The importance of nitriles as a key class of chemicals with applications across the sciences is widely appreciated. The natural world is an underappreciated source of chemically diverse nitriles. With this in mind, this review describes novel nitrile-containing molecules isolated from natural sources from 1998 to 2021, as well as a discussion of the biological activity of these compounds. This study gathers 192 molecules from varied origins across the plant, animal, and microbial worlds. Their biological activity is extremely diverse, with many potential medicinal applications.
Nitriles are common organic compounds, characterized by the presence of one or more nitrile functional groups, comprising a carbon atom linked by a triple bond to a nitrogen atom, which can result in polar compounds with high dielectric constants. Although more frequently found in organic compounds, the functional group is also present in some inorganic compounds, as cyanides.1 Structurally related functionalities include isonitriles, cyanates, and thiocyanates. Nitriles are versatile compounds with applications across different fields of chemistry. Many nitriles are used in organic chemistry, including as solvents and synthons. Acetonitrile, the smallest organic nitrile, is widely used in reverse phase chromatography applications,2 or as an organic solvent.3 Polymer chemistry has afforded several important nitrile-containing polymers, including polyacrylonitrile, poly(acrylonitrile-co-butadiene-co-styrene), poly(styrene-co-acrylonitrile), and nitrile rubber. Such polymers can be functionalized to provide new macromolecules with interesting properties.4 Nitrile rubber, for example, is a widely used replacement for latex in laboratory gloves. The pharmaceutical industry is a major user of nitriles. Many pharmaceuticals, including those in clinical development, contain the nitrile functional group. Seven roles have been identified for the nitrile moiety; namely as (1) a carbonyl bioisostere, (2) a hydroxyl and carboxyl surrogate, (3) an electron-withdrawing group, (4) an azomethine-water isostere (for cyanoquinolines and cyanopyridines), (5) a carboxyl transition state analogue, (6) a halogen bioisostere, and (7) an improver of ADME-toxicology profiles.5 Given the diversity of the biological roles of the nitrile moiety in drugs, these compounds are encountered in prescriptions for a large variety of medicinal indications. A recent review discusses more than 30 FDA-approved nitrile-containing pharmaceuticals; citing advantageous properties associated with the functional group, including enhanced binding affinity, improved pharmacokinetic profile, and reduced drug resistance.6 Examples (Figure 1) include anastrozole 1, used to treat estrogen-dependent breast cancer, verapamil 2, an antiarrhythmic agent used against angina, and tofacitinib 3, a Janus kinase inhibitor used to treat rheumatoid arthritis, psoriatic arthritis, and ulcerative colitis.
Examples of pharmaceutically important nitriles.
As stated, the nitrile group acts as a bioisostere of other functional groups, including carbonyl and halogens. Incorporation of a nitrile functionality into drug molecules can block metabolically labile sites, thus increasing their metabolic stability.7 The bioactivity and toxicity of nitriles highly depend on the compound considered. Exposure to toxic nitriles can lead to neurologic, hepatic, cardiovascular, renal, or gastrointestinal disorders.8 An important hazard is seen in cyanogenic compounds, capable of releasing hydrogen cyanide (HCN). Many natural nitriles exist as cyanogenic glycosides, having been reported from more than 2500 plant species, including important food sources. Such compounds have the ability to liberate HCN following ingestion and metabolism, and are therefore potentially hazardous.9
Interest in natural nitriles has been growing significantly since the discovery of the first examples, and a number. of reviews about such nitriles exist in the literature. Nitrile-bearing molecules are found across Nature, with examples in plants, insects, fungi, bacteria, sponges, and other aquatic organisms. An early review from 1981 discussed compounds bearing cyano or isocyano functionalities occurring in plants, animals, and fungi, based on 60 reference sources.10 A later publication, from 1990, gathered the available information about nitriles in natural products at that time. This study listed all known molecules as well as describing the biosynthesis of nitriles and their degradation and detoxification pathways.11 A further publication, from 1999, continued the work of the earlier reviews, collecting all nitrile natural products discovered between 1990 and 1999 and categorizing them according to the hybridization of the nitrile-bearing carbon.12 At that time, over 120 natural products were known to contain nitriles, a number that did not include the cyanogenic glycosides. In fact, a 2021 review of cyanogenic glycosides listed 112 cyanogenic glycosides reported to date.13 To continue previous works, this paper aims to discuss natural products containing at least 1 nitrile functional group discovered between 1998 and 2021. Compounds bearing the isomeric isonitrile functional group are not included in this review; the reader is referred to a recent comprehensive review.14 In addition, labile natural products such as the glucosinolates, which readily convert to other constituents, including nitriles, are also not within the scope of this article. Again, these have been the subject of recent review.15
Methodology
The databases used for this review were SciFinder, PubMed, and Google Scholar. To obtain relevant references, the keywords “nitrile” and/or “cyanide” and “natural products” were used, combined with criteria regarding language and time range. Two languages were included; English and French, and the time range covered 1998 to 2021, to ensure the continuation of the scope of previous papers. Articles obtained using these criteria were sorted to retain all those that contained at least one nitrile natural product that was not already recorded in the older papers. Analysis of the listed compounds consisted of gathering their names, chemical structures, biological properties, and the organism from which they originated.
Results and Discussion
Alkyl Nitriles
A group of 19 long-chain aliphatic nitriles, 4 to 22, representing a new class of natural products featuring unbranched or methyl-branched unsaturated nitriles with an ω-7 or C-3 double bond (Figure 2), were identified in the headspace of liquid and agar cultures of Pseudomonas veronii R02 and Micromonospora echinospora DSMZ 43816.16 Some of these nitriles showed antimicrobial activity; for example, minimum inhibitory concentration (MIC) values of 2 to 8 μg/mL were seen for 20 (3-pentadecenenitrile) against various strains of Staphylococcus aureus, including multiresistant strains. Compound 13 ((E)-12-methyltridec-3-enenitrile) was active against the fungus Mucor hiemalis. No appreciable cytotoxic effects were seen among these nitriles when tested on human HCT-116 (colon carcinoma) cells.
Alkyl nitriles from Pseudomonas veronii & Micromonospora echinospora.
Sponges in the genus Mycale contain various alkyl nitriles (Figure 3). Mycale microsigmatosa (Mycalidae), collected in Venezuela, yielded 5-(19’-cyanononadecanyl)pyrrole-2-carboxaldehyde 23. When tested for activity against Leishmania mexicana promastigotes, 23 had an LD50 of 12 μg/mL.17Mycale tenuispiculata Dendy (Mycalidae) (IIC-309), collected in Indian waters, afforded (6’Z)-5-(23’-cyano-6’-tricosenyl)pyrrole-2-carboxaldehyde 24.18 Three further mycalenitriles were isolated from Mycale cecilia collected in the Gulf of California. The compounds were evaluated for their cytotoxicity using a panel of cancer cell lines, with compounds 25 to 27 having GI50 values ranging from 2.4 to 4.8 μg/mL in PANC1 (pancreatic carcinoma), LOVO (colon adenocarcinoma), and HeLa (cervix epithelial adenocarcinoma) cells.19 Bioassay-guided isolation of a lipid extract from a Mycale sp. from Palau afforded various 5-alkylpyrrole-2-carbaldehyde metabolites, including fourteen mycalenitriles 28 to 41. Compounds were shown to inhibit hypoxia-inducible factor 1 (HIF-1) activation in T47D (breast carcinoma) cells, with the most active compounds (30 and 31) having IC50 values of 7.8 μM and 8.6 μM, respectively.20 It was noted that while a highly lipophilic alkyl side chain was essential for the observed HIF-1 inhibitory activity, extremely hydrophobic unsubstituted long-chain analogues were nearly inactive, highlighting the importance of the nitrile functionality in modulating activity. Compounds 30 and 31 were also shown to suppress cell respiration at the mitochondrial electron transport chain (ETC) complex I in a concentration-dependent manner (1-30 μM, with an IC50 < 10 μM).
Alkyl nitriles from Mycale spp.
Albanitriles A-G, 42 to 48, were isolated from a Mycale sp. SS5 sample, (Mycalidae) WAM Z35806, (Figure 4).21 Compounds 42 to 48 did not exhibit significant anti-bacterial or anti-fungal properties when tested against Bacillus subtilis, Escherichia coli, or Candida albicans, but did show moderate anti-protozoal activity against Giardia duodenalis, with 42 (octadeca-6,8,10-triynedinitrile) having activity at a minimum concentration of 12 μM, comparable to the positive control metronidazole (2.9 μM). Within the compounds tested, anti-protozoal activity appeared to be dependent on alkyl chain length. No cytotoxicity was noted and the compounds were not considered to be cyanogenic.
Albanitriles A-G.
A cyanohydrin-containing phosphonate, (cyano(hydroxy)methyl)phosphonic acid, 49 (Figure 5), was isolated from Streptomyces regensis strain WC-3744.22 The molecule represents the first naturally occurring, cyanohydrin-containing phosphonate to be isolated and characterized. The cyanohydrin of glyoxylic acid, 50 (Figure 5) was identified as the precursor of hydrocyanic acid in the fungus Marasmius oreades (Bolton) Fr., where it acts as a wound-activated chemical defence.23 (R)-3-Hydroxybutanonitrile 51 (Figure 5), was isolated from the culture of Aspergillus sp. KJ-9, a fungal endophyte isolated from the stem bark of Melia azedarach L. (Meliaceae).24 When screened against various phytopathogenic fungi (Gibberella saubinetti, Magnaporthe grisea, Botrytis cinerea, Colletotrichum gloeosporioides, and Alternaria solani), 51 showed MIC ranges of 6.25 to 50 μM, compared to the positive controls carbendazim and hymexazol. When assessed for antibacterial activity against Bacillus cereus and S aureus, 51 had a MIC value of 50 μM against both organisms, compared with the positive controls streptomycin sulfate (MIC 1.57 and 12.5 μM respectively) and ampicillin (MIC 3.13 and 1.57 μM, respectively), while 51 was inactive against E coli and B subtilis. Based on comparison of the GC-MS output from a methanolic extract of Mukia maderaspatana (L.) M.Roem. (Cucurbitaceae), an Indian traditional medicine collected in the dry deciduous forests of Tamil Nadu, India, with the aid of spectral libraries, compound 52, a cyanoacetamide derivative (Figure 5) was reported.25
Oxygenated alkyl nitriles.
Amino Nitriles
These nitriles are characterized by having an amino functionality in the alpha (or more unusually, beta) position with respect to the nitrile carbon. The phylum Porifera comprises various aquatic sponges. Renieramycins are marine alkaloids belonging to the broader saframycin family, and are classified structurally as 1,2,3,4-tetrahydroisoquinoline-quinone derivatives.26 Of the various renieramycins isolated to date, several examples stabilised with a nitrile functionality (vide infra) have been isolated from sponges in the genera Xestospongia and Cribrochalina. Renieramycins 53 to 65 (with lettered designations) bear a nitrile (Figure 6). These compounds have interesting cytotoxic activity and their potential as anticancer drugs was recently reviewed, highlighting that several have IC50s in the nanomolar range. This includes 53 and 56, with IC50 values of 5.6 to 9.6 nM in HCT116 (colorectal carcinoma), NCI-H460 (lung carcinoma), and DLDI (adenocarcinoma) cells; 54, with IC50 values of 28 and 40 nM in HCT116 and QG56 (lung carcinoma) cells; 57 to 58, with IC50 values of 15 to 71 nM in HCT116 and QG56 cells; 60, with an IC50 value of 4.7nM in T47D (breast carcinoma) cells, and 53, with IC50 values of 3.1 and 6.0 nM in U373MG (brain carcinoma) and MCF-7 (breast carcinoma) cells.27,28 However, as their separation and extraction from marine organisms is complex, the attention of researchers has lately focused more on their total synthesis from commercially available materials, and on the development of semisynthetic analogues.29
Amino nitriles: renieramycins.
Renieramycin-type alkaloids have also been obtained from the nudibranch, Jorunna funebris (Mollusca: Gastropoda:Opisthobranchia: Nudibranchia: Kentrodorididae). It has been suggested the nudibranch sequesters renieramycins ingested within its Xestospongia prey sponges.30 In addition to the renieramycin-type alkaloids, related nitriles were isolated from KCN-treated homogenates of Thai Jorunna funebris.31 These compounds, jorunnamycins A-C, 66 to 68, are stabilized renieramycin-type bistetrahydroisoquinolines (Figure 7). Jorunnamycins, notably 68, showed cytotoxic activity against a range of cancer cell lines, including colon (HCT116 IC50 1.5 nM), lung (QG56 IC50 2.8 nM), and prostate (DU145 IC50 0.32 nM) compared to those of ecteinascidin 770 (0.40, 1.8 and, 0.66 nM respectively).
Amino nitriles: jorunnamycins, ecteinascidins and others.
Further nitrile-substituted isoquinoline derivatives are ecteinascidins 770 and 786 (69-70), 2 members of an interesting class of compounds derived from sea squirts, which show cytotoxic activity (Figure 7). Both compounds were isolated from KCN-treated homogenates of the Thai tunicate Ecteinascidia thurstoni Herdman 1891. The KCN pre-treatment described in the isolation of these types of compound was developed to protect the labile α-hydroxyamine functionality of the natural products.32 The importance of the C-21 substituent is reinforced by the suggestion that elimination of the cyano (or hydroxyl) group at this position under physiological conditions results in a reactive iminium species that is responsible for covalent bond formation with the drug target. The related compound, ecteinascidin-743, or trabectedin, which became the first marine drug to be licensed to treat soft tissue sarcoma, has a C-21 hydroxyl functionality. It was subsequently licensed for relapsed ovarian cancer in combination with doxorubicin. Ecteinascidins work as selective transcription inhibitors, uniquely poisoning transcription-coupled nucleotide excision repair. The structural complexity and rarity of the natural ecteinascidins have led to the development of a semisynthetic process from another nitrile, the readily available cyanosafracin B.26 The simple 1,2,3,4-tetrahydroisoquinoline 71 (Figure 7) was isolated from a low polar fraction of the crude extract of the KCN-pretreated Thai tunicate, Ecteinascidia thurstoni.33
Examination of a Chinese strain of Streptomyces, sp. HS-NF-1006, led to the isolation of the anthracycline analog 72, designated 6’’-cyano-6’’-deoxy-TAN-1120 (Figure 7).34 Evaluation of the cytotoxicity of 72 was explored using 2 human tumor cell lines, human hepatocellular liver carcinoma (HepG2) cells and human lung tumor cells (A549). Compound 72 had IC50 values of 0.06 and 0.04 μM against these strains, respectively, compared to the reference compound doxorubicin, whose corresponding IC50 values were 1.49 and 0.40 μM.
Bezerramycin C 73, a phenoxazinone antibiotic with a beta amino (and amido) substitution pattern (Figure 7), was isolated from Streptomyces griseus (HKI 0545, DSM 41823) growing on the plaster of an old building in Herne, Germany. The compound showed anti-proliferative activity against human vascular endothelium cells (HUVEC), with a GI50 value of 14.5 μM, but was inactive against K-562 (human chronic myeloid cells) and HeLa cells.35
Auranthine 74, a benzodiazepinone with a γ-amido nitrogen (Figure 7), was isolated from the fungal pathogen Penicillium aurantiogriseum (CBS 112021). Although originally reported in 1986, its structure was recently refined, revealing the presence of a previously unrecognized nitrile moiety.36 Following this structural refinement, its biological activity was further assessed. Cytotoxicity was evaluated using human liver cancer cells (HepG2) and human kidney epithelial cells (IHKE), using T-2 toxin (10 μM) as positive control. Cytotoxicity was established to be low, with no statistical significance observed compared to the solvent-treated control, except at the highest concentration (100 μM) tested on IHKE cells.
Phenylnitriles
Organisms in the phylum Arthropoda produce a number of simple nitrile-containing molecules, typically produced by the arthropods as components of viscous proteinaceous defensive secretions. Various millipedes produce HCN, alongside benzoyl nitrile 75, mandelonitrile 76, and mandelonitrile benzoate 77 (Figure 8).37 The centipedes Strigamia bothriopus and Geophilus vittatus also produce 75, with 76 additionally found in the latter species.38 The sternal gland secretions of Himantarium gabrielis L. (Himantariidae), a geophilomorph centipede, revealed the presence of 75 to 77, and also benzyl nitrile 78. This isolation was the first report on the presence of 77 and 78 in secreted substances from centipedes.38 All 4 compounds are considered as HCN precursors.
Simple phenylnitriles.
Benzyl nitrile 78 has also been identified as a constituent of the volatile oil of Salvadora persica L. (Salvadoraceae),39 and in the acetone extract of root bark.40 The stem oil was shown to exhibit antibacterial effects (expressed as zones of inhibition) on both sensitive and resistant strains of Pseudomonas aeruginosa (Schroeter and Migula) and S aureus (Rosenbach). The oil also showed significant inhibition against C albicans (C. P. Robin) and Trichosporon cutaneum (Beurm, Govgerot, and Vaucher), with comparable results to the reference drugs erythromycin, nystatin, tetracycline, and chloramphenicol.
Highly reactive α-ketonitrile compounds had not been reported in any plant-derived natural product; however, in a study of Arabidopsis thaliana genes upregulated in response to pathogens, the unprecedented cyanogenic metabolite 4-hydroxy-1H-indole-3-carbonyl cyanide (4-OH-ICN; 79) was isolated (Figure 8).41
Cruciferous vegetables, including various Brassica spp., are known for their various benefits and seem active in preventing carcinogenesis. In a review of chemopreventive activities of volatile organic compounds of common vegetables, nitriles 80 (3-phenylpropanenitrile) and 81 (2-(1H-indol-3-yl-acetonitrile) (Figure 8) isolated from broccoli (Brassica oleracea L., Italica group), were highlighted for their cardioprotective and neuroprotective applications.42
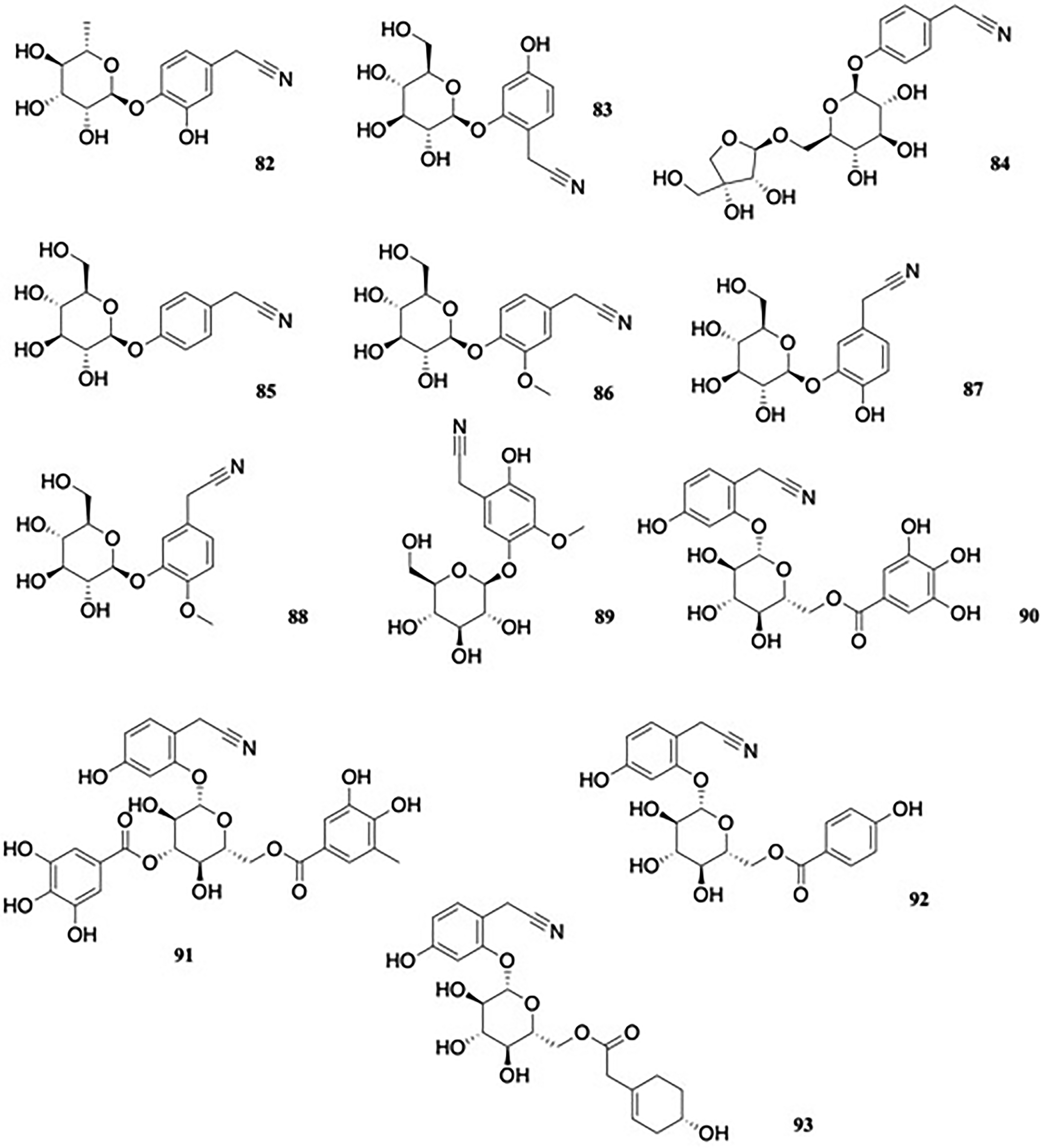
The new nitrile glycoside, 4-[(6-deoxy-α-L-mannopyranosyl)oxy]-3-hydroxybenzeneacetonitrile, named as niaziridin, 82 (Figure 9), alongside the previously known niazirin, were obtained from the leaves, pods, and bark of Moringa oleifera Lam. (Moringaceae).43 Niaziridin was described as a bioenhancer for drugs and nutrients, having synergistic properties alongside antibiotics and aiding gastrointestinal absorption of drugs and nutrients.
Phenylnitrile glycosides.
In a study of the activity of niazirin and niaziridin against clinical isolates of multidrug-resistant E coli, it was observed that the glycosides did not possess antibacterial activity of their own, but when used in combination, they reduced the MIC of tetracycline from 8 to 16-fold.44 Niaziridin (82) inhibited efflux pumps, downregulated expression of the efflux pump genes acrB and yojI, and over-expressed the porin forming genes ompA and ompX. It also decreased the mutation prevention concentration of tetracycline.44
Investigation of the ethanol extract of the roots of Semiaquilegia adoxoides DC. Makino (Ranunculaceae), a plant of traditional Chinese medicine, led to the isolation of the novel glycoside 2-(β-D-glucopyranosyloxy)-4-hydroxybenzeneacetonitrile 83 (Figure 9), although its biological activity was not reported.45 In a later study of nitrile and nitro-containing compounds from the roots of S. adoxoides, the researchers isolated various nitriles, including 4-[β-D-apiofuranosyl-(1→6)-O-β-D-glucopyranosyl oxy]phenylacetonitrile (84) (Figure 9).46
In addition to cyanogens (vide infra), the ornamental plant Hydrangea macrophylla (Thunb.) Ser. (Saxifragaceae) yielded four non-cyanogenic cyanoglucosides; 3-(β-D-glucopyranosyloxy)-4-hydroxybenzeneacetonitrile 85, named hydranitriloside A1; 3-(β-D-glucopyranosyloxy)-4-methoxybenzeneacetonitrile 86, named hydranitriloside A2; 4-(β-D-glucopyranosyloxy)-3-methoxybenzeneacetonitrile 87, named hydranitriloside B1; and 4-(β-D-glucopyranosyloxy)benzeneacetonitrile 88, named hydranitriloside B2 (Figure 9).47 Biological activities were not reported. Further constituents of H. macrophylla include 5-(β-D-glucopyranosyloxy)-2-hydroxy-4-methoxybenzeneacetonitrile 89, termed hydracyanoside E (Figure 9).48
Four esters; benzeneacetonitrile, 4-hydroxy-2-[[6-O-(3,4,5-trihydroxybenzoyl)-β-D-glucopyranosyl]oxy] 90; benzeneacetonitrile, 2-[[3,6-bis-O-(3,4,5-trihydroxybenzoyl)-β-D-glucopyranosyl]oxy]-4-hydroxy 91; benzeneacetonitrile, 4-hydroxy-2-[[6-O-(4-hydroxybenzoyl)-β-D-glucopyranosyl]oxy] 92, and benzeneacetonitrile, 4-hydroxy-2-[[6-O-[2-[(4S)-4-hydroxy-1-cyclohexen-1-yl]acetyl]-β-D-glucopyranosyl]oxy] 93, of a nitrile-containing phenolic glucoside were isolated from the leaves of Glochidion acuminatum Müll. Arg. (Euphorbiaceae)49 (Figure 9). The degalloylated analogue of 90 had been previously isolated from Ehretia philippinensis A. DC. (Boraginaceae), as ehretioside B.50 Biological data were not reported.
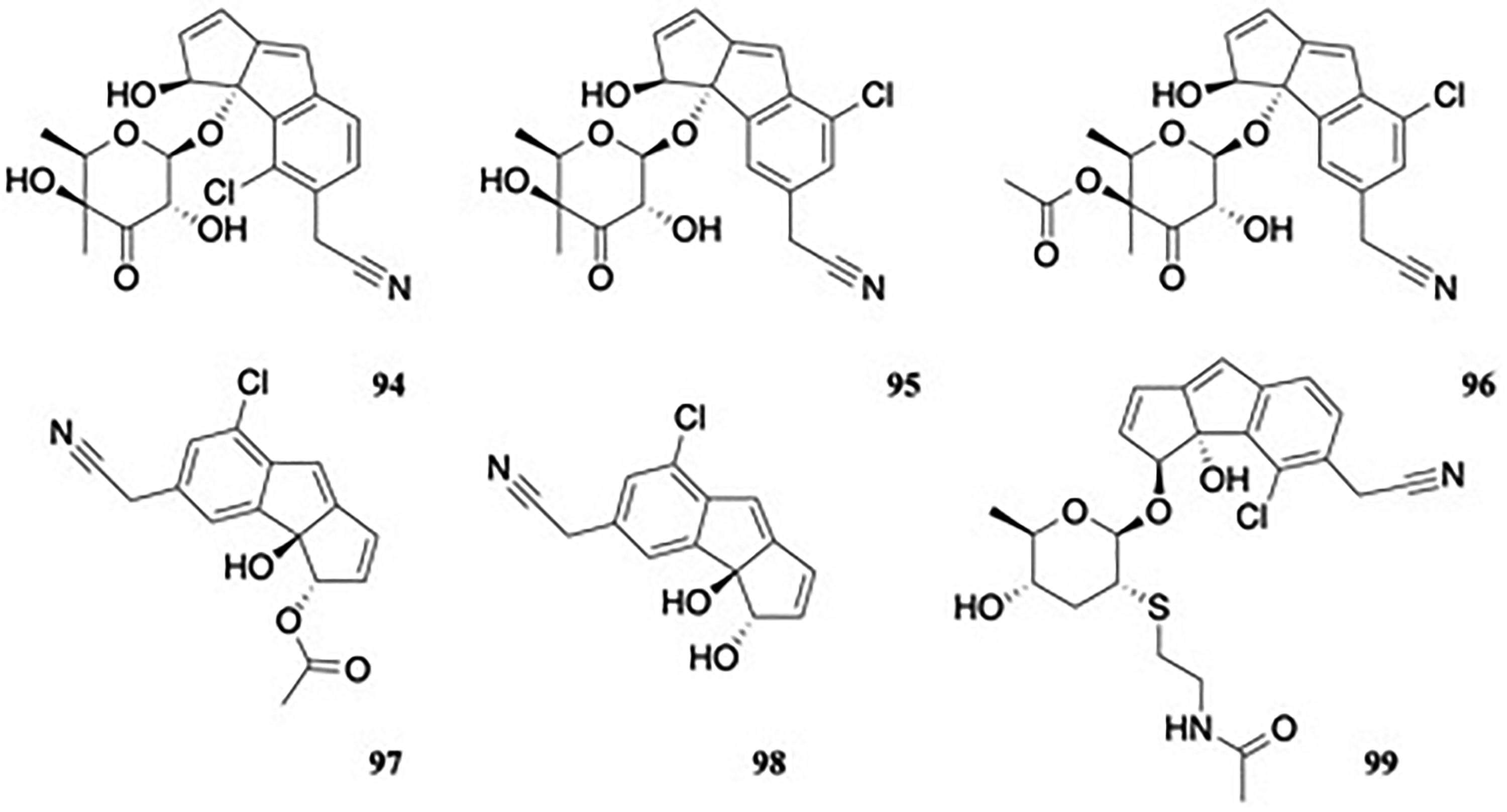
The Actinobacteria is mainly constituted of Gram-positive bacteria. Within this phylum, Salinispora pacifica CNS103, an obligate marine actinomycetes bacterial species, produces cyanosporasides A-F (94-99).51,52 These natural products contain a chlorinated cyclopenta[a]indene core (Figure 10). Bioactivity data were not reported for these isolates.
Cyanosporasides A-F.
Cyanogenic Glycosides
Cyanogenic glycosides represent the most common naturally occurring nitriles. These compounds are defined by the presence of an α-nitrile group adjacent to a glycosidic bond. These compounds are biosynthesized from a small number of amino acids, typically valine, leucine, isoleucine, phenylalanine, and tyrosine, or more rarely, 2-(2’-cyclopentenyl)-glycine or 2-(2’-hydroxy-3’-cyclopentenyl)-glycine.13 A typical biosynthetic sequence involves hydroxylation at the amino group, followed by elimination of carbon dioxide to afford an aldoxime and ultimately dehydration to yield the nitrile (Scheme 1). Further elaboration such as alpha-hydroxylation and β-glycosylation yields the glycosides. Hydrolysis of these compounds by glycosidases, followed by oxidation of the resultant cyanohydrin, releases the corresponding carbonyl compound (aldehyde/ketone), and hydrocyanic (prussic) acid. Many important food species (eg, cassava, lima beans, and almonds) contain cyanogens,9 which can only be consumed following treatment of the source plant material to destroy the cyanide content, typically by heat treatment.
Scheme 1. Biosynthesis of cyanogenic glycosides
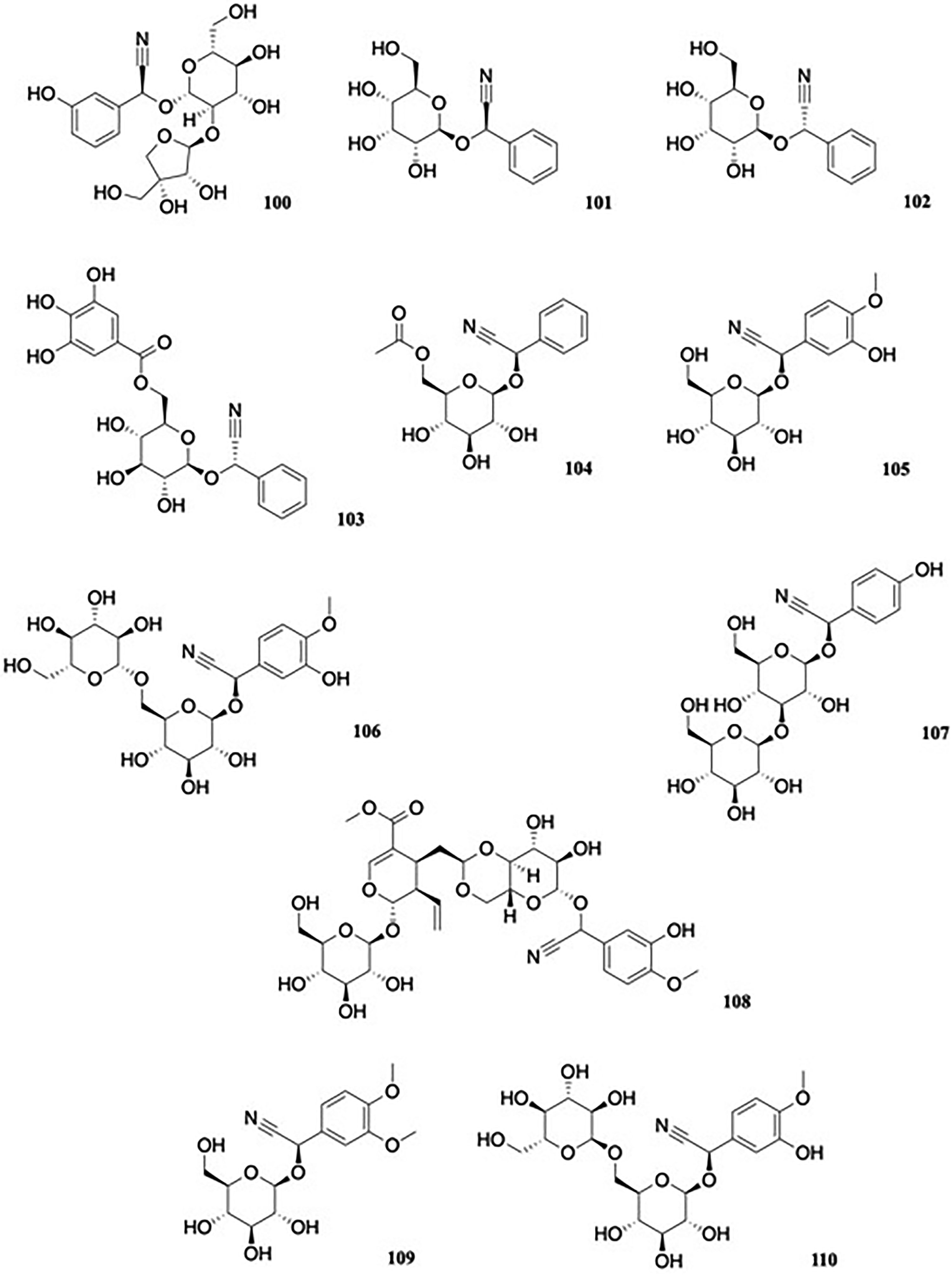
Alongside already known constituents, a new cyanogenic glycoside, 2S-β-D-apio-D-furanosyl-(1→2)-β-D-glucopyranosylmandelonitrile, 100, was isolated from the leaves of Sambucus nigra L. (Adoxaceae) (Figure 11).53 Biological data were not reported.
Cyanogenic glycosides.
Leaf and stem material of the passion fruit, Passiflora edulis Sims (Passifloraceae) were found to contain new cyanogenic glycosides (2R)-β-D-allopyranosyloxy-2-phenylacetonitrile 101 and (2S)-β-D-allopyranosyloxy-2-phenylacetonitrile 102 (Figure 11), alongside known compounds.54 The galloylated cyanogen (αS)-α-[[6-O-(34,5-trihydroxybenzoyl)-β-D-glucopyranosyl]oxy]benzeneacetonitrile, named 6′-O-galloylsambunigrin, 103 (Figure 11) was isolated from the foliage of the Australian tropical rainforest tree species Elaeocarpus sericopetalus F. Muell. (Elaeocarpaceae).55 Biological activity was not reported. Exploration of the ethyl acetate fraction of the ethanolic extract of the traditional Chinese medicinal plant Dracocephalum peregrinum L. (Lamiaceae) afforded a novel compound, 104 (Figure 11), (2R)-β-D-(6-O-acetyl)-glucosyl-2-phenylacetonitrile, named as peregrinumcin A, a 6’-O-acetyl derivative of the known cyanogen 2R-prunasin.56 The compound was tested for its effects on nitric oxide (NO) and nuclear factor (NF)-kB activity on RAW 264.7 and pNF-kBluc-293 cells, and showed 49% inhibitory activity of NO production induced by LPS at a dose of 100 μg/ml compared to the standard aminoguanidine (75% inhibition at 50 mM), but lacked significant effects on NF-kB activity at the same dose (18% inhibition compared to 75% inhibition by the standard LGT at 10 μg/mL). Reports of toxicity following ingestion of the leaves of the ornamental plant Hydrangea macrophylla (Thunb.) Ser. (Saxifragaceae) led to the isolation of three new cyanogenic glycosides: (αR)-α-(β-D-glucopyranosyloxy)-3-hydroxy-4-methoxybenzeneacetonitrile, named hydracyanoside A 105; (αR)-α-[(6-O-β-D-glucopyranosyl-β-D-glucopyranosyl)oxy]-3-hydroxy-4-methoxybenzeneacetonitrile, named hydracyanoside B 106, and (αR)-α-[(3-O-β-D-glucopyranosyl-β-D-glucopyranosyl)oxy]-3-hydroxy-4-methoxybenzeneacetonitrile, named hydracyanoside C 107 (Figure 11) from the leaves and/or stems, along with the known cyanogen, taxiphyllin.57
(αR)-α-[[4,6-O-[(1R)-2-[(2S,3R,4S)-3-Ethenyl-2-(β-D-glucopyranosyloxy)-3,4-dihydro-5-(methoxycarbonyl)-2H-pyran-4-yl]ethylidene]-β-D-glucopyranosyl]oxy]-3-hydroxy-4-methoxybenzeneacetonitrile, named as hydracyanoside D, 108 (Figure 11), was later isolated from the plant.48 Most recently, two additional cyanogenic glycosides, [(2R)-2-(β-D-glucopyranosyloxy)-2-(3,4-dimethoxy-phenyl)] acetonitrile 109 and {(2R)-2-[α-D-glucopyranosyl(1→6)β-D-glucopyranosyloxy]-2-(3-hydroxy-4-methoxy-phenyl)}acetonitrile 110 (Figure 11), were isolated from the aerial parts of H. macrophylla.58
Hedyotis scandens Roxb. (Rubiaceae) is used in traditional Chinese medicine for the treatment of respiratory diseases. Compound 111, benzeneacetonitrile 6′-O-(5″-O-p-coumaroyl)-β-D-apiofuranosyl-β-D-glucopyranoside, termed hedyotoside A (Figure 12), was isolated from the ethyl acetate fraction of the ethanol extract of the whole plant.59 Hedyotoside A was subject to evaluation for antiviral properties using the cytopathic effect reduction assay but showed no inhibitory activity against RSV at its maximal nontoxic concentration, compared to the control drug ribavirin (IC50 value of 1.5 μg/mL).
Cyanogenic glycosides.
A new cyanogenic glucoside, benzeneacetonitrile, α-[[6-O-(2-methyl-1-oxo-2-propen-1-yl)-β-D-glucopyranosyl]oxy]-, (αR)-, named 6’-methacrylate prunasin 112 (Figure 12), was isolated from aerial parts of Centaurea microcarpa Coss. & Dur. (Asteraceae).60
The cyanogenic glycoside 2-(β-D-glucopyranosyloxy)-2-(hydroxymethyl)butanenitrile, named as sachaloside V, 113 (Figure 12), alongside known cyanogens rhodiocyanoside A, lotaustralin, and heterodendrin, was isolated from the methanolic extract of the roots of Rhodiola sachalensis A. Bor. (Crassulaceae).61 The effects of the extract and its principal constituents were evaluated on D-Galactosamine-induced cytotoxicity in primary cultured mouse hepatocytes, with both the crude extract (98.8% inhibition at 100 μg/mL) and some isolated constituents showing promising activity. However, those constituents (including 113) occurring in smaller quantities (such as the 0.0007% reported for 113) were not individually assessed.
Linum grandiflorum Desf. (Linaceae) has been used as a medicinal herb for various ailments, and has shown cytotoxicity against the EL4 cell line. In addition to its previously known nitrile content in the form of various cyanogenic glycosides, other research established the presence of 2-[(3’-isopropoxy-O-β-D-glucopyranosyl)oxy]-2-methylbutanenitrile 114 in the leaves of the plant (Figure 12).62 When evaluated for cytotoxic activity, 114 had an IC50 value of 0.3 μM against the EL4 cell line, compared to the positive control, the cytotoxin thapsigargin, which had an IC50 value of 1.9 μM.63
The first report of a cyanogenic glycoside in mistletoes was that of 2-methylpropionitrile-2-O-(6-O-galloyl)-β-D-pyranoglucoside, named as linamarin gallate 115 (Figure 12), isolated from the leafy twigs of the Nigerian species Loranthus micranthus L. (Loranthaceae), parasitic on Hevea brasiliensis Müll.Arg. (Euphorbiaceae).64 The compound was evaluated for antioxidant activity and had an IC50 value of 39 μM, being more active than the reference compound chlorogenic acid (IC50 value of 67.9 μM) in the DPPH assay.
The aqueous acetone extract of the Chinese medicinal species, Balanophora involucrata Hook. f. & Thomson (Balanophoraceae), a parasitic plant that grows on the roots of leguminous plants, showed radical scavenging activity. Phytochemical profiling revealed a novel cyanogenic glycoside, proacacipetalin 6’-O-β-D-glucopyranoside 116 (Figure 12).65
Linustatins A ((2-[(O-β-D-glucopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranosyl)oxy]-2-methyl-(2R)-butanenitrile) 117, B ((2-[(O-β-D-glucopyranosyl-(1→6)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranosyl)oxy]-2-methyl-(2R)-butanenitrile) 118, and C ((2-[(O-β-D–glucopyranosyl-(1→6)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranosyl)oxy]-2-methyl-propionitrile) 119, new cyanogenetic triglycosides (Figure 12), were isolated from linseed meal (L. usitatissimum L. (Linaceae)).66 Compounds 117 to 119 had moderate activity against aldose reductase (30.0%, 35.5%, and 30.0% at 1 × 10-5M, respectively, compared to 99.4% for the positive control, epalrestat) and weak activities against α-glucosidase, DPP-IV, and FBPase.
Nitrilosides
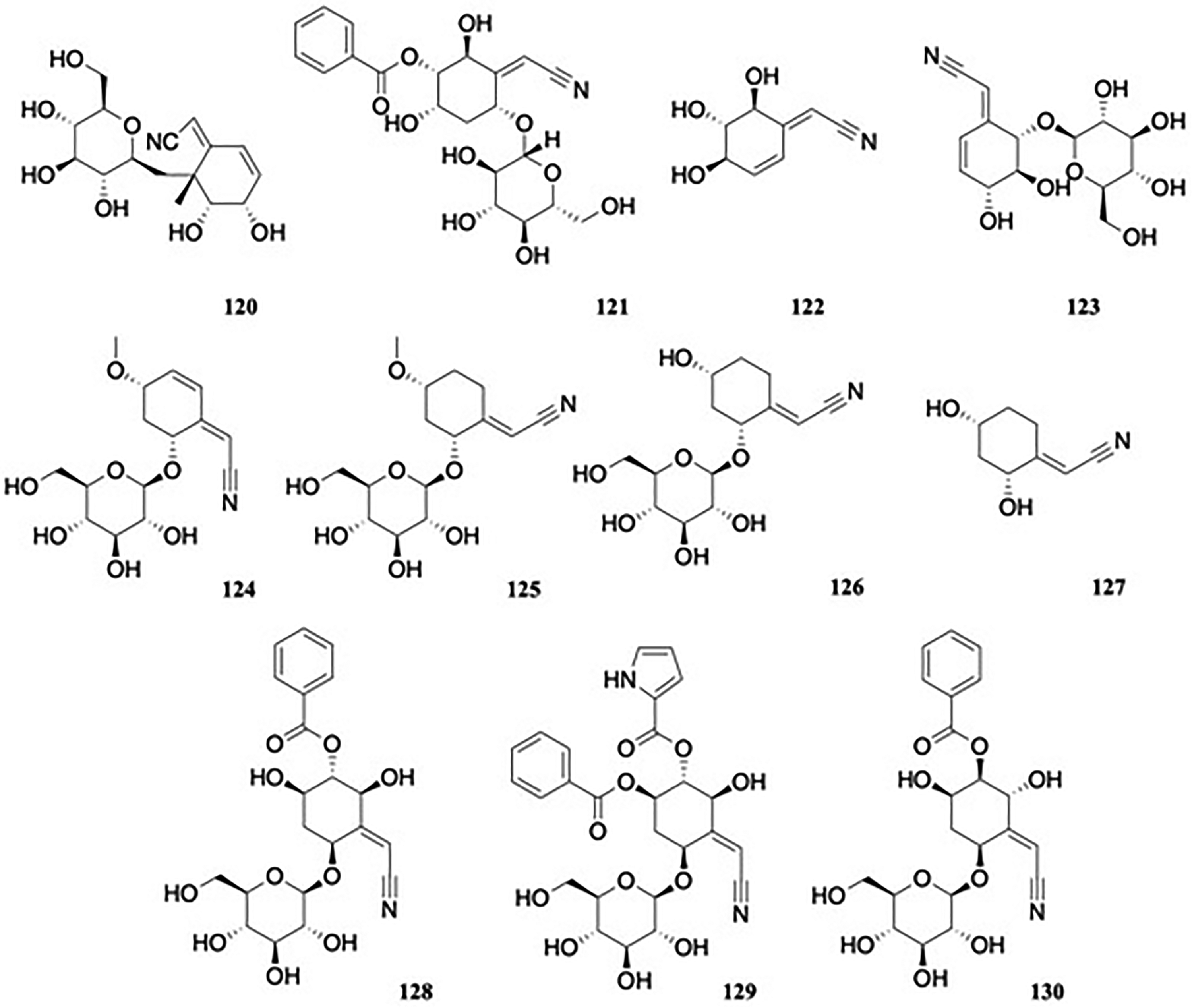
A smaller group of cyanoglycosides are characterized by the position of the cyano group not next to the glycosidic bond of the molecule, so their hydrolysis does not result in the release of cyanide, thus decreasing their toxicity. One such compound is (2Z)-8-(β-D-glucopyranosyloxy)-6,7-dihydroxycyclohex-4-en-3-ylidene]-ethane-nitrile, named as riachin 120 (Figure 13), obtained from the root bark of Bauhinia pentandra (Bong.) Vog. Ex Steud (Fabaceae).67 Riachin was tested for antimicrobial activity against various bacterial strains (E coli ATCC 25922, S aureus ATCC 25923, and P aeruginosa ATCC 25853), using gentamicin, amikacin, and clindamycin as control antibiotics. The MIC in all cases was ≥1024 μg/mL, with the exception of the sample against E coli 25922, which had a MIC value of 128 μg/mL, thus showing the compound had no antimicrobial activity against the tested organisms.68 However, it did show synergism when combined with amikacin against P aeruginosa and combined with clindamycin against S aureus, an effect suggested to be due to altered pharmacokinetics.
Nitrilosides.
(2Z)-[(2S,3S,4S,6R)-3-(Benzoyloxy)-6-(β-D-glucopyranosyloxy)-2,4-dihydroxycyclohexylidene]acetonitrile, named as lanceolin C 121 (Figure 13), is another nitrile glucoside, isolated from the stem heartwood of Lophira alata Banks ex C.F.Gaertn. (Ochnaceae).69 It was subsequently isolated from the related L. lanceolata Van Tiegh. Ex Keay, and its antitubercular activity were evaluated against M. tuberculosis AC45 and AC83 strains; however, the compound was not considered active, with a MIC value of 62.5 μg/mL against AC45 and a CMI >250 µg/mL against strain AC83, compared to the control drug isoniazid (MIC 3.9 µg/mL).70 In the aforementioned study of nitrile and nitro-containing compounds from the roots of Semiaquilegia adoxoides,46 another novel nitrile isolated was the cyclohexenylidene derivative 122, (1E,4α,5β,6α)-45,6-trihydroxy-2-cyclohexen-1-ylideneacetonitrile (Figure 13).
The leaves of Bauhinia holophylla (Bong.) Steud. (Fabaceae), a plant used in Brazilian traditional medicine to treat diabetes71 afforded the new compound (-)-(1E,4R,5S,6S)-6-(β-D-glucopyranosyloxy)-4,5-dihydroxy-2-cyclohexen-1-ylideneacetonitrile 123 (Figure 13), a stereoisomer of the known compound (-) lithospermoside, itself also isolated, along with its steric isomer aglycone at C-1, ( + )-(1E,4R,5S,6S)-45,6-trihydroxy-2-cyclohexen-1-ylideneacetonitrile.
In addition to known nitriles, four new cyclohexylideneacetonitrile derivatives; acetonitrile, 2-[(4R,6S)-6-(β-D-glucopyranosyloxy)-4-methoxy-2-cyclohexen-1-ylidene]-, (2Z) 124, named menisdaurin B; acetonitrile, 2-[(2S,4S)-2-(β-D-glucopyranosyloxy)-4-methoxycyclohexylidene]-, (2Z) 125, named menisdaurin C; acetonitrile, 2-[(2S,4S)-2-(β-D-glucopyranosyloxy)-4-hydroxycyclohexylidene]-, (2Z) 126, named menisdaurin D, and acetonitrile, 2-[(2S,4S)-2,4-dihydroxycyclohexylidene], (2Z) 127, named menisdaurin E (Figure 13) were isolated from the hypocotyl of the mangrove Bruguiera gymnorrhiza (L.) Savigny (Rhizophoraceae).72 The compounds showed anti-hepatitis B virus (HBV) activities, with EC50 values ranging from 8.7 to 30.7 μg/mL, compared with the reference drug, lamivudine (EC50 value of 0.1 μg/mL).
Campyloside A, (1S,3S,4S,5R)-4-benzoyloxy-2-cyanomethylene-3,5-dihydroxycyclohexyl-1-O-β-glucopyranoside 128 and campyloside B, (1S,3S,4S,5R)-5-benzoyloxy-2-cyanomethylene-3-hydroxy-4-(2-pyrrolcarboxyloxy)cyclohexyl-1-O-β-glucopyranoside 129, were extracted from the roots of Campylospermum glaucum (Tiegh) Farron (Ochnaceae),73 and Campyloside C 130 from Campylospermum excavatum (Tiegh) Farron (Figure 13).74 Extracts of C. glaucum showed good antimicrobial activity against Gram-positive cocci (Enterococcus hirae ATCC 9790, Enterococcus sp. P054, S aureus ATCC 25923, S aureus U271 and S. saprophyticus), with MIC values of 5 mg/mL, 2.5 mg/mL, 2.5 mg/mL, 2.5 mg/mL and 1.25 mg/mL, respectively, compared to the standard gentamicin, with MIC values of 16 μg/mL, 2 μg/mL, 16 μg/mL, <0.125 μg/mL and <0.125 μg/mL. However, no activity was seen against either Gram-negative bacilli or fungi, and the isolated campylosides A and B were inactive on the same bacterial strains.73
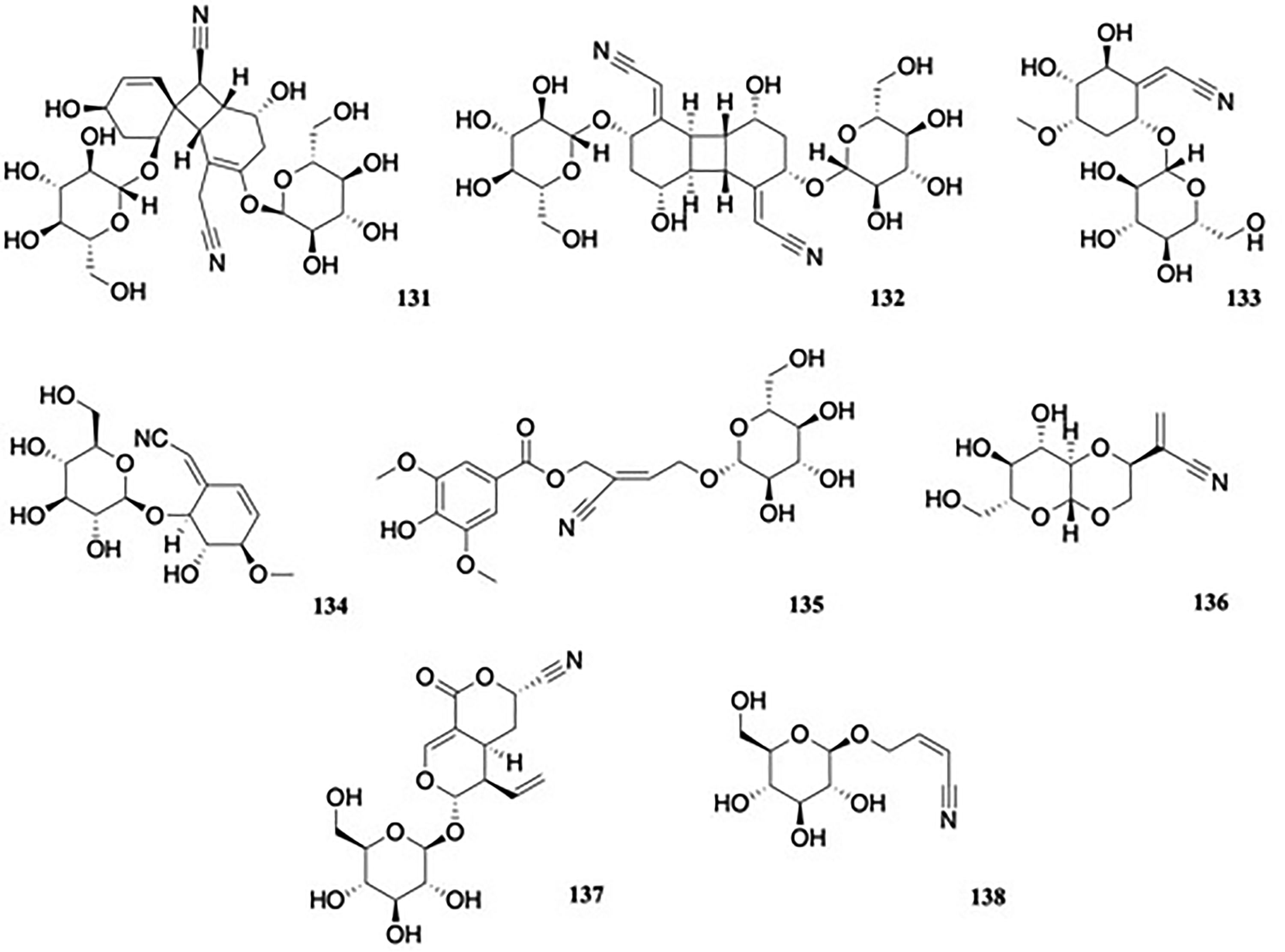
Simmondsia chinensis (Link) C. Schneider (Simmondsiaceae) is a tree of desert and semidesert areas of the southwestern United States and northwestern Mexico, known commonly as jojoba, and has important ethnopharmacological and commercial applications. Examination of the aqueous leaf extract revealed (1S,4'S,5R,6R,6’R,7S,8S)-2-(cyanomethyl)-5,6’-dihydroxy-3,4’-bis[[(2R,3R,4S,5S,6R)-34,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy]spiro[bicyclo[4.2.0]oct-2-ene-8,3’-cyclohexene]-7-carbonitrile 131, named as simmonoside A and (2Z)-2-[(2S,4R,5Z,6S,8R)-5-(cyanomethylidene)-4,8-dihydroxy-2,6-bis[[(2S,3R,4S,5S,6R)-34,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy]-2,3,4,4a,4b,6,7,8,8a,8b-decahydrobiphenylen-1-ylidene]acetonitrile 132, named as simmonoside B,75 alongside four known cyanogenic glycosides, menisdaurin, simmondsin, 4-demethyl simmondsin 133 (itself first reported as a pure isolate in 200076) and 5-demethyl simmondsin (Figure 14). Compounds 131 and 132 represent the first noncyanogenic cyanoglucoside dimers possessing a unique four-membered ring, representing novel dimerization patterns at α,β-unsaturated carbons of a nitrile group in 131 and γ,δ-unsaturated carbons in 132. When screened for cytotoxicity and COX-2 inhibitory effects, 131 and 132 exhibited moderate cytotoxicity against A-549 human lung adenocarcinoma (IC50 values of 21.5 and 17.7 μM) and SGC-7901 human gastric cancer (IC50 values of 26.8 and 20.4 μM), compared to the standard cisplatin (IC50 values of 11.6 and 7.5 μM), and COX-2 inhibition activities (IC50 values of 13.5 and 11.4 μM, compared to 1.6 μM for the positive control indomethacin).
Nitrilosides.
The 6-glycoside of the nitrile-containing metabolite (Z)-5α,6β-dihydroxy-4β-methoxy-2-cyclohexene-Δ(1,α)-acetonitrile 134 (Figure 14), was isolated from an ethanol extract of the crushed canes of Bauhinia aurea H.Lév. (Fabaceae).77 Biological activities were not reported.
Another cyanoside, (2Z)-2-cyano-4-(β-D-glucopyranosyloxy)-2-buten-1-yl 4-hydroxy-3,5-dimethoxybenzoate 135, named as rhobupcyanoside A (Figure 14), was first isolated from the ethanol extract of the stems of Rhodiola bupleuroides.78 The compound was re-isolated later from R. wallichiana and its antitumor effects evaluated.79 The related compound rhobupcyanoside B, 136 was later isolated from R. bupleuroides and its effects on inhibition of α-glucosidase evaluated.80136 had an IC50 value of 278.28 ± 0.55 μM while the IC50 for the positive control, acarbose was 210.40 ± 0.32 μM.
(3S,4aS,5R,6S)-5-Ethenyl-6-(β-D-glucopyranosyloxy)-4,4a,5,6-tetrahydro-1-oxo-1H,3H-pyrano[3,4-c]pyran-3-carbonitrile 137, named as hydracyanoside F (Figure 14), was isolated among the constituents of H. macrophylla.48
Alliarinoside, or (Z)-4-(β-D-glucopyranosyloxy)but-2-enenitrile 138 (Figure 14), is a cyanoallyl glycoside isolated from Alliaria petiolata (M.Bieb.) Cavara & Grande (Brassicaceae), or garlic mustard, an invasive plant of North America.81 The compound is known as an allelochemical, acting as a feeding inhibitor against early instar larvae of Pieris napi oleracea.
Methylbutenenitriles
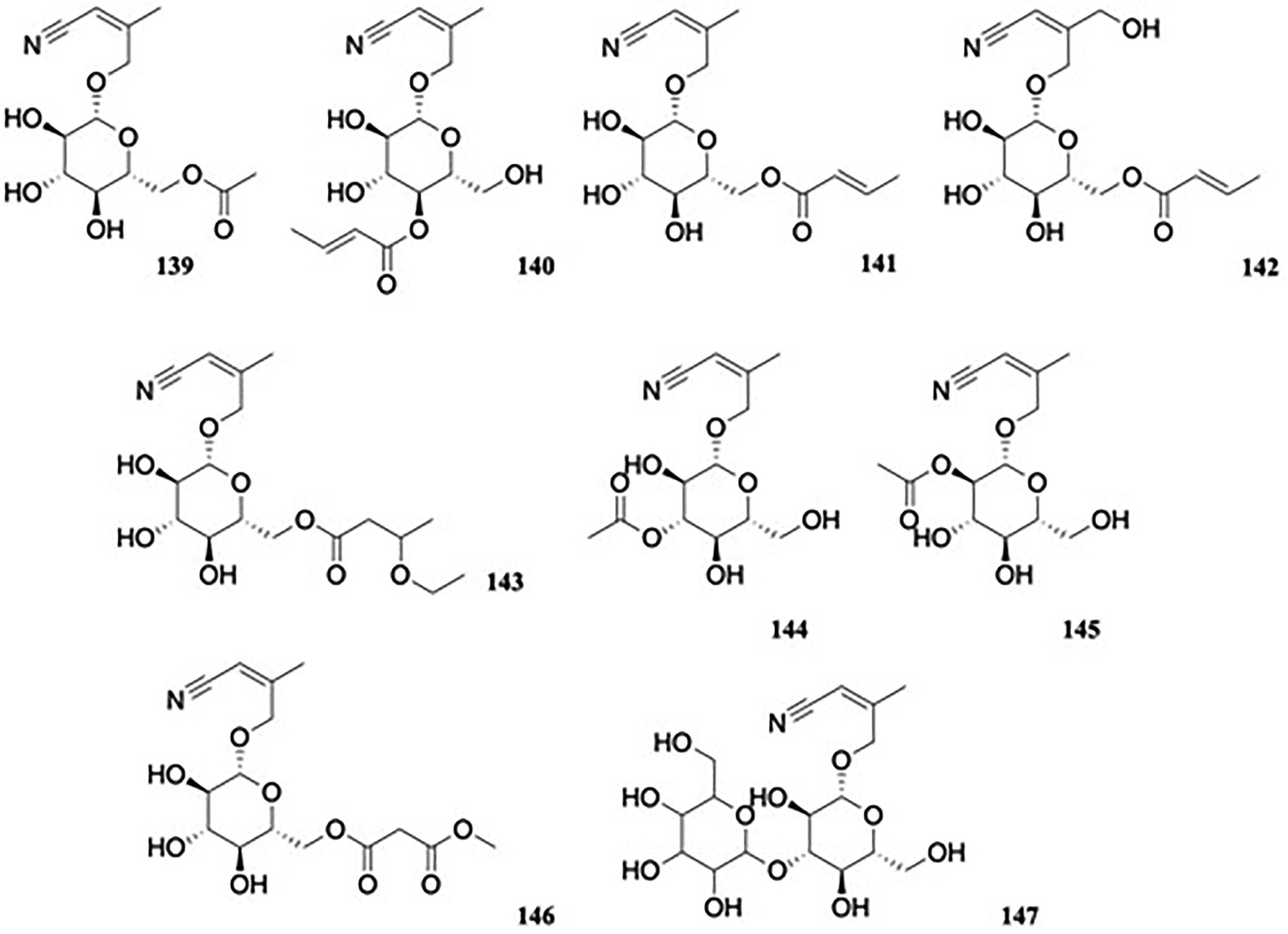
Five new γ-hydroxynitrile glucosides, [(6-acetate)-β-D-glucopyranosyloxy]-3-methyl-2Z-butenenitrile 139, 4-[4-(2E)-2-butenoate]-β-D-glucopyranosyloxy-3-methyl-2Z-butenenitrile 140, 4-[6-(2E)-2-butenoate]-β-D-glucopyranosyloxy-3-methyl-2Z-butenenitrile 141, 4-[6-(2E)-2-butenoate]-β-D-glucopyranosyloxy-3-hydroxymethyl-2Z-butenenitrile 142, and 4-[6-(3-ethyoxyl)-butyrate]-β-D-glucopyranosyloxy-3-methyl-2Z-butene-nitrile 143, named prinsepicyanosides A–E, (Figure 15) were isolated from the seeds of the Chinese medicinal plant Prinsepia utilis Royle (Rosaceae).82 Among these, prinsepicyanoside B 140 was the first reported γ-hydroxynitrile glucoside in which the hydroxyl group on C-4 of glucose was derivatized. When screened for antimicrobial activity, 139 inhibited Salmonella gallinarum with a MIC value of 30.1μg/mL, compared to the standard drug ciprofloxacin with a MIC value of 0.41 μg/mL.
Methylbutenenitriles.
Subsequently, 4 additional glycosides, prinsepicyanosides F-I, 144 to 147 (Figure 15) were obtained from the seed oil residue, with 146 exhibiting weak α-glucosidase inhibitory activity (15% inhibition (at 50 μM) compared to 61.2% and 63.2% inhibition using the standard drugs quercetin (at 10 μM) and acarbose (at 500 μM), respectively.83
Ribes species (Grossulariaceae), such as R. rubrum and R. nigrum, are usually cultivated for their edible berries, but their seeds contain interesting metabolites. 2-(4-hydroxybenzoyloxymethyl)-4-β-D-glucopyranosyloxy-2(E)-butenenitrile 148 and 2-(4-hydroxy-3-methoxybenzoyloxymethyl)-4-β-D-glucopyranosyloxy-2(E)-butenenitrile 149 were isolated from R. rubrum seeds,84 while 2-trans-p-coumaroyloxymethyl-4-β-D-glucopyranosyloxy-2(E)-butenenitrile 150, known as nigrumin-5-p-coumarate and 2-trans-feruloyloxymethyl-4-β-D-glucopyranosyloxy-2(E)-butenenitrile 151, known as nigrumin-5-ferulate were obtained from R. nigrum seeds85 (Figure 16). These structures are related to the previously isolated sutherlandin-5-trans-p-coumarate 152 found in Sorbaria sorbifolia (L.) A. Br. var. stellipila MAX. (Rosaceae).86 Astringency of nitriles 148 and 149 was evaluated, and showed that these compounds were not primarily responsible for the astringency of the plant, having threshold concentrations similar to those of most of the naturally occurring flavon-3-ol glycosides.84
Methylbutene- and methylbutanenitriles.
The γ-hydroxynitrile glycoside (E)-2-(hydroxymethyl)-4-[(2R,3R,4S,5S,6R)-34,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxybut-2-enenitrile 153, known as sarmentosin (Figure 16), previously known from Sedum sarmentosum Bunge (Crassulaceae),87 and also known to be sequestered by lepidopteran species in the genus Parnassius (Papilionidae),88 was detected for the first time from the genus Bryophyllum/Kalanchoe, being isolated from both B. pinnatum L. Pers. (Crassulaceae) and Kalanchoe laciniata.89
In a study of the chemical constituents of a traditional Tibetan medicine, Rhodiola kirilowii (Regel) Maxim, used as an adaptogen in China, an ethanol extract of the roots was evaluated.90 In addition to known compounds, a new nitrile, 4-(β-D-glucopyranosyloxy)-3-hydroxy-2-(hydroxymethyl)-butanenitrile 154 (Figure 16), was isolated.
Glucosides with an unusual (2S,3R)-2-ethyl-2,3-dihydroxybutyric acid as the acyl moiety at the 6-position of glucose were extracted from the branches of Microtropis japonica Hallier (Celastraceae), a tree present in Kanto and Kyushu, Japan, the Okinawa Islands, and Taiwan91 These were (2R,3S)-3-hydroxy-2-methylbutanenitrile-β-D-glucopyranoside 6-O-(2’'S,3’’R)-2’’-ethyl-2’’,3’’-dihydroxybutyrate 155, named as microtropin A, (2R,3R)-3-hydroxy-2-methylbutanenitrile-β-D-glucopyranoside 6’-O-(2’'S,3’’R)-2’’-ethyl-2’’,3’’-dihydroxybutyrate 156, named as microtropin B, (2S,3S)-3-hydroxy-2-methylbutanenitrile-β-D-glucopyranoside 6’-O-(2’'S,3’’R)-2’’-ethyl-2’’,3’’-dihydroxybutyrate 157, named as microtropin C and (E)-4-hydroxy-2-(hydroxymethyl)but-2-enenitrile 4-O-β-D-glucopyranoside 6’-O-(2’'S,3’’R)-2’’-ethyl-2’’,3’’-dihydroxybutyrate 158, named as microtropin D (Figure 16). The nitrile component was either alkane (155-157) or α,β-unsaturated (158) in nature. The biological activity of these compounds was not studied. Further work on the plant led to the isolation of the Z-isomer of 158, named microtropin Q (159).92
Cyanolipids
The cyanolipids (CLs) represent a further category of natural nitriles, and are derived through amino acid (leucine) metabolism.93 They occur alongside acylglycerols and triacylglicerides in seed oils of members of the Boraginaceae, the Hippocastanaceae, and the Sapindaceae. Four types of CL structures (160-163) may be defined (Figure 17) with fatty acids esterified to a mono or a dihydroxynitrile moiety. CLs of types I and IV are cyanogenic, releasing cyanohydrins which spontaneously decompose forming HCN, while CLs of types II and III are not cyanogenic.
Cyanolipid classes.
Within the 4 CL classes, the primary variation lies in the fatty acid chains. An overview of CL-containing plants in the Sapindaceae profiled from 1998 to 2021 is shown in Table 1.
Cyanolipids (CL) Isolated From Plant Seed Oils Profiled from 1998 to 2021.
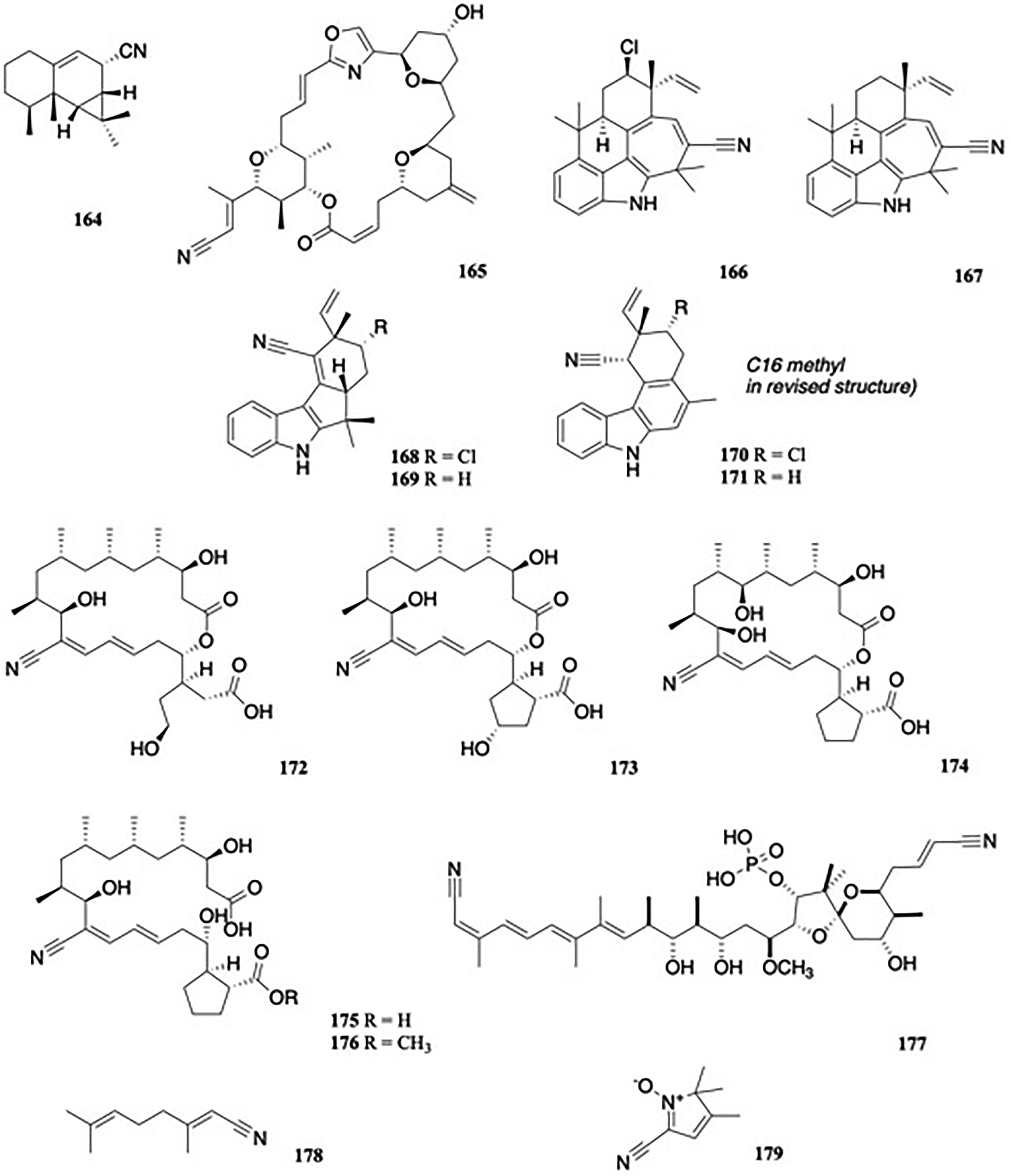
Among the metabolites of Axinyssa isabela (Halichondriidae), a marine sponge, was isolated the nitrile axinynitrile-A, 164.101 The presence of a nitrile in this compound represents a departure from the nitrogen-containing terpenoids so far described from sponges of the order Halichondrida, which usually contain isocyano, isothiocyanate, or formamide groups, and less frequently thiocyanate or isocyanate functionalities. When tested in cytotoxicity assays against the human cancer cell lines MDA-MB-231 (breast adenocarcinoma), A-549 (lung adenocarcinoma), and HT-29 (colon adenocarcinoma), 164 (Figure 18) was inactive at the highest concentration tested (10 μg/mL), compared to the positive control doxorubicin (GI50 = 0.1, 0.1, and 0.1 μM against MDA-MB-231, A-549, and HT-29, respectively.
α,β-unsaturated nitriles.
Hemi-phorboxazole A, 165 (Figure 18), an oxazole-containing macrolide with a terminal nitrile, was isolated from Phorbas sp..102 Although 165 displayed no activity when tested against C albicans and two human cancer cell lines, a semisynthetic analogue featuring B and C-ring modifications exhibited significant tumor cell growth inhibitory activity in the nanomolar range against HCT-116 (colon) and SK-BR-3 (breast) cells, while another derivative featuring solely C-ring modification displayed promising antifungal activity against C albicans.103
The phylum Cyanobacteria is composed of Gram-negative bacteria. The cyanobacterium Fischerella ambigua (Stigonemataceae) has been studied for its various secondary metabolites. Among these can be found ambiguines, hapalindoles, fischambiguines and 12-epi-hapalindoles.104 Although more typically characterized by an isonitrile or isothiocyanate functionality, some nitrile-containing metabolites are known to occur in this species,105 such as ambiguine G nitrile 166, first isolated from the blue-green alga Hapalosiphon delicatulus (UH isolate IC-13-1) (Stigonemataceae),106 and the novel deschloro analogue, ambiguine Q nitrile, 167 (Figure 18). Ambiguine G nitrile inhibited the growth of S aureus and M. tuberculosis, with MIC values of 6.6 and 53.7 μM, respectively, compared to the appropriate controls gentamicin (0.65 μM) and rifampin (0.10 μM), but had no effect on the growth of E coli.105
Four nitrile-containing fischerindole derivatives, namely 12-epi-fischerindole I nitrile 168, deschloro 12-epi-fischerindole I nitrile 169, 12-epi-fischerindole W nitrile 170, and deschloro-12-epi-fischerindole W nitrile 171 were separately isolated from Fischerella sp. (SAG strain number 46.79) (Figure 18), and evaluated for anticancer properties against HT-29 (colon), NCI-H460 (large lung), MCF-7 (breast), and SF268 (glioblastoma) lines,107 although the structure originally attributed to 171 was later revised.108 Compound 169 had an IC50 value of 23 μM against HT-29 cells, and was completely inactive against the other cell lines tested, while 170 and 171 were unstable under assay conditions. None of the compounds showed inhibition of 20S proteasome activity.
The borrelidins are a group of 18-membered polyketide macrolides produced by several species of Streptomyces, with the parent compound, borrelidin, known since 1949. Recent work on a saltern-derived halophilic Nocardiopsis species (strain HYJ128) from Korea led to the isolation of three new borrelidins, C-E, 172 to 174 (Figure 18), all possessing an α,β-unsaturated nitrile.109 Borrelidins C and D displayed antibacterial activity, particularly against the Gram-negative pathogen Salmonella enterica (MIC values of 16 and 63 μM, respectively), compared to the standard drug ampicillin (1.4 μM). Moderate cytotoxicity was demonstrated by 172 and 173 against the SNU638 (IC50 values of 5.5 and 8.7 μM) and K562 (IC50 values of 5.7 and 6.7 μM) carcinoma cell lines, compared to the standard drug etoposide (IC50 values of 0.57 and 1.5 mM).
Co-culture of Streptomyces rochei MB037 with the gorgonian-derived fungus Rhinocladiella similis 35 afforded two new fatty acids with a nitrile group, borrelidins J and K, 175 to 176 (Figure 18).110 The compounds exhibited interesting antibacterial activity against methicillin-resistant S aureus, with MIC values of 0.20 and 1.56 μg/mL respectively, compared to the reference drug ciprofloxacin (0.31 μg/mL).
The calyculins are a diverse group of cytotoxic polyketides. First isolated from the marine sponge Discodermia calyx in 1986, with related structures subsequently isolated from other sponges, now 18 such structures are known, with eleven of these bearing a nitrile functional group.111 More recently, a truncated form of calyculin A, named hemicalyculin A 177 (Figure 18), was isolated from D. calyx.112 When evaluated for its ability to inhibit protein phosphatase, a known target of the calyculins, the IC50 values of 177 were 14 nM and 1 nM against PP1γ and PP2A, respectively, compared with the parent structure calyculin A (IC50 values of 8.2 nM and 1 nM). In a comparison of enzyme inhibitory and cytotoxic activities, 177 proved much less active than calyculin A against P388 cells (IC50 values of 450 ng/mL vs 0.170 ng/mL). The absence of key basic functional groups on 177 was proposed to account for its reduced membrane permeability.
Heracleum is an umbelliferous plant genus widely distributed in Asia. Of the 10 species known in Iran, some are used for food and medicinal purposes. In a study of 2 Heracleum species, the essential oils obtained from the aerial parts of the plants were evaluated for antimicrobial activity.113 Geranyl nitrile 178 comprised 3.89% of H. transcaucasium essential oil, while the oil of the related species H. ansiactis indicated the presence of 5-cyano-22,3-trimethyl-2H-pyrrole 1-oxide 179 (4.83%) (Figure 18). The oils were screened against a panel of microorganisms (E coli ATCC (8739), P aeruginosa ATCC (9027), Staphylococcus epidermidis ATCC (12228), and S aureus ATCC (6538)); however, the essential oils of both species were inactive against the tested microbial strains when compared with the standard, amikacin.
Aromatic Nitriles
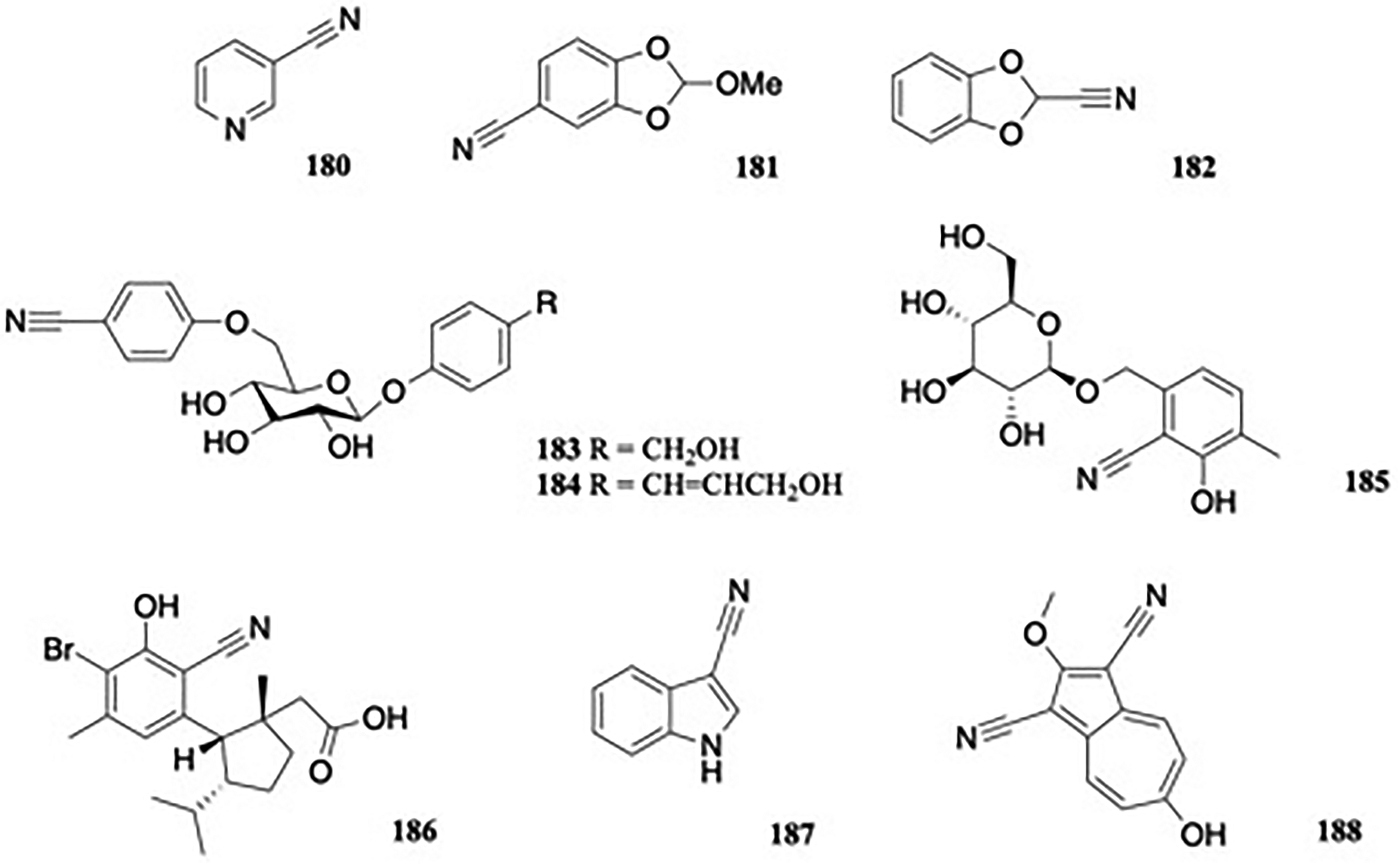
The unusual natural product, pyridine-3-carbonitrile 180 (Figure 19) was identified as a chemotaxonomic marker in Mercurialis annua L. (Euphorbiaceae), allowing differentiation from the closely related M. perennis.114
Aromatic nitriles.
Ryecarbonitrilines A and B (181-182) (Figure 19) were isolated from rye (Secale cereale L. (Poaceae)) root exudate.115 The compounds were assayed for their potential allelopathic effect on seed germination and radicle growth. Ryecarbonitriline A, 181 significantly induced Orobanche cumana seed germination at the concentration range tested (0.06-0.60 mM), but did not induce the germination of seeds of O. crenata or O. minor.
Two new benzonitrile glucosides, 4-hydroxymethylphenyl 6-(4-cyanophenyl)-β-D-gluco-pyranoside 183, termed brugmanioside A and 4-(hydroxyprop-2-en-3-yl)-phenyl 6-(4-cyanophenyl)-β-D-glucopyranoside 184, named brugmansioside B, were isolated from the flowers of Brugmansia arborea L. (Solanaceae), collected in Korea (Figure 19).116 When tested for cytotoxicity using human gastric adenocarcinoma cells (AGS) and human hepatocyte carcinoma cells (HepG2), the compounds exhibited moderate activity; at concentrations higher than 25 μg/mL (183) and 100 μg/mL (184) against AGS, and higher than 12.5 μg/mL (183) and 25 μg/mL (184) against HepG2.
Extraction of commercial Rhodiola crenulata L. (Crassulaceae) of Tibetan-origin afforded another benzonitrile, 2-cyano-3-hydroxy-4-methxyl-benzenemethanol-7-O-β-D-glucopyranoside 185, named crenulatanoside A (Figure 19), alongside the known nitriles sarmentosin and rhodiocyanoside A.117 Due to suggestions that Rhodiola spp. may lower blood glucose and represent potential treatments for diabetes, the compound was evaluated for its inhibitory activity against α-glucosidase but significant activity was not recorded.
Hamigeran R, 186 (Figure 19), considered to be the first benzonitrile-based marine natural product, was isolated from the New Zealand marine sponge Hamigera tarangaensis (PTN2_79F).118 When tested for cytotoxicity using the human promyelocytic leukemic (HL-60) cell line, the mean IC50 value for 186 was over 100 μM.
3-Indolecarbonitrile (187) was isolated from Salvadora persica L. (Salvadoraceae) (Figure 19), which is widely used for oral hygiene.119
Euphorbia dracunculoides Lam. (Euphorbiaceae), an annual herb of southwest Asia, North Africa, and South Europe was collected in Pakistan and various extracts were evaluated for antioxidant and anti-inflammatory activity. Due to the potent effects of the n-hexane extract on the aerial parts of the plant and the ability to reduce carrageenan-induced paw edema, the n-hexane extract was subject to GC-MS analysis. Among 30 constituents identified by comparison with mass spectral libraries, 0.4% of the nitrile dicyanoazulene 188 (Figure 19) was reported.120
Miscellaneous Nitriles
Another member of the tetracyclic ambiguine subclass of alkaloids, named 12-epi-ambiguine B nitrile, 189 (Figure 20), was isolated from Fischerella 52-1 from Lake Tennessee, USA, with the species determined as either F. ambigua or F. muscicola.121 Researchers were interested in whether the toxicity of cyanobacteria, particularly in relation to freshwater harmful algal blooms and eutrophication, could in part be related to teratogenic effects. Using the zebrafish embryo model to probe the potential teratogenicity of 189, it was observed that the compound had relatively limited effects on development, only at the highest concentrations tested (≥10 µg/mL), with a slight curvature of the body axis noted in 3 days postfertilization. Although there appeared to be an ability to recover from the teratogenic effects, the teratogenic effects of such compounds clearly limit their development as potential lead compounds. Notably, teratogenicity was observed at concentrations previously demonstrated to be required for antimicrobial activity within this class of compounds.
Miscellaneous nitriles.
The new nitrile derivative, 3α,4β-dihydroxy-6-oxo-1-cyclohexene-1-acetonitrile 190 (Figure 20), was isolated from the whole plant of Aquilegia ecalcarata Maxim (Ranunculaceae).122 The compound did not show any cytotoxic activity when tested against different classes of cancer cell lines, including GLC-82 (human pulmonary adenoma) and HCT (human large intestine carcinoma).
Extraction of the methanol extract of the white flowers of Impatiens balsamina L. (Balsaminaceae) yielded two new phenolic compounds 3-benzofuranacetonitrile, 2,3-dihydro-3-hydroxy-2-oxo-, (3S) 191 and 3-benzofuranacetonitrile, 3-(β-D-glucopyranosyloxy)-2,3-dihydro-2-oxo-, (3S) 192 (Figure 20), both containing a nitrile group.123 Compounds 191 and 192 did not show appreciable cytotoxicity using various human cancer cell lines (A549, SK-OV-3, SK-MEL-2, and HCT15), as evaluated using the sulforhodamine B assay, compared to the control drug, etoposide (IC50 values of 1.74, 1.96, 1.33, and 2.95 μM, respectively). The neuroprotective activity of the compounds was assessed by determining their effects on nerve growth factor (NGF) secretion in C6 cells, with 191 and 192 inducing NGF secretion by 155.6 and 136.7% respectively. Anti-neuroinflammatory activity by measuring NO production in lipopolysaccharide (LPS)-stimulated BV-2 cells, with compounds 191 and 192 showing IC50 values of 64.08 to 93.36 μM, with little effect on cell viability. In this regard, the compounds were 3- to 4-fold less potent than the control, L-NG-monomethyl-l-arginine (NMMA), with an IC50 value of of 20.53 μM.
Conclusions
In summary, this review aims to review gathered information about recently discovered natural nitrile-containing metabolites, and to classify them based on structural characteristics. Indeed, 192 molecules were identified, and are shown alongside their biological activities. It is evident that nitriles occur in diverse organisms and are elaborated within varied molecular architectures. The many interesting biological activities recorded for these compounds indicate their potential as “hit” compounds for medicinal applications, once toxicological, metabolic, and chemical properties are more clearly understood and optimized, and issues concerning sustainable sourcing and the development of scalable, efficient synthetic methods are addressed. This is of particular relevance to many of the marine nitriles discussed, which are retrievable in vanishingly small amounts—the spectroscopic characterization of 165 was initially performed on just 16.5 μg102! However, the rewards justify the effort to characterize and study these metabolites, as seen with the clinical success of the ecteinascidins. The development of tailored extraction techniques and formation of stabilized derivatives expands the potential of these compounds as drug-like entities. In addition, the commercialization of the ecteinascidins highlights the impact of the development of sustainable methodologies such as the use of more readily available precursors,32 an analogous strategy to that seen with other valuable natural products such as the taxanes.
Footnotes
Acknowledgments
We acknowledge RCSI for hosting CS on an Erasmus programme.
Author Contributions (Roles)
JB was responsible for conceptualization of the review. CS and JB both contributed to literature searches and to authorship of the paper. JB was responsible for editing the text.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Ethical Approval/Patient Consent
This research did not require ethical approval.
Informed Consent
Not applicable, because this article does not contain any studies with human or animal subjects.
Trial Registration
Not applicable, because this article does not contain any clinical trials.
ORCID iD
James Barlow
References
1.
SorokinDYvan PeltSTourovaTPTakaichiSMuyzerG. Acetonitrile degradation under haloalkaline conditions by Natronocella acetinitrilica gen. nov., sp. nov. Microbiology. 2007;153(4):1157‐1164.
2.
ArakawaT. Acetonitrile as solvent for protein interaction analysis. Int J Biol Macromol. 2018;114:728‐732.
3.
McConveyIFWoodsDLewisMGanQNancarrowP. The importance of acetonitrile in the pharmaceutical industry and opportunities for its recovery from waste. Org Process Res Dev. 2012;16(4):612‐624.
4.
SruthiPRAnasS. An overview of synthetic modification of nitrile group in polymers and applications. J Polym Sci. 2020;58(8):1039‐1061.
5.
FlemingFFYaoLRavikumarPCFunkLShookBC. Nitrile-containing pharmaceuticals: efficacious roles of the nitrile pharmacophore. J Med Chem. 2010;53(22):7902‐7917.
6.
WangXWangYLiXYuZSongCDuY. Nitrile-containing pharmaceuticals: target, mechanism of action, and their SAR studies. RSC Med Chem. 2021;12:1650‐1671.
7.
WangJLiuH. Application of nitrile in drug design. Chinese J Org Chem. 2012;32(9):1643‐1652.
8.
McKinneyRJDeVitoSC. Nitriles. In Kirk-Othmer Encyclopedia of Chemical Technology, (Ed.). 2000:https:doi.org/10.1002/0471238961.1409201813031109.a01
9.
ConnEE. Cyanogenic glycosides. The Biochemistry of Plants: A Comprehensive Treatise. 1981;7:479‐500.
10.
HuebelW. Nitriles in live organisms. Deutsche Apotheker Zeitung. 1981;121(17):863‐868.
11.
LegrasJLChuzelGArnaudAGalzyP. Natural nitriles and their metabolism. World J Microbiol Biotechnol. 1990;6(2):83‐108.
YulviantiMZidornC. Chemical diversity of plant cyanogenic glycosides: an overview of reported natural products. Molecules. 2021;26:719.
14.
EmsermannJKauhlUOpatzT. Marine isonitriles and their related compounds. Mar Drugs. 2016;14(1):16.
15.
BlaževićIMontautSBurčulF, et al.Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry. 2020;169:112100.
16.
Montes VidalDvon Rymon-LipinskiALRavellaS, et al.Long-chain alkyl cyanides: unprecedented volatile compounds released by Pseudomonas and Micromonospora bacteria. Angew Chem Int Ed Engl. 2017;356(15):4342‐4346.
17.
CompagnoneRSOliveriMCPiñaIC,et al.5-Alkylpyrrole-2-Carboxaldehydes From the Caribbean sponges Mycale microsigmatosa and Desmapsamma anchorata. Nat Prod Lett. 1999;13(3):203‐211.
18.
VenkateshamURama RaoMVenkateswarluY. New 5-alkylpyrrole-2-carboxaldehyde derivatives from the sponge Mycale tenuispiculata. J Nat Prod. 2000;63(9):1318‐1320.
19.
OrtegaMJZubiaMSanchezMCSalvaJCarballoJL. Structure and cytotoxicity of new metabolites from the sponge Mycale cecilia. Tetrahedron. 2004;60(11):2517‐2524.
20.
MaoS-CLiuYMorganJBJekabsonsMBZhouY-DNagleDG. Lipophilic 2,5-disubstituted pyrroles from the marine sponge Mycale sp. Inhibit mitochondrial respiration and HIF-1 activation. J Nat Prod. 2009;72(11):1927‐1936.
21.
SalaSFromontJGomezOVuongDLaceyEFlemattiGR. Albanitriles A-G: antiprotozoal polyacetylene nitriles from a Mycale marine sponge. J Nat Prod. 2019;82(12):3450‐3455.
CasparJSpitellerP. A free cyanohydrin as arms and armour of Marasmius oreades. Chembiochem. 2015;16(4):570‐573.
24.
XiaoJZhangQGaoYQShiXWGaoJM. Antifungal and antibacterial metabolites from an endophytic Aspergillus sp. Associated with Melia azedarach. Nat Prod Res. 2014;28(17):1388‐1392.
25.
MallikadeviTPaulsamySJamunaSKarthikaK. Analysis for phytoceuticals and bioinformatics approach for the evaluation of therapeutic properties of whole plant methanolic extract of Mukia maderaspatana (L.) M. Roem. (Cucurbitaceae) – A traditional medicinal plant in western districts of Tamil Nadu, India. Asian J Pharm Clin Res. 2012;5(4):163‐168.
26.
SaitoN. Chemical research on antitumor isoquinoline marine natural products and related compounds. Chem Pharm Bull (Tokyo). 2021;69(2):155‐177.
27.
FangYLiHJiB,et al.Renieramycin-type alkaloids from marine-derived organisms: synthetic chemistry, biological activity and structural modification. Eur J Med Chem. 2021;210:113092.
28.
SaitoNHiramatsuAHiradeH, et al.Chemistry of renieramycins. 16. Structure of 7–desmethylrenieramycin O ( = 14μ-hydroxyrenieramycin S) from blue sponge, Xestospongia sp. Heterocycles.2017;95(2):748‐752.
29.
ChamniSSirimangkalakittiNChanvorachotePSaitoNSuwanboriruxK. Chemistry of renieramycins. 17. A new generation of renieramycins: hydroquinone 5–O-monoester analogues of renieramycin M as potential cytotoxic agents against non-small-cell lung cancer cells. J Nat Prod. 2017;80(5):1541‐1547.
30.
HeW-FLiYFengM-T,et al.New isoquinolinequinone alkaloids from the South China Sea nudibranch Jorunna funebris and its possible sponge-prey Xestospongia sp. Fitoterapia. 2014;96:109‐114.
31.
CharupantKSuwanboriruxKAmnuoypolSSaitoEKuboASaitoN. Jorunnamycins A-C, new stabilized renieramycin-type bistetrahydroisoquinolines isolated from the Thai nudibranch Jorunna funebris. Chem Pharm Bull (Tokyo). 2007;55(1):81‐86.
32.
SuwanboriruxKCharupantKAmnuoypolSPummanguraSKuboASaitoN. Ecteinascidins 770 and 786 from the Thai tunicate Ecteinascidia thurstoni. J Nat Prod. 2002;65(6):935‐937.
33.
KimuraSPangkruangWYokoyaM,et al.Isolation, structure characterization, and synthesis of stabilized 1,2,3,4–tetrahydroisoquinoline marine natural product from potassium cyanide pretreated Thai tunicate, ecteinascidia thurstoni. Heterocycles. 2014;88(1):363‐375.
34.
WanXRenH-JDuM-N,et al.A new cyano-substituted anthracycline metabolite from Streptomyces sp. HS-NF-1006. J Antibiot. 2017;70:219‐221.
35.
GomesPBNettMDahseHMSattlerIMartinKHertweckC. Bezerramycins A-C, antiproliferative phenoxazinones from Streptomyces griseus featuring carboxy, carboxamide or nitrile substituents. Eur J Org Chem. 2010;20(2):231‐235.
36.
KalininaSAKalininDVHövelmannY,et al.Auranthine, a benzodiazepinone from Penicillium aurantiogriseum: refined structure, absolute configuration, and cytotoxicity. J Nat Prod. 2018;81(10):2177‐2186.
37.
MakarovSECurcićBPTesevićVV, et al.Defensive secretions in three species of polydesmids (Diplopoda, polydesmida, polydesmidae). J Chem Ecol. 2010;36(9):978‐982.
38.
VujisićLVVučkovićIMMakarovSE, et al.Chemistry of the sternal gland secretion of the Mediterranean centipede Himantarium gabrielis (Linnaeus, 1767) (Chilopoda: Geophilomorpha: Himantariidae). Naturwissenschaften. 2013;100(9):861‐870.
39.
AlaliFHudaibMAburjaiTKhairallahKAl-HadidiN. GC-MS Analysis and antimicrobial activity of the essential oil from the stem of the Jordanian toothbrush tree Salvadorapersica. Pharm Biol. 2005;42(8):577‐580.
40.
SalemMZMBehirySISalemAZM. Effectiveness of root-bark extract from Salvadora persica against the growth of certain molecularly identified pathogenic bacteria. Microb Pathog. 2018;117:320‐326.
41.
RajniakJBarcoBClayNKSattelyES. A new cyanogenic metabolite in Arabidopsis required for inducible pathogen defence. Nature. 2015;525:376‐379.
42.
LasekanOAzeezS. Chemo-preventive activities of common vegetables? Volatile organic compounds (VOCs). Pharm Anal Acta. 2014;5:306.
43.
KhanujaSPSAryaJSTiruppadiripuliyurRSK, et al.Nitrile glycoside useful as a bioenhancer of drugs and nutrients, process of its isolation from Moringa oleifera. 2005; United States Patent 6,858,588.
44.
DwivediGRMauryaAYadavDK,et al.Comparative drug resistance reversal potential of natural glycosides: potential of synergy niaziridin & niazirin. Curr Top Med Chem. 2019;19(10):847‐860.
45.
SuYFZhangZXGuoCYGuoDA. A nobel cyanogenic glycoside from Semiaquilegia adoxoides. J Asian Nat Prod Res. 2005;7(2):171‐174.
46.
ZhangHLiaoZXYueJM. Cyano- and nitro-containing compounds from the roots of Semiaquilegia adoxoides. Chinese J Chem. 2010;22(10):1200‐1203.
47.
WangZ-BGaoH-YYangC-JSunZWuL-J. Novel cyanoglucosides from the leaves of Hydrangea macrophylla. Helv Chim Acta. 2011;94(5):847‐852.
48.
WangZNakamuraSMatsudaHWuLYoshikawaM. New cyanoglycosides, hydracyanosides D, E, and F, from the leaves of Hydrangea macrophylla. Heterocycles. 2010;81(4):909‐916.
49.
Yamashita-HiguchiYSugimotoSMatsunamiKInagakiMOtsukaHTakedaY. Nitrile-containing phenolic glucosides from the leaves of Glochidion acuminatum. Chem Pharm Bull (Tokyo). 2015;63(1):49‐53.
50.
SimpolLROtsukaHOhtaniKKasaiRYamasakiK. Nitrile glucosides and rosmarinic acid, the histamine inhibitor from Ehretia philippinensis. Phytochemistry. 1994;36(1):91‐95.
51.
OhD-CWilliamsPGKauffmanCAJensenPRFenicalW. Cyanosporasides A and B, chloro- and cyano-cyclopenta[a]indene glycosides from the marine actinomycete “Salinispora pacifica”. Org Lett. 2006;8(6):1021‐1024.
52.
LaneALNamSJFukudaT,et al.Structures and comparative characterization of biosynthetic gene clusters for cyanosporasides, enediyne-derived natural products from marine actinomycetes. J Am Chem Soc. 2013;135(11):4171‐4174.
53.
DellagrecaMFiorentinoAMonacoPPreviteraLSimonetAM. Cyanogenic glycosides from Sambucus nigra. Nat Prod Lett. 2000;14(3):175‐182.
54.
SeiglerDSPauliGFNahrstedtALeenR. Cyanogenic allosides and glucosides from Passiflora edulis and Carica papaya. Phytochemistry. 2002;60(8):873‐882.
55.
MillerREStewartMCaponRJWoodrowIE. A galloylated cyanogenic glycoside from the Australian endemic rainforest tree Elaeocarpus sericopetalus (Elaeocarpaceae). Phytochemistry. 2006;67(13):1365‐1371.
56.
FuPZhaoC-CTangJShenY-HXuX-KZhangW-D. New flavonoid glycosides and cyanogenic glycosides from Dracocephalum peregrinum. Chem Pharm Bull. 2009;57(2):207‐210.
57.
NakamuraSWangSXuFMatsudaHWuLYoshikawaM. The absolute stereostructures of cyanogenic glycosides, hydracyanosides A, B, and C, from the leaves and stems of Hydrangea macrophylla. Tet Lett. 2009;50(32):4639‐4642.
58.
YangC-JWangZ-BZhuD-LYuYLeiY-TLiuY. Two new cyanogenic glucosides from the leaves of Hydrangea macrophylla. Molecules. 2012;17:5396‐5403.
59.
WangG-CLiTDengF-YLiY-LYeW-C. Five new phenolic glycosides from Hedyotis scandens. Bioorg Med Chem Lett. 2013;23(5):1379‐1382.
60.
BaatoucheSCherietTSarriD, et al.Centaurea microcarpa coss. & dur. (asteraceae) extracts: new cyanogenic glucoside and other constituents. Nat Prod Res. 2019;33(21):3070‐3076.
61.
NakamuraSLiXMatsudaH,et al.Bioactive constituents from Chinese natural medicines. XXVI. Chemical structures and hepatoprotective effects of constituents from roots of Rhodiola sachalinensis. Chem Pharm Bull. 2007;55(10):1505‐1511.
62.
MagdyMDMChristensenLPIbrahimNAAwadNEZeidIFPedersenEB. New acylated flavone and cyanogenic glycosides from Linum grandiflorum. Nat Prod Res. 2009;23(5):489‐497.
63.
MohammedMMDChenMZhaiLIbrahimNA. The cytotoxic activity of Linum grandiflorum leaves. Eur J Chem. 2010;1(2):110‐114.
64.
AgboMOLaiDOkoyeFBOsadebePOProkschP. Antioxidative polyphenols from Nigerian mistletoe Loranthus micranthus (linn.) parasitizing on Hevea brasiliensis. Fitoterapia. 2013;86:78‐83.
65.
SheG-MZhangY-JYangC-R. A new phenolic constituent and a cyanogenic glycoside from Balanophora involucrata (Balanophoraceae). Chem Biodivers. 2013;10(6):1081‐1087.
66.
YangQYSongLZhangJF,et al.Cyanogenetic glycosides and simple glycosides from the linseed meal. Fitoterapia. 2015;106:78‐83.
67.
SilvaTMSLinsACdSSarmento-FilhaMJ, et al.Riachin, a new cyanoglucoside from Bauhinia pentandra and its antioxidant activity. Chem Nat Compd. 2013;49:685‐690.
68.
de FariasPAFigueredoFGLucasAM, et al.Antibiotic-modifying activity of riachin, a non-cyanogenic cyanoglycoside extracted from Bauhinia pentandra. Drug Des Devel Ther. 2015;9:3067‐3072.
69.
MessangaBBTihRGSondengamBLBlondABodoB. Lanceolin C, a new nitrile glycoside from Lophira alata. Fitoterapia. 1998;69(5):439‐442.
70.
NkotJLBikoboDSNZintchemAAA, et al.Antitubercular evaluation of root extract and isolated phytochemicals from Lophira lanceolata against two resistant strains of Mycobacterium tuberculosis. Pharm Biol. 2018;56(1):318‐324.
71.
SaldanhaLLQuintiliano DelgadoAMarcourtL, et al.Hypoglycemic active principles from the leaves of Bauhinia holophylla: comprehensive phytochemical characterization and in vivo activity profile. PLoS One. 2021;16(9):e0258016.
72.
YiXXDengJGGaoCH, et al.Four new cyclohexylideneacetonitrile derivatives from the hypocotyl of mangrove (Bruguiera gymnorrhiza). Molecules. 2015;20(8):14565‐14575.
73.
Abouem à ZintchemABikoboDNde Théodore AtchadéA, et al.Nitrile glucosides and serotobenine from Campylospermum glaucum and Ouratea turnarea. Phytochemistry. 2008;69(11):2209‐2213.
74.
NjockGBBGrougnetREfstathiouA, et al.A nitrile glucoside and biflavones from the leaves of Campylospermum excavatum (Ochnaceae). Chem Biodivers. 2017;14(11).
75.
Abdel-MageedWMBayoumiSALAl-wahaibiLH, et al.Noncyanogenic cyanoglucoside cyclooxygenase inhibitors from Simmondsia chinensis. Org Lett. 2016;18(8):1728‐1731.
76.
Van BovenMBussonRCokelaereMFloGDecuypereE. 4-Demethyl Simmondsin from Simmondsia chinensis. Ind Crops Prod. 2000;12(3):203‐208.
77.
ShangX-YLiSWangS-JYangY-CShiJ-G. Chemical constituents of Bauhinia aurea. J Asian Nat Prod Res. 2012;14(10):966‐972.
78.
DongLChenMLiMLiaoZ-HSunM. A new cyanosides from Rhodiola bupleuroldes. Yaoxue Xuebao. 2009;44(12):1383‐1386.
79.
ChaiYQZhaoGHWangRJ,et al.[Anti-tumor metastatic constituents from Rhodiola wallichiana]. Zhongguo Zhong Yao Za Zhi. 2015;40(2):258‐263.
80.
WangHDongLGeJ-Q,et al.Rhobupcyanoside B, A new cyanoside from Rhodiola bupleuroides. J Asian Nat Prod Res. 2016;18(11):1108‐1114.
81.
OlsenCEMøllerBLMotawiaMS. Synthesis of the allelochemical alliarinoside present in garlic mustard (Alliaria petiolata), an invasive plant species in North America. Carbohydr Res. 2014;394:13‐16.
82.
GuanBLiTXuXK, et al.γ-Hydroxynitrile glucosides from the seeds of Prinsepia utilis. Phytochemistry. 2014;105:135‐140.
83.
ShiQ-QLuS-YPengX-RZhouLQiuM-H. Hydroxynitrile glucosides: bioactive constituent recovery from the oil residue of Prinsepia utilis. J Agric Food Chem. 2021;69(8):2438‐2443.
84.
SchwarzBHofmannT. Isolation, structure determination, and sensory activity of mouth-drying and astringent nitrogen-containing phytochemicals isolated from red currants (Ribes rubrum). J Agric Food Chem. 2007;55(4):1405‐1410.
85.
LuYYeap FooLWongH. Nigrumin-5-p-Coumarate and nigrumin-5-ferulate, two unusual nitrile-containing metabolites from black currant (Ribes nigrum) seed. Phytochemistry. 2002;59(4):465‐468.
86.
KimDKZeeOP. A new cyanogenic glycoside from Sorbaria sorbifolia var. stellipila. Chem Pharm Bull. 2000;48:1766‐1767.
87.
FangSYanXLiJFanZXuXXuR. Studies on the chemical constituents of Sedum sarmentosum bunge. IV. The structures of sarmentosine and isosarmentosine. Huaxue Xuebao. 1982;40(3):273‐280.
88.
BjarnholtNNakoniecznyMKędziorskiA,et al.Occurrence of sarmentosin and other hydroxynitrile glucosides in Parnassius (papilionidae) butterflies and their food plants. J Chem Ecol. 2012;38(5):525‐537.
89.
Fernandes JMTermentziAMandovaT, et al.Detection, isolation, and 1H NMR quantitation of the nitrile glycoside sarmentosin from Bryophyllum pinnatum hydro-ethanolic extract. J Agric Food Chem. 2021;69(29):8081‐8089.
90.
YangL-MHuRFuH-Z. A new cyano-compound from Rhodiola kirilowii. Chin Herb Med. 2011;3(4):241‐243.
91.
UemuraYSugimotoSMatsunamiK,et al.Microtropins A-I: 6′-O-(2″S,3″R)-2″-ethyl-2″, 3″-dihydroxybutyrates of aliphatic alcohol β-D-glucopyranosides from the branches of Microtropis japonica. Phytochemistry. 2013;87:140‐147.
92.
TerazawaSUemuraYKoyamaY, et al.Microtropins Q-W, ent-labdane glucosides: microtropiosides G-I, ursane-type triterpene diglucoside and flavonol glycoside from the leaves of Microtropis japonica. Chem Pharm Bull (Tokyo). 2017;65(10):930‐939.
93.
AvatoPTavaA. Rare fatty acids and lipids in plant oilseeds: occurrence and bioactivity. Phytochem Rev. 2021.
94.
AvatoPRositoIPapadiaPFanizziFP. Cyanolipid-rich seed oils from Allophylus natalensis and A. dregeanus. Lipids. 2005;40(10):1051‐1056.
95.
SinhaA. Chemical studies of cyanolipids in Koelreuteria apiculata. Int J Chem Stud. 2016;4(4Pt.C):216‐217.
96.
RoomiYASrivastavaVKHasanSQ. Cyanolipid in Lepisanthes tetraphylla seed oil. J Oil Technol Assoc India (Mumbai, India). 1998;30(2):65‐67.
97.
AvatoPRositoIPapadiaPFanizziFP. Characterization of seed oil components from Nephelium lappaceum L. Nat Prod Commun. 2006;1(9):751‐755.
98.
Ca LagoRPsc SimoniM de LdaCPintoA. Cyanolipids in Paullinia carpopodea cambess and P. cupana kunth seed oils. Acta Amazon. 2000;30(1):101‐105.
99.
TavaAAvatoP. Analysis of cyanolipids from Sapindaceae seed oils by gas chromatography-EI-mass spectrometry. Lipids. 2014;49(4):335‐345.
100.
RawatSNigamGSrivastavaVKHasanSNHasanSQ. Cyanolipids in Sapindus obovatus seed oil and reinvestigation of the seed oil of Heliotropium indicum, H. eichwaldi and D. viscosa. J Oil Technol Assoc India (Mumbai, India). 2002;34(2):69‐71.
101.
ZubíaEOrtegaMJCarballoJL. Sesquiterpenes from the sponge Axinyssa isabela. J Nat Prod. 2008;71(12):2004‐2010.
102.
DalisayDSMolinskiTF. Structure elucidation at the nanomole scale. 2. Hemi-phorboxazole A from Phorbas sp. Org Lett. 2009;11(9):1967‐1970.
103.
SmithABLiuZHoganAMLDalisayDSMolinskiTF. Hemi-phorboxazole A: structure confirmation, analogue design and biological evaluation. Org Lett. 2009;11(16):3766‐3769.
104.
MicallefMLSharmaDBunnBMGerwickLViswanathanRMoffittMC. Comparative analysis of hapalindole, ambiguine and welwitindolinone gene clusters and reconstitution of indole-isonitrile biosynthesis from cyanobacteria. BMC Microbiol. 2014;14(1):1‐18.
105.
MoSKrunicASantarsieroBDFranzblauSGOrjalaJ. Hapalindole-related alkaloids from the cultured cyanobacterium Fischerella ambigua. Phytochemistry. 2010;71(17–18):2116‐2123.
106.
HuberUMooreREPattersonGML. Isolation of a nitrile-containing indole alkaloid from the terrestrial blue-green alga Hapalosiphon delicatulus. J Nat Prod. 1998;61:1304‐1306.
107.
KimHKrunicALantvitD,et al.Nitrile-containing fischerindoles from the cultured cyanobacterium Fischerella sp. Tetrahedron. 2012;68(15):3205‐3209.
108.
LuZYangMChenPXiongXLiA. Total synthesis of hapalindole-type natural products. Angew Chem Int Ed. 2014;53(50):13840‐13844.
109.
KimJShinDKimS-H,et al.Borrelidins C–E: new antibacterial macrolides from a saltern-derived halophilic Nocardiopsis sp. Mar Drugs. 2017;15:166.
110.
YuMLiYBanakarSP,et al.New metabolites from the co-culture of marine-derived actinomycete Streptomyces rochei MB037 and fungus Rhinocladiella similis 35. Front Microbiol. 2019;10. https://www.frontiersin.org/article/10.3389/fmicb.2019.00915
111.
FagerholmAEHabrantDKoskinenAMP. Calyculins and related marine natural products as serine-threonine protein phosphatase PP1 and PP2A inhibitors and total syntheses of calyculin A, B, and C. Mar Drugs. 2010;8:122‐172.
112.
WakimotoTMatsunagaSTakaiAFusetaniN. Insight into binding of calyculin A to protein phosphatase 1: isolation of hemicalyculin A and chemical transformation of calyculin A. Chem Biol. 2002;9:309‐319.
113.
TorbatiMNazemiyehHLotfipourFAsnaashariSNematiMFathiazadF. Composition and antibacterial activity of Heracleum transcaucasicum and Heracleum anisactis aerial parts essential oil. Adv Pharm Bull. 2013;3(2):415‐418.
114.
LorenzPDucksteinSConradJKnödlerMMeyerUStintzingFC. An approach to the chemotaxonomic differentiation of two European dog’s mercury species: mercurialis annua L. And M. perennis L. Chem Biodivers. 2012;9(2):282‐297.
115.
CimminoAFernández-AparicioMAvolioFYoneyamaKRubialesDEvidenteA. Ryecyanatines A and B and ryecarbonitrilines A and B, substituted cyanatophenol, cyanatobenzo[1,3]dioxole, and benzo[1,3]dioxolecarbonitriles from rye (Secale cereale L.) root exudates: novel metabolites with allelopathic activity on Orobanche seed germination and radicle growth. Phytochemistry. 2015;109:57‐65.
116.
KimH-GKoJ-HOhHJ,et al.New cytotoxic benzonitrile glycosides from Brugmansia arborea flowers. Bull Korean Chem Soc. 2018:39(5);687-690.
117.
YangY-NLiuZ-ZFengZ-MJiangJ-SZhangP-C. Lignans from the root of Rhodiola crenulata. J Agric Food Chem. 2012;60(4):964‐972.
118.
WoollyEFSinghAJRussellERMillerJHNorthcotePT. Hamigerans R and S: nitrogenous diterpenoids from the New Zealand marine sponge Hamigera tarangaensis. J Nat Prod. 2018;81(2):387‐393.
119.
FaragMAFahmySChoucryMAWahdanMOElsebaiMF. Metabolites profiling reveals for antimicrobial compositional differences and action mechanism in the toothbrushing stick “Miswak” Salvadora persica. J Pharm Biomed Anal. 2017;133:32‐40.
120.
MajidMKhanMRShahNA, et al.Studies on phytochemical, antioxidant, anti-inflammatory and analgesic activities of Euphorbia dracunculoides. BMC Complement Altern Med. 2015;15(1):1‐15.
121.
WaltonKGantarMGibbsPDLSchmaleMCBerryJP. Indole alkaloids from Fischerella inhibit vertebrate development in the zebrafish (Danio rerio) embryo model. Toxins (Basel). 2014;6:3568‐3581.
122.
ChenSBGaoGYLiYSYuSCXiaoPG. Cytotoxic constituents from Aquilegia ecalcarata. Planta Med. 2002;68(6):554‐556.
123.
KimCSSubediLKimSY,et al.Two new phenolic compounds from the white flower of Impatiens balsamina. Phytochem Lett. 2015;14:215‐220.