
Abstract
To investigate the scientific connotation of harvest period for Prunella vulgaris L. (P vulgaris, known as Prunellae Spica), the triterpenoids and phenols of Prunellae Spica in developmental stages were quantified by high performance liquid chromatography, and the anti-inflammatory and anti-breast cancer properties of which were investigated. Furthermore, Grey correlation and Pearson correlation analysis were used to screen the anti-inflammatory and anti-breast cancer–related effective ingredients, and a multidimensional network of “ingredient-target-pathway” through network pharmacology was constructed. The results showed that the harvest time of Prunellae Spica was closely related to its chemical composition and pharmacological activity. Phenols, such as salvianic acid A, caffeic acid, and salviaflaside, mainly accumulated in late development, while rosmarinic acid showed the opposite. Triterpenes, such as oleanolic acid and ursolic acid, mainly accumulated in early development, while betulinic acid accumulated during ripening. The anti-breast cancer activity of Prunellae Spica in early development was stronger than that in the later, but the anti-inflammatory activity in late development was stronger than that in the early stage. Significantly associated with anti-inflammatory activity in Prunellae Spica was salviaflaside, which may regulate TNF and NOD-like receptor signaling pathways by acting on targets such as CASP7, CASP8, CASP3, NOD2, and CASP1. Significantly associated with anti-breast cancer activity were oleanolic acid and ursolic acid, which may regulate Ovarian steroidogenesis and Prolactin signaling pathways on targets such as PTGS2, CYP19A1, ESR2, CYP17A1, and MAPK3. These results suggest that P vulgaris could be harvested before ripening for its anti-breast cancer use, and after ripening for its anti-inflammatory use.
Introduction
Prunella vulgaris is a perennial herb of the Lamiaceae family, which has been used in China for thousands of years. The dried spica of P vulgaris, which are collected in the summer, can be used as the standard medicinal material in the Chinese Pharmacopoeia, while it is more commonly used with whole grass in Europe and Taiwan. Prunella vulgaris is light and crisp, with a slight fragrance and light taste. Traditionally, it is preferred for P vulgaris to have a purple-brown color and large ears. 1 According to the theory of traditional Chinese medicine, P vulgaris is bitter, pungent, and cold in nature. It has the effects of clearing fire, improving eyesight, reducing swelling, and dispelling masses. It is often used to treat red and swollen eyes, night pain, headache, dizziness, scrofula, gallbladder tumors, breast carbuncles, breast addiction, and breast tenderness. 2 Modern pharmacological studies have shown that P vulgaris mainly contains phenolic acids, flavonoids, and triterpenes, that provide a great assortment of biological properties, such as blood lipid-lowering,3,4 blood sugar-lowering, 5 anti-inflammatory, 6 antitumor, 7 and immunoregulatory 5 effects.
Previous studies have shown that the characteristic components of P vulgaris, such as betulinic acid, ursolic acid, and their derivatives exerted antiestrogenic effects, which can be used as therapeutic agents against estrogen-dependent breast cancer. 8 Oleanolic acid and ursolic acid from P vulgaris can inhibit the production of nitric oxide (NO) and prostaglandin E2 (PGE2), the expression of NF-κB, and activate the expression of Nrf2, thereby exhibiting strong anti-inflammatory activity. 9 Ursolic acid inhibits the activation of NF-κB, which is induced by various carcinogens, through suppression of IκBα kinase and p65 phosphorylation. 10 In additional, ursolic acid induced apoptosis in human breast cancer cells through intrinsic and extrinsic apoptotic pathways. 11 Rosmarinic acid, a characteristic phenolic acid component of P vulgaris, can inhibit the expression of COX-2, thereby inhibiting the production of PGE2 and NO. 12
During the growth of plants, the contents of active substances are affected by many factors, such as the external environment, nutritional status and physiological state, and show certain fluctuations. At present, there are not enough research data on the pharmacological activity and material basis of P vulgaris during its growth and development. Therefore, the triterpenes and phenols of Prunellae Spica during developmental stages were separated and quantified in the present study. Anti-inflammatory and anti-breast cancer properties of Prunellae Spica at different developmental stages were investigated in vitro to guide the harvest of P vulgaris and improve its medical use. Moreover, Grey correlation and Pearson correlation analysis were used to screen the effective ingredients of Prunellae Spica, and a multidimensional network of “ingredient-target-pathway” through network pharmacology was constructed to explore the potential molecular mechanism of the active ingredients from Prunellae Spica in the treatment of inflammation and breast cancer. The data obtained provide a theoretical basis and practical information for the production of high-quality P vulgaris herbal materials, which can also help recognized the optimum time for herb harvest.
Results
Determination of the Major Components in Prunellae Spica During Development
To obtain detailed information on the phytochemicals, the characteristic peaks of triterpene and phenolic extracts from Prunellae Spica at different stages were determined by high performance liquid chromatography (HPLC). Betulinic acid, oleanolic acid, and ursolic acid were identified as the main triterpenoids in Prunellae Spica, and salvianic acid A, caffeic acid, salviaflaside, hyperoside, isoquercitrin, and rosmarinic acid were identified as the main phenolic components (Figure 1).
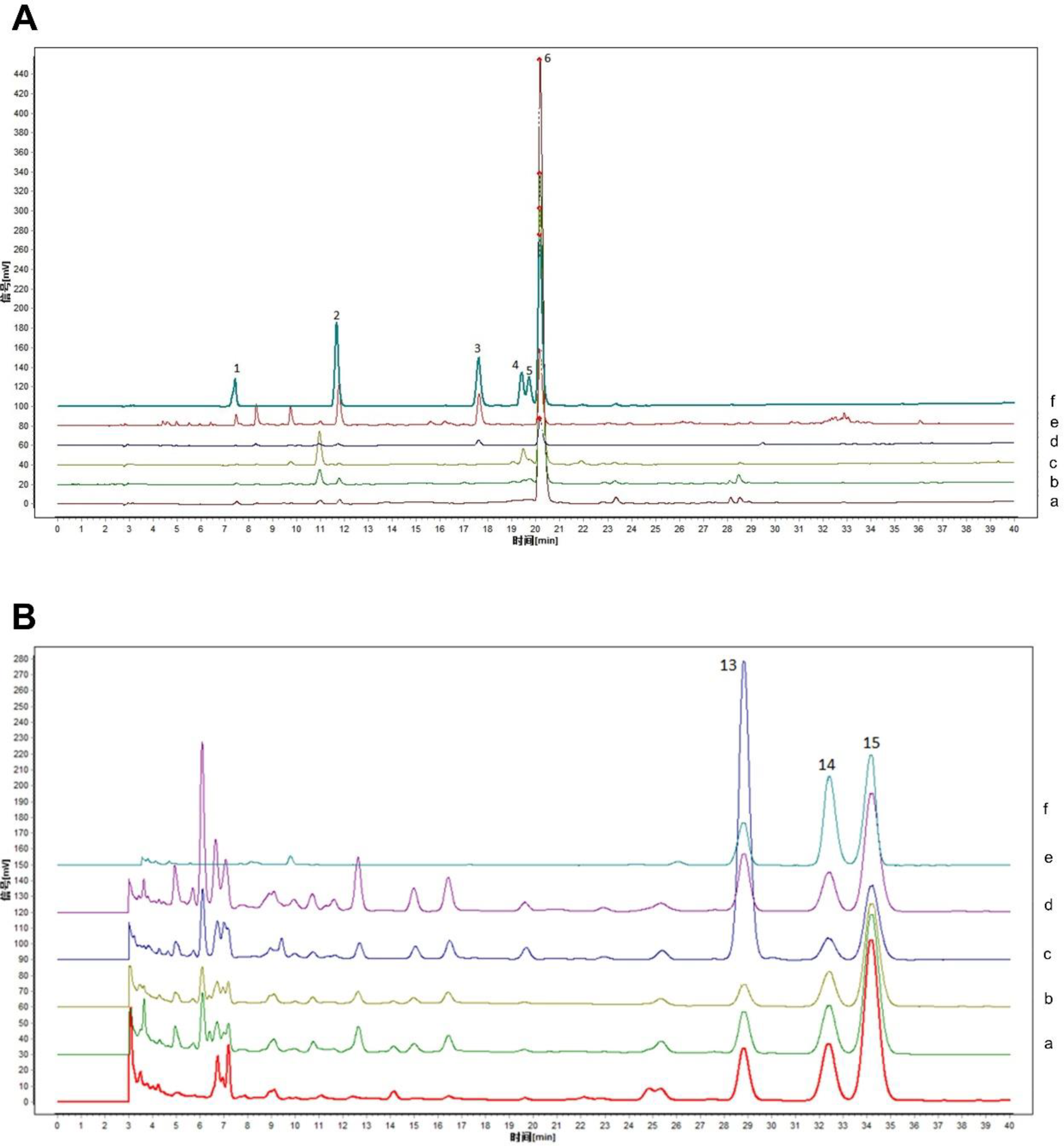
High performance liquid chromatography (HPLC) diagrams of Prunellae Spica extracts from different periods. (A) HPLC diagrams of phenolic extracts from Prunellae Spica at different stages and mixed standards. 1. salvianic acid A; 2. caffeic acid; 3. salviaflaside; 4. hyperoside; 5. isoquercitrin; 6. rosmarinic acid. Stage1 (a), stage2 (b), stage3 (c), stage4 (d), stage5 (e), and Mixed standard (f). (B) HPLC diagrams of triterpene extracts and mixed standards of Prunellae Spica from different periods. 13. betulinic acid; 14. oleanolic acid; 15. ursolic acid. Stage1 (a), stage2 (b), stage3 (c), stage4 (d), stage5 (e), and Mixed standard (f).
It can be seen that the content of betulinic acid gradually increased with the development of Prunellae Spica, while the contents of oleanolic acid and ursolic acid showed the opposite trend (Figure 2). Among the phenols, the contents of salvianic acid A, caffeic acid, and salviaflaside were the highest in stage 5 (Figure 3). The contents of hyperoside and isoquercitrin first increased and then decreased with the development of Prunellae Spica, while the content of rosmarinic acid decreased continuously (Figure 3).

Variation trend of triterpenoids in Prunellae Spica at different stages (

Variation trend of phenolic components in Prunellae Spica at different stages (
The Anti-Inflammatory Activities of Spica Prunellae at Development Stages
RAW 264.7 cells were treated with triterpenoids and phenoloids of Prunellae Spica at different concerntrations for 24 h, and the cytotoxicity was assessed using MTT assay kit. As shown in Figure 4, the triterpenoids and phenoloids of Prunellae Spica showed a certain dose-dependent inhibitory effect in the concentration range of 6.25 to 400 μg/mL, and the inhibitory effect in the later stages of development was relatively weak. Except for the extract during the heading period, Prunellae Spica extracts did not exhibit cytotoxity at 50 μg/mL. Based on these results, a concentration of 50 μg/mL was selected in the following experiments. In order to evaluate the anti-inflammatory properties, we investigated the effects of triterpenes and phenolic extracts of Prunellae Spica on NO production and IL-6 release in LPS-induced RAW264.7 cells. The increased pro-inflammatory mediators such as NO and IL-6 in LPS-induced macrophages were significantly blocked by pretreatment with both the triterpenes and phenolic extracts of Prunellae Spica at different periods (P < .05) (Figure 5A). These results suggest that a certain degree of anti-inflammatory activity was observed in the groups treated with the extracts of Prunellae Spica at all 5 stages, and the anti-inflammatory activities of Prunellae Spica in the later stages of development were stronger than that in the early stages (Figure 5B). In addition, the anti-inflammatory activities of phenolic extracts were stronger than that of triterpene extracts (Figure 5B).

The effects of triterpenes and phenols from Prunellae Spica at different stages on the proliferation of RAW 264.7 macrophages (

The anti-inflammatory activities of Spica Prunellae phenols and triterpenes at development stages. (A) The effects of phenols and triterpenes from Prunellae Spica at different stages on nitric oxide (NO) production and IL-6 release in RAW264.7 cells (
The Anti-Breast Cancer Activities of Prunellae Spica at Development Stages
The inhibitory effects of triterpene and phenolic extracts from Prunellae Spica on the growth of MDA-MB-231 cells were examined by MTT assay. IC50 is the median concentration of a substance that causes 50% inhibition. It was revealed that Prunellae Spica phenolics exerted antiproliferative effects on the MDA-MB-231 cell line and exhibited IC50 values of 256.23 ± 87.48, 354.63 ± 105.46, 549.28 ± 184.62, 660.55 ± 133.28, and 777.38 ± 114.76 μg/mL at different stages (Figure 6A left). However, the IC50 values of Prunellae Spica triterpenes at different stages toward MDA-MB-231 cell line were 109.57 ± 20.63, 225.05 ± 54.79, 277.28 ± 60.54, 409.85 ± 98.04, and 633.72 ± 172.36 μg/mL, respectively (Figure 6A right). However, it was found that with the development of Prunellae Spica, its antiproliferative activity gradually decreased (Figure 6B). The extracts had the strongest anti-breast cancer activity when taken from Prunellae Spica at the heading stage. The inhibitory effect of triterpene extracts on the growth of MDA-MB-231 cell line was stronger than that of phenolic extracts (Figure 6B).
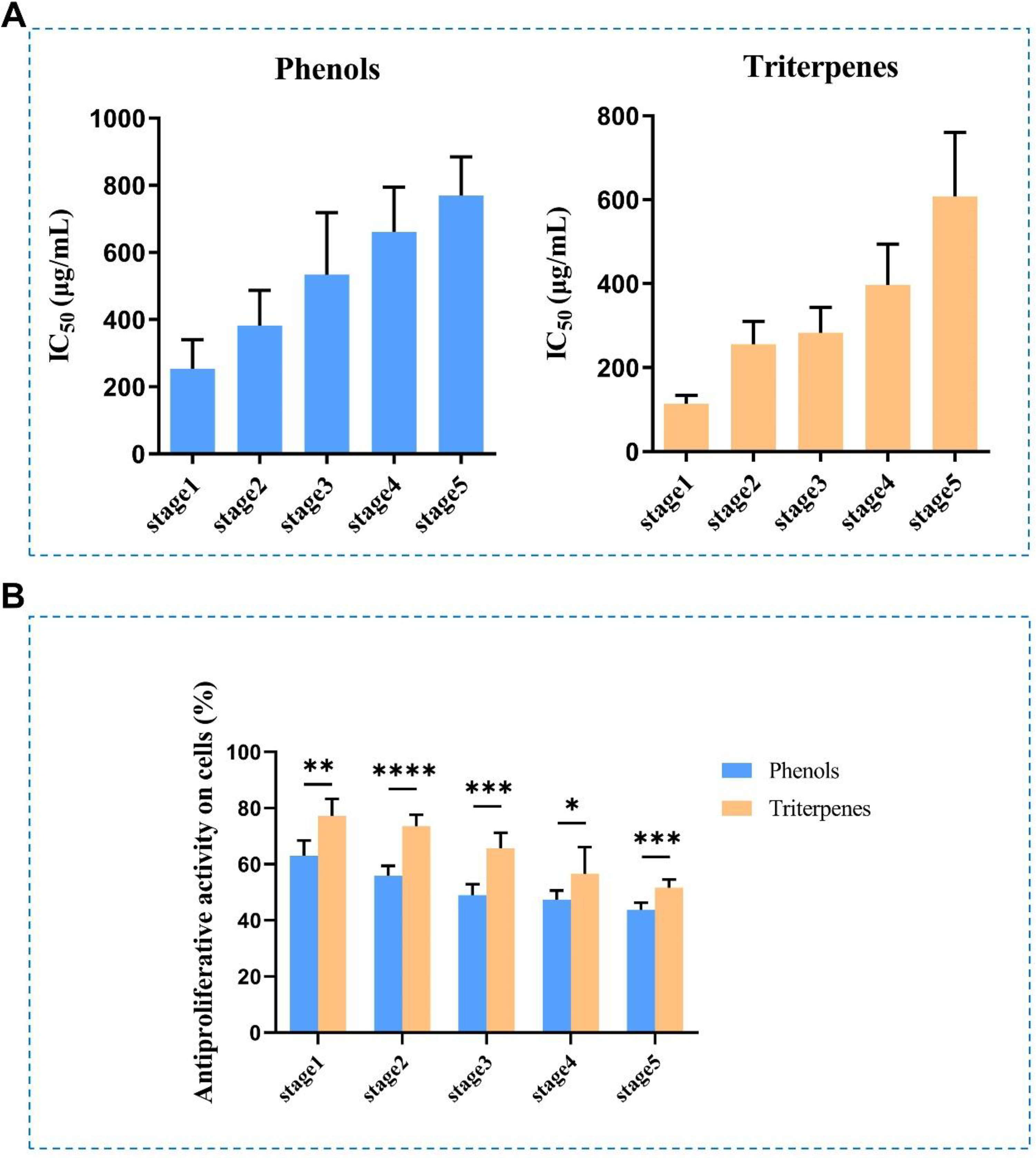
The anti-breast cancer activities of Prunellae Spica phenols and triterpenes at development stages (
Correlation Analysis between Components and Pharmacological Activities
Grey correlation analysis between the components and pharmacological activities was performed (Table 1). The correlations between compounds and anti-inflammatory activity were rosmarinic acid, salviaflaside, and hyperoside as the top 3. The correlations between compounds and anti-breast cancer activity were betulinic acid, ursolic acid, and oleanolic acid as the top 3.
Correlation of the Major Triterpenes and Phenols With the Inhibition Rates of NO and IL-6 Release or MDA-MB-231 Proliferation.

The Pearson correlation coefficients (r) among phenolic and triterpene content against anti-inflammatory and anti-breast cancer activities were performed (Figure 7). The correlation coefficient number was between −1 and 1. As the number of correlation coefficient obtained from linear relationship nears to 1, it indicates the stronger relationship between 2 variables. Meanwhile, positive (+) indicate the positive direction of relationship. It showed that, the salviaflaside content was positively correlated with anti-inflammatory activity of Prunellae Spica significantly (r2 = 0.391, P = .03), whereas positive correlation was observed between rosmarinic acid, oleanolic acid, ursolic acid, and isoquercitrin content corresponding to anti-breast cancer activity significantly (r 2 = 0.748, 0.542, 0.478, 0.439 and P = 2.06 × 10− 6 , .002, .007, .02, respectively).

Correlation cluster heatmap. The heatmaps quantify the compounds–activities relationships. (A) Pearson correlations between phenol contents and the inhibition rates of nitric oxide (NO) and IL-6 release or MDA-MB-231 proliferation. (B) Pearson correlations between triterpene content and the inhibition rates of NO and IL-6 release or MDA-MB-231 proliferation. P values <.05, .01 are marked with 1, 2 asterisks, respectively.
Comprehensive Grey correlation and Pearson correlation analysis, we speculate that salviaflaside is the mainly component responsible for the anti-inflammatory effect, while oleanolic acid and ursolic acid may be the components responsible for the anti-breast cancer activity of Prunellae Spica. According to the literature, the ability of oleanolic acid13,14 and ursolic acid15,16 on treating breast cancer was confirmed in previous studies. This is consistent with the conclusion of this study.
Network Pharmacology Analysis
The concept of network pharmacology was first proposed by Hopkins. 17 It combines network biology with polypharmacology, based on the poor efficacy of highly selective single-target drugs. 18 Through network pharmacology, we can directly identify drugs and disease targets from a large amount of data and understand the mechanisms and pathways between them. 19
In this study, through network pharmacology, 76, 71, and 19 targets that were strongly associated with the active ingredients oleanolic acid, ursolic acid, and salviaflaside, respectively, were obtained from the Swiss Target Prediction database (Probability > 0) (Figure 8A and Figure 9A). A total of 15195 targets related to breast cancer and 10920 targets related to inflammation were obtained from the Genecards database; 808 breast cancer–associated genes (relevance score > 20 points) screened from the GeneCards database were compared with the obtained oleanolic acid and ursolic acid–related targets, and 17 potential targets with anti-breast cancer effects were identified (Figure 8A). The obtained salviaflaside-related targets were compared with the inflammation-related targets, and 17 potential targets for anti-inflammatory effects were identified (Figure 9A).

Network pharmacology analysis of ursolic acid and oleanolic acid related to breast cancer. (A) Venn diagram of targets related to ursolic acid (red circle), oleanolic acid (green circle), and breast cancer (blue circle). (B) Protein–protein interaction (PPI) interaction network diagram of common targets of ursolic acid and oleanolic acid–breast cancer. (C) GO biological processes and a diagram showing their -lgP relationship with breast cancer. (D) KEGG signaling pathways and a bubble diagram showing their -lgQ relationship with breast cancer. (E) Ingredient-target-KEGG pathway multidimensional network. The yellow diamond–shaped nodes represent the disease (breast cancer), the red arrow–shaped nodes represent the components (ursolic acid and oleanolic acid), the blue square–shaped nodes represent the pathway, and the yellow circle–shaped nodes represent the targets corresponding to components, disease, and pathway.
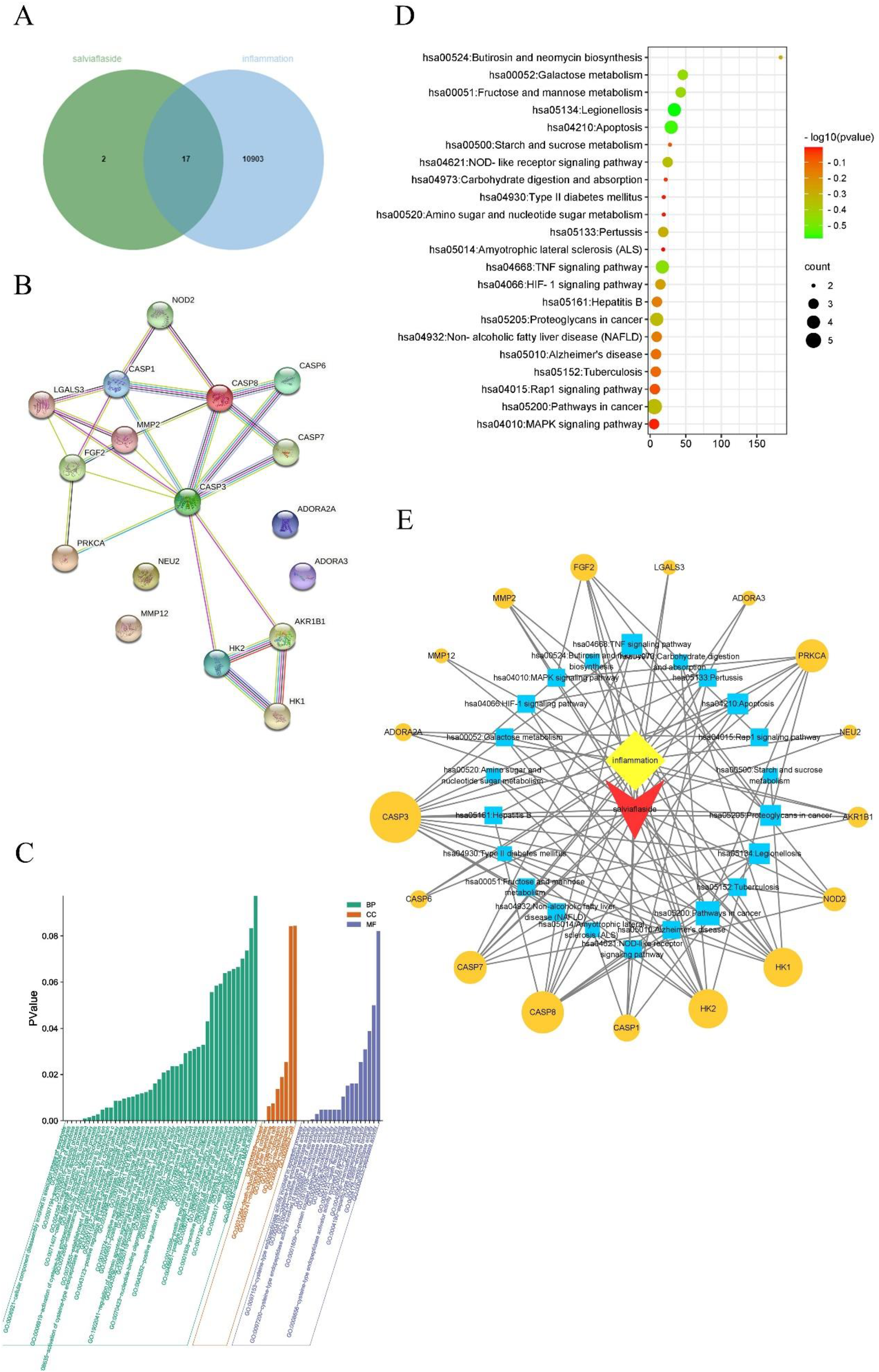
Network pharmacology analysis of salviaflaside related to inflammation. (A) Venn diagram of targets related to salviaflaside (green circle) and inflammation (blue circle). (B) Protein–protein interaction (PPI) interaction network diagram of common targets of salviaflaside-inflammation. (C) GO biological processes and a diagram showing their -lgP relationship with inflammation. (D) KEGG signaling pathways and a bubble diagram showing their -lgQ relationship with breast cancer. (E) Ingredient-target-KEGG pathway multidimensional network. The yellow diamond–shaped nodes represent the disease (inflammation), the red arrow–shaped nodes represent the compound (salviaflaside), the blue square–shaped nodes represent the pathway, and the yellow circle–shaped nodes represent the targets corresponding to components, disease and pathway.
The common targets were input into STRING online database to collect the protein–protein interaction (PPI) information so as to build this work. The network of oleanolic acid and ursolic acid breast cancer genes contains 17 nodes and 55 edges (Figure 8B). The network of salviaflaside-inflammation genes contains 17 nodes and 25 edges (Figure 9B).
Then, GO and KEGG enrichment analysis were performed. GO biological process (BP) consists of molecular function (MF), BP, and cellular component (CC) to interpret antitumor BPs at key targets. 20 The results of GO enrichment analysis showed that 42 BPs, 7 CC, and 23 MF were predicted related to breast cancer (Figure 8C). The BP of GO involves the transcription initiation from RNA polymerase II promoter, positive regulation of transcription from RNA polymerase II promoter, and chromosome segregation. In CC, protein complex, nucleoplasm, and nucleoplasm were highly correlated with breast cancer. In the MF category, most proteins were addressed to steroid binding, steroid hormone receptor activity, and enzyme binding. KEGG enrichment has 12 signaling pathways related to breast cancer (Figure 8D), and the pathways mainly involved in ovarian steroidogenesis, prolactin signaling pathway, progesterone-mediated oocyte maturation, and oocyte meiosis. There were 44 BPs, 8 CC, and 18 MF were predicted related to inflammation (Figure 9C). The BPs of GO were mainly involved in CC disassembly involved in execution phase of apoptosis, proteolysis, proteolysis, execution phase of apoptosis, and so on. The CCs of GO were mainly involved in cytosol, death-inducing signaling complex, and mitochondrial outer membrane. In the MF category, most proteins were involved in cysteine-type endopeptidase activity, cysteine-type peptidase activity, and fructokinase activity. Twenty-two pathways related to inflammation were predicted. The major pathways were TNF signaling pathway, NOD-like receptor signaling pathway, HIF-1 signaling pathway, Rap1 signaling pathway, and MAPK signaling pathway (Figure 9D).
The multidimensional network of “Component-Target Disease-KEGG pathway” was constructed by Cytoscape 3.8.2 software. We speculate that oleanolic acid and ursolic acid of Prunellae Spica may play a role in the anti-breast cancer effect by regulating Ovarian steroidogenesis, Prolactin signaling pathway and other signaling pathways on targets such as PTGS2, CYP19A1, ESR2, CYP17A1, and MAPK3 (Figure 8E). Salviaflaside in Prunellae Spica may regulate TNF signaling pathway, NOD-like receptor signaling pathway, and other signaling pathways by acting on targets such as CASP7, CASP8, CASP3, NOD2, and CASP1 to play a role in the anti-inflammatory effect of Prunellae Spica (Figure 9E). The results showed that the effective components from Prunellae Spica could treat breast cancer and inflammation by multitarget and multisignal pathways.
Discussion
Prunella vulgaris is a perennial herb that has a long history for use as a kind of medicinal and edible plant. Prunellae Spica is the dried fruit spike of P vulgaris. In China, Prunellae Spica, as the main ingredient of the well-known brand of herbal tea, has great market demand. Additionally, it has been used as a traditional medicine mostly in temperate regions of Europe and Asia, northwestern Africa, and North America for hundreds of years and is regarded as the medicinal part of P vulgaris in the Chinese Pharmacopoeia. Prunella vulgaris is the opposite of most plants, and it sinks into the shade. According to the theory of 5 fortunes and 6 qi, “Yang is born in the winter solstice, and yin is born in the summer solstice.” Prunella vulgaris withers when “yin is born,” that is, the trend from top to bottom. As a result, it can restrain the hyperactive yang of the liver and dispel the stagnation of wind and heat. In addition, P vulgaris mainly enters the Jueyin Liver Meridian. Traditional Chinese medicine believes that P vulgaris has the effects of clearing the liver and improving eyesight, dispersing knots and detoxifying, treating headache, dizziness, tinnitus, scrofula, gall tumor, breast addiction, breast carbuncle, rock-breast, mumps, carbuncle furuncle swollen poison, acute and chronic hepatitis, and hypertension. Among them, “scrofula” is a chronic suppurative disease that occurs in the neck. The mastitis, mumps, and carbuncle furuncle swollen poison refer to mastitis, mumps, and purulent inflammation, respectively. It can be seen that anti-inflammatory is one of the main pharmacological activities of P vulgaris, which is likely to be one of the reasons for P vulgaris harvest after withering.
(Ming Dynasty) Xiyong Miao's pharmacological work “Ben Cao Jing Shu” records: “P. vulgaris…Get dandelion grass to treat all breast carbuncle (Ru-yong) and rock-breast (Ru-yan) Fang….” 21 Ruanjian Sanjie (RJSJ) decoction is composed of 4 herbs, including Pinellia ternata, Prunella vulgaris, Cremastra appendiculata, and Sargassum pallidum, and has traditionally been used for softening hard lumps and resolving hard tissue masses, which shows potent cytotoxicity against breast cancer cells in vitro. 22 Besides, P vulgaris is contained in Chinese patent medicines such as Rupi Sanjie Capsule, Xiaoru Sanjie Capsule, Rukang Soft Capsule, and Ruhe Neixiao Capsule, which are clinically used for the treatment of breast diseases. Modern medical research shows that, P vulgaris may be a potential adjuvant medicine for breast cancer treatment. 23
Prunella vulgaris contains triterpenoids and their saponins, phenolic acids, sterols and associated glycosides, flavonoids, organic acids, volatile oil, and saccharides. 1 Flavonoids, phenolic acids, stilbenes, and tannins (hydrolysable and condensed) are a class of important secondary metabolites with multiple phenolic hydroxyl groups, 24 which are mainly synthesized by the phenylpropanoid metabolic pathway. 25 In the present study, the contents of rosmarinic acid, ursolic acid, and oleanolic acid in Prunellae Spica basically decreased, which was contrary to the change trend of betulinic acid content. The contents of caffeic acid, salvianic acid A, and salviaflaside had similar trends of change, and they all accumulated in a large amount during P vulgaris ripening. It is possible that the developmental stage of the plant affects the chemical composition of the species. Chen et al investigated the changes in the bioactive compounds present in P vulgaris spicas and the whole plant at 5 harvest times over 2 months found that the highest levels of RA, UA, and OA were observed at the flowering stage, whereas the lowest levels of RA, UA, and OA were observed at the mature-fruiting stage.26,27 Similar results were obtained for cardoon heads harvested during the flowering season 28 and for globe artichoke bracts, floral stems, and receptacles, where the highest phenolic content was detected in samples collected during the spring. 29 Ayan et al reported that the highest level of total phenol content was at full-flowering in Hypericum pruinatum. 30 Additionally, Verma and Kasera indicated that the peak concentration of phenols was observed at the flowering stage in Boerhavia diffusa. 31 These are basically consistent with our findings in the present study. Our previous studies reported that 32 terpenoids were mainly synthesized in the maturity stage of Prunellae Spica, with the key enzymes and coding genes in downstream pathways being mainly expressed during ripening. Flavonoids mainly accumulated before ripening, with highly expressed pathway enzymes and coding genes. The accumulation of phenylpropanoids was relatively active throughout the development process. It may be related to the important function of triterpenes and phenolic on plant defense mechanisms. 33
Caffeic acid belongs to the class of phenylpropionic acids 34 and is the basic structural unit of phenolic acids. 35 The precursor compound of caffeic acid is phenylalanine, which is also a precursor compound of rosmarinic acid. 36 As one of the chemical markers for the quality evaluation of P vulgaris, rosmarinic acid is the core constituent unit of a series of complex phenolic acids,37,38 which has some bioactive glycoside derivatives, such as salviaflaside (rosmarinic acid-3-O-glucoside). 39 The synthesis of rosmarinic acid has great significance for the formation of complex phenolic compounds. 40 Salvianic acid A is also a basic component of caffeic acid derivatives in plant metabolites and the hydrolysate of caffeic acid. 41 In the present study, salvianic acid A, caffeic acid, and salviaflaside mainly accumulated in late development, while rosmarinic acid showed the opposite. It means that there is transformation between these components. During the development of Prunellae Spica, the seeds form in stage 3 and mature gradually. Salviaflaside is a specific component in P vulgaris. 42 The accumulation of salviaflaside started at stage 3, and its content gradually increased with the maturity of the seed, leading to the gradual decrease of its precursor rosmarinic acid. According to the literatures, salviaflaside exhibited effective anti-inflammatory, antiviral, and antioxidation activities in mice caused by H1N1 virus, 43 as well as anti-breast cancer activity through inducing the apoptosis of MDA-MB-231 cells. 44 NLRP3, the most well studied Nod-like receptor, forms a complex comprised of adaptor proteins, such as the serine protease caspase-1 (CASP1). 45 NLRP3 inflammasome activation governs the cleavage and activation of CASP1, a cysteine protease responsible for the processing and secretion of pro-interleukin IL-1β and IL-18, which are closely related to the induction of inflammation.46,47 Ursolic acid could target CASP3, alleviate inflammation-associated downstream signal transduction factors and exhibited anti-inflammation activities. 48 Whether salviaflaside mediated this mechanism to exert anti-inflammatory effect in P vulgaris needs further verification.
In the present study, oleanolic acid and ursolic acid of Prunellae Spica may play a role in the anti-breast cancer effect by regulating genes, such as PTGS2, CYP19A1, ESR2, CYP17A1, and MAPK3. Inducible cyclooxygenase (COX2; also known as PTGS2) have essential roles in the physiological response to tissue injury, but excessive expression of these inflammatory enzymes drives carcinogenesis. 49 COX2 is implicated in cancer cell growth. 50 Ursolic acid treatment could cause a marked decrease in COX2 and IL-6 mRNA expression in tumor-induced mice. 51 The derivatives of oleanolic acid (SZC014) treatment could significantly inhibit the COX2 expression in MCF-7 cells. 50 CYP19A1 upregulation in peripheral tissues is associated with familial incidences of cancer, breast cancer in particular. 52 Estrogen is a product of the CYP19A1 gene. 53 Because estrogen signaling is crucial for breast cancer cell growth, strategies designed to suppress estrogen signaling have been used to treat breast cancer patients. A recent study demonstrated in vitro and in vivo antiestrogenic effects of P vulgaris extracts. 54 Ursolic acid as antiestrogenic compounds responsible for the antiestrogenic effects of P vulgaris, which suppressed estrogen receptor–mediated signaling through inhibition of estrogen receptor synthesis at the transcriptional level. 8
In conclusion, our data showed that, during its development, Prunellae Spica has a dual therapeutic effect, exerting anti-breast cancer in its early stages and anti-inflammatory in its late stages. Therefore, the harvest period can be selected according to the desired effect of Prunellae Spica. For example, Prunellae Spica harvested at the turning and ripening stages could be used an anti-inflammatory drug, and the plant could be harvested at the heading stage for use as an anti-breast cancer drug. The anti-inflammatory and anti-breast cancer effects of Prunellae Spica result from multiple components, multiple targets, and multiple pathways. These findings provide new ideas and methods for further research on the material basis and mechanism of the anti-inflammatory and anti-breast cancer effects of P vulgaris. They also provide a reference for determining the appropriate harvest period and the rational development and utilization of P vulgaris.
Materials and Methods
Plant Material
Prunella vulgaris was identified by Professor Limin Gong, School of Pharmacy, Hunan University of Traditional Chinese Medicine. The test site was the Pharmaceutical Plantation Garden of Hunan University of Traditional Chinese Medicine, at the location with geographical coordinates 112°54 E, 28°08 N, altitude 88 m above sea level, subtropical monsoon climate, and sandy loam soil. Organic compound fertilizer (1.2 × 103 kg/hm) was applied to the soil once as a base fertilizer before planting. The annual precipitation was 1361.6 mm, the annual average temperature was 17.2 °C, and the annual sunshine was 1529.3 h. Prunellae Spica samples were collected from April 15, 2019, to June 16, 2019, at the heading stage (stage 1), flowering stage (stage 2), green fruit stage (stage 3), turning stage (stage 4), and ripening stage (stage 5) (Table 2).
Morphology and Sample Information of Prunellae Spica at Different Developmental Stages.

Standards and Reagents
Ursolic acid, oleanolic acid, betulinic acid, salvianic acid A, caffeic acid, salviaflaside, hypericin, isoquercitrin, and rosmarinic acid standard products (purity > 98%) were purchased from Vikki Biotechnology Co., Ltd. Methanol and acetonitrile were chromatographically pure and purchased from Merck, Germany. Phosphoric acid was chromatographically pure and purchased from Tianjin Institute of Chemical Reagents. DMEM was purchased from HyClone. Phosphate-buffered saline (PBS) was purchased from Proceeding Company. Fetal bovine serum was purchased from Biological Industries, Israel. A kit for measuring NO levels was purchased from Biyuntian Company. A kit for measuring IL-6 levels was purchased from Wuhan Huamei Company.
Cells and Culture
Both RAW 264.7 macrophages and MDA-MB-231 cells were purchased from Wuhan Pu Nuo Sai Life Technology Co., Ltd. RAW264.7 cells were cultured in DMEM containing 10% fetal bovine serum, and MDA-MB-231 cells were cultured in DMEM containing 10% fetal bovine serum and a 1% penicillin–streptomycin mixture. The cells were placed in a constant temperature incubator at 37 °C with 5% CO2. The adherence of the cells was observed, and the medium was changed the next day. RAW 264.7 cells that showed good adherent growth were digested with ice-cold PBS, MDA-MB-231 cells were digested with 0.25% trypsin, and logarithmic growth phase cells were selected for the experiment.
Preparation of Standard Solutions and Sample Solutions
A total of 1.065, 1.425, and 1.86 mg of betulinic acid, oleanolic acid, and ursolic acid, respectively, were accurately weighed for use as reference substances, and methanol was added to prepare reference substance stock solutions. Appropriate amounts of the above 3 reference substance stock solutions were precisely mixed, and methanol was added to prepare mixed reference solutions with mass concentrations of 0.355, 0.475, and 0.62 mg/mL. A total of 4.71, 2.04, 5.24, 4.56, 3.51, and 4.79 mg of salvianic acid A, caffeic acid, salviaflaside, hyperoside, isoquercitrin, and rosmarinic acid, respectively, were weighed as reference substances, and methanol was added to prepare a control stock solution. Appropriate amounts of the above 6 reference stock solutions were precisely mixed, and methanol was added to prepare mixed reference solutions with mass concentrations of 0.471, 0.204, 0.524, 0.456, 0.351, and 0.479 g/L.
One gram of Prunellae Spica powder was accurately weighed, and then 25 mL of methanol was added to make a mixture and ultrasonicated for 1 h. The supernatant was concentrated to the extract at 60 °C, washed twice with 8 mL of petroleum ether, and then dissolved in 4 mL of 20% methanol. Four milliliters of the solution was transferred to an activated C18 SPE cartridge (500/3 mL SPE solid phase extraction cartridge, purchased from Shenzhen Comma Biotechnology Co., Ltd.) and eluted with 1 mL of 20% methanol and 3 mL of 50% methanol to collect the phenolic sample solution. Finally, 5 mL of methanol was added for elution to collect the triterpene sample solution.
High Performance Liquid Chromatography Conditions
Triterpene
The HPLC analysis was performed on an Agilent HPLC system equipped with a Kromasil 100-5C18 analytical column (250 × 4.6 mm, 5 μm) at 30 °C. The mobile phase consisted of A (methanol)/B (0.1% phosphoric acid) (85:15, v/v) with a flow rate of 1 mL·min−1, and the detection wavelength was set at 210 nm. The injection volume was 10 μL.
Phenols
The HPLC analysis was performed on an Agilent HPLC system equipped with an Agilent Zorbax SB-C18 column (250 × 4.6 mm, 5 μm) at 30 °C. The elution was performed with solvents A (methanol) and B (0.1% glacial acetic acid). Separation was initiated using the following gradient: 0 to 5 min, 5% to 30%; 5 to 10 min, 30% to 35%; 10 to 20 min, 35% to 48%; 20 to 25 min, 48% to 60%; 25 to 30 min, 60% to 80%; and 30 to 40 min, 80% to 100%. The flow rate was 1 mL/min, and the detection wavelength was set at 280 nm. The injection volume was 10 μL.
Method Validation and Quantitative Determination
The method was validated with respect to precision, stability, repeatability, linearity, and accuracy (recovery). 1, 2, 4, 6, 8, and 10 μL of the 3 triterpene and 6 phenols mixed reference solutions were analyzed, and the linearity of the injection volume (X) with the peak area (Y) was determined. The equations of the calibration curve for the ingredients are shown in Table 3. The contents of the individual samples were calculated accordingly. The high-correlation coefficient values (r > 0.9995) obtained indicated that there were good linear correlations between the concentrations of the relative standard deviations (RSDs) of investigated compounds and their peak areas within the test concentrations. In addition, the precision, repeatability, and stability for each compound were less than 3.0%.
Quantitation of the 9 Components Under Optimized HPLC Conditions.

A total of 1.00 g of Prunellae Spica powder (S4) with a known content was weighed (6 parts in total), and appropriate amounts of betulinic acid, oleanolic acid, and ursolic acid reference solutions were added to prepare a test solution according to the method of preparation. Based on the peak area of each triterpene compound, the sample recovery and RSDs of betulinic acid, oleanolic acid, and ursolic acid were calculated to verify the accuracy of the method. One gram of Prunellae Spica powder (S4) with a known content was weighed (6 parts in total), and an appropriate amount of salvianic acid A reference solution was added. One gram of Prunellae Spica powder (S19) with a known content (6 parts in total) was weighed; coffee acid, salviaflaside, hyperoside, isoquercitrin, and rosmarinic acid reference solutions were added to prepare a test solution according to the preparation method; and the peak area of each phenol was measured. The sample recovery rates and RSDs of salvianic acid A, caffeic acid, salviaflaside, hyperoside, isoquercitrin, and rosmarinic acid were used to verify the accuracy of the method. The developed method showed good recoveries at a range of 97.06% to 102.92%, indicated that the HPLC method was efficient, accurate, and sensitive for quantitative determination of the major chemical markers in Prunellae Spica.
Effect on Viability of RAW 264.7 Cells
After rinsing with PBS 3 times, RAW 264.7 cells in logarithmic growth phase were added to 2 mL of precooled PBS, and a pipette was used to collect the cells. The cell density was adjusted to 1 × 104 cells/well, and the cells were plated in 96-well plates and incubated overnight at 37 °C and 5% CO2. Prunellae Spica triterpenes and phenolic extracts from different stages were added at final concentrations of 3.13, 6.25, 12.5, 25, 50, 100, 200, and 400 μg/mL, and the control group, positive drug group (50 μg/mL of dexamethasone), and LPS model group (1 μg/mL) were set up at the same time, with 4 replicate wells for each concentration. After incubation for 24 h, 100 μL MTT (0.5 mg/mL) was added to each well, and the cells were incubated at 37 °C for 4 h. Then, the supernatant was discarded, 150 μL DMSO was added to each well, and the plate was shaken for 10 min. The absorbance (A) value was measured at 490 nm. The control group consisted of untreated cells was considered as 100% of viable cells. Results are expressed as percentage of viable cells when compared with control groups.
Effect on LPS-Induced NO Production and IL-6 Release
After rinsing with PBS 3 times, RAW 264.7 cells in logarithmic growth phase were added to 2 mL of precooled PBS, and a pipette was used to collect the cells. The cell density was adjusted to 1 × 104 cells/well, and the cells were plated in 96-well plates and incubated overnight at 37 °C and 5% CO2. Prunellae Spica triterpenes and phenolic extracts from different stages were added at a final concentration of 50 μg/mL, and the control group, positive drug group (50 μg/mL of dexamethasone), and LPS model group (1 μg/mL) were set up at the same time, with 4 replicate wells for each concentration. After incubation for 2 h, the cells were stimulated with LPS at a concentration of 1 μg/mL, and the supernatant was aspirated after 24 h. According to the instructions of the NO and IL-6 detection kits, the absorbance value was measured to calculate NO and IL-6 content. There were 3 replicate wells for each group.
Antiproliferative Activity Against MDA-MB-231 Cells
The density of MDA-MB-231 cells in logarithmic growth phase was adjusted to 8 × 103 cells/well, and the cells were plated in 96-well plates and incubated overnight at 37 °C and 5% CO2. Triterpene or phenolic extracts of Prunellae Spica from different periods were added at final concentrations of 15.625, 31.25, 62.5, 125, 250, and 500 μg/mL. After culturing for 24 h, 100 μL MTT reagent (0.5 mg/mL) was added to each well, and the cells were incubated at 37 °C for 4 h. Then, the supernatant was discarded, and 150 μL DMSO was added to each well. After shaking for 10 min, the absorbance (A) value was measured at 490 nm with a microplate reader. The antiproliferative activity was calculated according to the formula, antiproliferative activity (%) = (1-A administration/A control) × 100%.
Correlation Analysis
The Pearson correlation between triterpene, phenol contents, and anti-inflammatory, anti-breast cancer activities of Prunellae Spica in different stages were plotted and the correlation coefficient (r) calculated by using SPSS 25.0. The results were visualized as a heat map using R version 3.6.1.
The grey correlation analysis method was used to calculate the correlation between triterpene, phenolic components, and anti-inflammatory, anti-breast cancer activities of Prunellae Spica in different stages. First, the original data were transformed by the averaging method. The transformed mother sequence was denoted as {X0(t)}, and the subsequence was denoted as {Xi(t)}. The pharmacodynamic indexes of the triterpene and phenolic extracts of Prunellae Spica in each period were used as the parent sequence, and the content of the corresponding characteristic component was used as the subsequence.
When t = k (k was the peak number), the parent sequence was denoted as {X0(k)}, the subsequence was denoted as {Xi(k)}, and the absolute difference between the parent sequence and the subsequence was Δ
0i
(k)=|X0(k) - Xi(k)|. The correlation coefficient η(k) between the parent sequence and the subsequence at t = k was calculated according to the formula (Ⅰ):
According to the formula (Ⅱ), the correlation coefficient corresponding to each subsequence was averaged, and ri was obtained as the correlation degree between the subsequence and the parent sequence:
Network Pharmacology Analysis
The active ingredients were entered into the Swiss Target Prediction database (http://www.swisstargetprediction.ch) for the targets directly related to each ingredient with the species limited to “Homo sapiens.” 55 Genecards database (https://www.genecards.org/) was utilized to collect the inflammation and breast cancer–related disease genes and targets by using “inflammation” and “breast cancer” as keywords, respectively.56,57 The common targets of active ingredient–related targets and inflammation or breast cancer–related targets were identified and imported into STRING (https://www.string-db.org/) for PPI. Cytoscape 3.8.2 was applied for the network visualization and network analysis. 58 The KEGG pathway and GO (Gene Ontology) enrichment analysis of the common target interaction network were performed by using DAVID (https://david-d.ncifcrf.gov,ver.6.8) database. 59 Enrichment GO term and enrichment dot bubble were plotted by http://www.bioinformatics.com.cn, a free online platform for data analysis and visualization.
Statistical Analysis
SPSS 25.0 software was used for statistical analysis, and comparisons between groups were performed by one-way variance analysis. The results are expressed as `x ± s, and P < .05 was considered statistically significant.
Footnotes
Acknowledgments
The authors would like to thank Hunan University of Chinese Medicine for the financial support. The authors would also like to express our sincere gratitude to Prof Limei Lin for her assistance in proofreading the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by “Outstanding Youth Program of Hunan Provincial Department of Education” (Grant No: 21B0397), “The Natural Science Foundation of Hunan Province” (Grant No: 2023JJ60128), “Hunan Province Science and Technology Innovation Leading Talent Project” (Grant No: 2021RC4034), “Hunan Science and Technology Innovation Team Project” (Grant No:2021RC4064), “Hunan Province Traditional Chinese Medicine Research Program” (Grant No: 2021054), “Scientific research project of Hunan Provincial Public Health and Family Planning Commission” (Grant No: 202113051481), “Changsha Natural Science Foundation Project” (Grant No: kq2202268). This project was financially supported by the First-class Discipline Project on traditional Chinese medicine of Hunan University of Chinese Medicine (No. 201803).