration-dependent response, reaching 59.10% inhibition at 1000 μg/mL. Additionally, the root EO, with an IC50 value of 815.53 ± 3.24 μg/mL, exhibited the highest tyrosinase inhibitory activity among the EOs. The flower and leaf EOs displayed relatively lower inhibitory activities (IC50 values > 1000 μg/mL), indicating weaker potency against tyrosinase. The results also showed kojic acid had a significantly higher inhibition of tyrosinase, with an IC50 value of 60.02 ± 1.25 μg/mL, compared to the EOs. It is also noted that the antityrosinase of the A. pedunculata EOs could be more potent than those of Cinnamomum cassia (IC50 = 6160 μg/mL). 29
Tyrosinase Inhibitory Activity of the Acronychia pedunculata EOs and Kojic Acid.
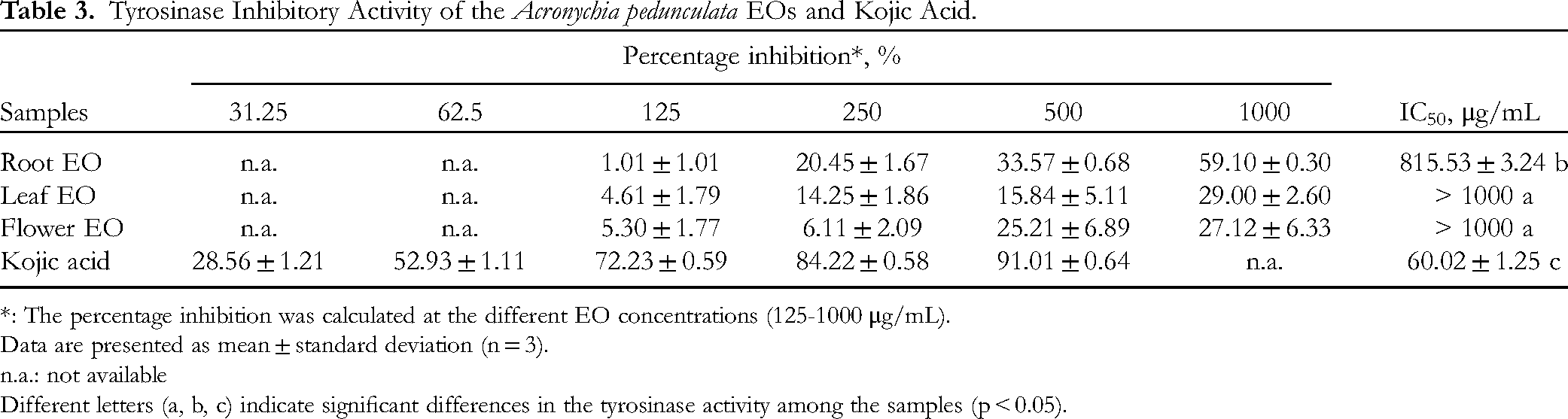
*: The percentage inhibition was calculated at the different EO concentrations (125-1000 μg/mL).
Data are presented as mean ± standard deviation (n = 3).
n.a.: not available
Different letters (a, b, c) indicate significant differences in the tyrosinase activity among the samples (p < 0.05).
Previously reported, certain volatile organic compounds, such as α-pinene, ocimene, and caryophyllene, identified in EOs of plants in the Rutaceae family, have demonstrated tyrosinase inhibitory effects.30–32 One recent study has shown that spathulenol identified in Paramignya scandens EOs may have a potential to inhibit tyrosinase. 8 Another recent study on EOs of Citrus plants has revealed that α-melanocyte-stimulating-hormone induced B16BL6 cells treated with limonene, β-elemene, and farnesene significantly suppressed the release of tyrosinase. 33 The current study has previously discussed the presence of these compounds in the EOs obtained from A. pedunculata. This suggests that these compounds may contribute to the antityrosinase activity exhibited by the EOs.
This study marks the first documentation of the anti-α-amylase and antityrosinase activities observed in the flower, leaf and root EOs of A. pedunculata. The findings of this research affirm the potential of A. pedunculata as a source of both antidiabetic and skincare products. However, further experiments are needed to substantiate the inhibitory effects of A. pedunculata EOs on α-amylase and tyrosinase enzymes.
Molecular Docking
Evaluation of the Docking Model
Before screening compounds, the co-crystal ligand was re-docked to the active site of the target to determine the root mean square deviation (RMSD), which helps to evaluate the suitability of the docking parameters. To evaluate the similarity structural, we determined the RMSD value by Chimera software. As a result of evaluating the docking model, our RMSD was obtained as 0.721 Å for QV4 and 0.417 Å for KOJ. The values all agree with RMSD < 1.5 Å (Figure S4, Supplemental Material).
Molecular Docking of Compounds to the Target Protein
After preparing ligand, we docked five compounds to screen for enzyme 2QV4 and 3QN1 inhibitory activity. Our results obtained two compounds, which have negative docking score (ΔG) smaller than the positive control kojic acid (ΔG = −5.6 kcal/mol) were shown in Table 2.
As seen in Table 4, two compounds limonene and bicyclogermacrene have lower binding energy values with 3QN1 than the positive control kojic acid. Therefore, these two compounds need to be further evaluated for their potential to become drugs in the future. Figure S5 is the 2D interaction results of limonene, bicyclogermacrene and the positive control compound with protein 3QN1. Accordingly, limonene shows the strongest interaction with the tyrosinase enzyme through bonds with amino acids: HIS 204, HIS208, HIS42, HIS 60. These are amino acids located in the active site of the protein 3QN1. 34 At the same time, this compound also shows the ability to form bonds with amino acids similar to positive controls: HIS 204, HIS60, VAL218. On the other hand, bicyclogermacrene is predicted to have weak inhibitory ability as it shows little interaction with the target protein at the active site.
The Docking Results of Five Compounds.

Conclusions
The current investigation revealed that EOs derived from the roots and flowers of A. pedunculata, collected in Tay Ninh province, Vietnam, are primarily composed of monoterpene hydrocarbons, while the EO extracted from the plant's leaves is predominantly composed of sesquiterpene hydrocarbons. Major compounds identified in the EOs include α-pinene, β-(E)-ocimene, (E)-β-caryophyllene, and bicyclogermacrene. This study reports, for the first time, the in vitro anti-α-amylase and antityrosinase activities of these EOs. Notably, the root EO exhibited stronger inhibitory effects on α-amylase and tyrosinase compared to the leaf and flower EOs. Additionally, a molecular docking study was conducted on the five compounds present in the EO against α-amylase and tyrosinase. The findings attribute the tyrosinase inhibitory activity to the presence of limonene, suggesting its potential as a lead compound for further antityrosinase investigations. These results contribute additional scientific evidence regarding the biological activity of A. pedunculata EOs. Further research is warranted to deepen our understanding of their mechanisms of action and explore additional bioactive properties.
Experimental
Plant Collection and Identification
The roots, leaves, and flowers of A. pedunculata were gathered from Tan Chau district, Tay Ninh province, Vietnam (11°35′52.9″ N; 106°23′19.5″ E) in August 2023. The plant material was identified by Assoc. Prof. Dr Nguyen Hoang Tuan (Faculty of Pharmacognosy and Traditional Medicine, Hanoi University of Pharmacy, Vietnam). A voucher specimen (code: AP-NP-HC35) has been securely stored at the Department of Chemistry, Vinh University, Vinh City, Nghean, Vietnam.
Chemicals
A series of C7–C30 n-alkanes, sodium sulfate (Na2SO4), and dichloromethane (CH2Cl2) were purchased from Merck (Darmstadt, Germany). Acarbose (94.1%) was obtained from the National Institute of Drug and Quality Control in Hanoi, Vietnam. Kojic acid, p-Nitrophenyl-α-D-glucopyranoside, tyrosinase from mushroom, and α-amylase from Bacillus sp. were obtained from Sigma-Aldrich (St. Louis, MO, USA).
GC/MS Analysis
In our current study, the EOs from the roots, leaves, and flowers of A. pedunculata were analyzed by the GC/MS system including an Agilent 7890B GC System equipped with a 5977B MSD model. The chemical compositions were identified according to the standard methods outlined in our previous study.8,35
α-Amylase Inhibitory Activity
The enzyme inhibition assay was carried out according to a previously described methodology. 36 In brief, a diluted EO from A. pedunculata was added to an α-amylase solution (0.14 U/mL, 10 µL), followed by incubation for 15 min at 37 °C. To initiate the reaction, a 0.25% starch solution was added, followed by another 15-min incubation at 37 °C. A blank sample underwent the same steps, except without the addition of α-amylase. The enzymatic reaction was terminated by adding 50 µL of 1 M HCl. Subsequently, KI3 solution (100 µL) was introduced, and the absorbance was determined at 595 nm. The percentage of enzyme inhibition was calculated using the formula:
Percentage of inhibition (%) = [1 – (Absorbance of Sample/Absorbance of Blank)] × 100%. The IC50 values (μg/mL), which represent the concentration required for 50% enzyme inhibition, were determined from the inhibition curve constructed using Microsoft Excel. The EO was evaluated at concentrations of 31.25–2000 µg/mL to estimate its inhibitory activity. Acarbose, a reference standard, was tested at concentrations of 31.25–500 µg/mL.
Tyrosinase Inhibitory Activity
Tyrosinase inhibition capacity of the EOs was evaluated following a previously reported method. 37 Diluted EOs ranging from 125–1000 μg/mL (100 μL) were mixed with 40 μL of tyrosinase (80 U/mL) and 40 μL of 0.5 mM L-DOPA substrate. The resulting mixture underwent a 20-min incubation at 37 °C, after which the absorbance was measured at 490 nm. Kojic acid was used as a reference standard inhibitor. IC50 value (μg/mL) was determined to assess the inhibitory effect of the EOs against the tyrosinase enzyme.
Preparation of Protein Structures
The crystal structure of the enzyme α-amylase (ID: 2QV4) and tyrosinase (ID: 3QN1) was retrieved from the Protein Data Bank RCSB (https://www.rcsb.org/). The 2QV4 complex contains the co-crystal ligand QV4 and 3QN1 complex contains the co-crystal ligand KOJ. We removed water molecules and co-crystals ligands from the protein molecule by using Discovery Studio software. Next, we used MGL Autodock Tools 1.5.7 software to add hydrogen atoms, optimize polar hydrogens, and Kollman charges. Based on previous publications about the active site of proteins 2QV4 and 3QN1, we have selected the active site with the grid box size as: (70 Å×70 Å×70 Å), the grid center (x,y,z) corresponding (16,11; 49,57; 25,06) and (60 Å×60 Å×60 Å), the grid center (x,y,z) corresponding (−9,16; −19,73; 6,98). This protein was saved in pdbqt format to prepare for the docking program. 38
Preparation of Ligands
Based on the results of isolation of compounds in the roots, leaves and flowers of A. pedunculata and the feasibility of molecular docking, compounds were collected and structures downloaded from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) in SDF format. Then we converted them to PDB format using Chimera software. Finally, the ligands were optimized by Avogadro software using Conjugate Gradients method and converted to the PDBQT format using Autodock Tools 1.5.7.
Performance of Molecular Docking
Then we proceed molecular docking enzyme 2QV4 and 3QN1 with ligands by using Autodock Tools software. Positive controls acarbose and kojic acid was downloaded from PubChem database. The docking scores of positive controls is used for selecting compounds with α-amylase and tyrosinase inhibitory effects.
Evaluation of Docking Results
To evaluate the docking results, the co-crystal ligand after being separated from the protein was redocked to the active site of the target. The process was performed successfully if the root-mean-square deviation (RMSD) value was less than or equal to 1.5 Å. For substances that need docking, their binding ability is assessed through interaction with amino acids and the interaction energy is calculated by the scoring function of Autodock Vina.
Statistical Analysis
The experiments were carried out in triplicate, and the data were reported as mean ± standard deviation. To assess the significant differences in bioactivities between the EOs and reference standards, statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's post-hoc test at a significance level of p < 0.05. The statistical analyses were performed using Minitab 19 software (Pennsylvania, USA). Venn diagram was prepared using Microsoft Excel.
Supplemental Material
sj-docx-1-npx-10.1177_1934578X241285530 - Supplemental material for Acronychia pedunculata Essential Oils from Vietnam: Chemical Constituents, Enzymatic Inhibitory Activities and Molecular Docking Study
Supplemental material, sj-docx-1-npx-10.1177_1934578X241285530 for Acronychia pedunculata Essential Oils from Vietnam: Chemical Constituents, Enzymatic Inhibitory Activities and Molecular Docking Study by Nguyen T. Kim-Anh, Trang H.D. Nguyen, Danh C. Vu, Duc Giang Le, Hoang Van Trung, Bui Thanh Tung, Nguyen Thi Phuong, Le Thi Huong and Nguyen Thi Ngan in Natural Product Communications
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical approval is not applicable to the article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.