
Abstract
Mucor circinelloides is an oleaginous fungus that utilizes a wide variety of carbon substrates for its growth. The different sources of carbon strongly influence the total lipid content of the fungus. These different carbon substrates are assimilated and dissimilated through different metabolic pathways before entering into the TAG synthesis pathway. In the present study, we attempted to explore the mechanism of ex-novo lipid biosynthesis in M. circinelloides WJ11 in response to exogenous plant oil as a carbon source through transcriptomic analysis. The lipid content of WJ11 grown in a media containing mixed soybean oil with glucose as a carbon source was up to 43.8%, an increase of 13.9% as compared to glucose alone as the carbon source. RNA-Seq analysis was performed to investigate global gene expression patterns in the oil-treated WJ11. Based on RNA-seq analysis, among the 4646 differentially expressed genes (DEGs), 2379 were up-regulated and 2267 down-regulated. The expression of acetyl-CoA synthetase, 6-phosphofructokinase, alcohol dehydrogenase (NADP+), fructose-bisphosphate aldolase, and pyruvate kinase was down-regulated while genes related to triglyceride synthesis were up-regulated. The majority of genes and pathways related to lipid biosynthesis were up-regulated indicating a diversion of metabolic pathways towards lipid biosynthesis. The data generated advance the genomic resources and provide insights into the mechanisms of ex-novo lipid accumulation in fungi that use exogenous oil as a carbon source.
The ability of some microorganisms to transform external carbon sources into lipids is considered a promising tactic for the production of essential fatty acid and biodiesel. 1 -3 Oleaginous microorganisms have features like easy cultivation, fast propagation, high oil content, and are easy to modify genetically. These microorganisms when grown in a high carbon and low nitrogen medium, accumulate more lipid in their cell dry weight. 4 The eventual goal of a single-cell oil is to produce nutritive supplements for animals and human food. Oleaginous fungi have, in the last few years, come under the spotlight due to their ability to produce fatty acids of high value while utilizing a wide range of cheap carbon sources for high-value lipid biosynthesis. 5 Fungi follow 2 general mechanisms for lipid synthesis: “ex-novo” when they use hydrophobic substrates such as cooking oils and “de-novo” when they use hydrophilic substrates such as glucose as a carbon source. 6 In oleaginous fungi, excess hydrophilic carbon and low nitrogen in the culture media play a vital role in de-novo lipid synthesis; under these conditions, the excess carbon is guided into lipid biosynthesis. 7 -10 On the other hand, ex-novo lipid accumulation is a growth-associated process that takes place at the same time as the cell grows. 11 When hydrophobic substrates (rich in plant oil) are given to oleaginous fungi, at first fats and oils are broken down into free fatty acids outside the fungal cells by lipase-catalyzed hydrolysis. 12 The free fatty acids, with the help of an active transport system, are transferred into the fungal cells. 13 Inside the cells, the free fatty acids are either degraded by β-oxidation for energy or biotransformed for the synthesis of new components (microbial lipids) of the cells. 14,15 In fungi, ex-novo lipid accumulation is independent of the presence of nitrogen in the culture medium and is associated with anabolic activities. 16 Among the huge number of fungal species, only some are known that have oleaginous properties and accumulate lipids up to 70% of their dry biomass. 17 Some microbial species that naturally produce PUFAs have been commercially explored and utilized. The importance of M. circinelloides, belonging to the Mucolares group of fungi, is of special interest to the lipid biotechnology industry due to its high total lipid content. 18,19 During the mid-1980s in the United Kingdom, M. circinelloides was reported as the first microorganism used commercially to produce polyunsaturated fatty acids like γ-linolenic acid (GLA). 20 M. circinelloides is considered as an important model organism to study lipid accumulation in oleaginous microorganisms. To overcome traditional limitations of low yield, various strategies of metabolic engineering have been used to modify M. circinelloides which made it very effective to produce high lipid contents. 21 A high lipid-producing strain, M. circinelloides WJ11, however, has been isolated from soil by our laboratory that produced up to 36% (w/w) lipid. 22 The genome of M. circinelloides WJ11 has been sequenced and comparative genomic approaches now provide an easy way to identify multiple genes that are expressed differentially. 22,23 In the present study, M. circinelloides WJ11 has been studied in medium supplemented with mixed plant oil and glucose as a carbon source. High biomass and lipid content were observed in the fungus in response to a mixed, oil and glucose carbon source. In this study, we have focused on the transcriptomics of M. circinelloides WJ11 grown in media with soybean oil to monitor lipid biosynthesis. The complete transcriptomics study seems essential to realize the regulatory mechanisms involved in the cascade of metabolic processes and the identification of targets of interest for further genetic and metabolic engineering. This study is the first attempt to explore the transcriptomics of M. circinelloides WJ11 and its metabolic shift from growth to lipid accumulation in response to a plant oil as a carbon source.
Materials and Methods
Culture Conditions
Mucor circinelloides WJ11 used in this experiment was isolated and preserved in Colin Ratledge Center for Microbial Lipids of Shandong University of Technology. 22 M. circinelloides WJ11 was grown in a specific modified lipid production K&R media. 24 The fungus was initially cultivated as a seed culture in K&R medium containing 30 g/L glucose, 1.5 g/L MgSO4•7H2O, 3.3 g/L ammonium tartrate, 7.0 g/L KH2PO4, 2.0 g/L Na2HPO4, 1.5 g/L yeast extract, 0.1 g/L CaCl2•2H2O, 8 mg/L FeCl3•6H2O, 1 mg/L ZnSO4•7H2O, 0.1 mg/L CuSO4•5H2O, 0.1 mg/L Co(NO3)2•6H2O, and 0.1 mg/L MnSO4•5H2O. Seed culture was prepared by inoculating 100 µL of WJ11 spores (107spores/mL) in a 1 L baffled flask containing 250 ml of K&R seed media and was incubated at 28 °C for 24 hours with shaking at 150 rpm. 10% (v/v) seed culture was used to inoculate a 4 L fermenter (New Brunswick, an Eppendorf company) containing 2.5 L modified K&R fermentation medium. The composition of the fermentation medium for the control was 80 g/L glucose and 2 g/L ammonium tartrate; the rest of the ingredients remained the same as the seed media. In the treatment group (plant oil as carbon source) the modified K&R fermentation medium was prepared with mixed carbon sources, 35 g/L glucose and 24.3 g/L soybean oil (average molecular weight of soybean oil is 835 g/mol and glucose 180 g/mol, therefore 45 g glucose was replaced with 24.3 g/L to maintain the equal molarity of carbon in the control and treatment groups). Fermenters were controlled at 28℃ with stirring at 700 rpm, aeration at 2 v/v min-1 and pH 6 for 4 days.
Biomass and Lipid Analysis
Fermentation samples were collected at 24, 48, 72, and 96 hours, and filtered through a Buchner funnel. The resulting biomass was washed 3 times with normal saline, frozen at −80℃ overnight and then dried in a vacuum freeze-dryer. The cell dry weight of the control and treated strain was determined gravimetrically. Lipid extraction was achieved by acid hydrolysis following the protocol described by Folch et al., with some modifications. 25 The lipid content was calculated according to the following formula:

RNA Extraction, Library Construction, and Sequencing
Based on growth and lipid accumulation characteristics, fermentation samples collected at 24 hours were selected for RNA analysis. Samples were filtered through a Buchner funnel to collect cell biomass. This was immediately rinsed 3 times with PBS buffer and placed in liquid nitrogen for 15 minutes. Then, frozen samples were stored at −80℃ for further experiments. Total RNA from treated samples of M. circinelloides WJ11 was isolated using TRIzol, according to the manufacturer’s instructions (Invitrogen Life Technologies). Total RNA was processed by the mRNA enrichment method. mRNA with polyA tail was enriched by using Oligo dT magnetic beads and the requisite RNA was collected after purification. The purified RNA in the presence of interrupted buffer was cut into shorter fragments and the resulting RNA was subjected to reverse transcription by using random N6 primers to synthesize first and second-strand cDNA. After end repair and the addition of a poly (A) tail, the cDNA was ligated with adaptors and the ligated product was amplified by PCR using specific primers to construct a cDNA library template. Finally, the cDNA library was sequenced by the MGISEQ2000 genetic sequencer (Shenzhen, China).
Transcriptome Mapping, Annotation, and Differential Expression Analysis
The original sequencing data contain reads with low quality, high N content of unknown bases and contaminated linkers. Before data analysis, the low-quality reads were removed to ensure the reliability of the results. The sequenced data were filtered with SOAPnuke (v1.5.2). 26 The reads containing the sequence of the adapter were deleted, those with a low-quality base ratio (base quality less than or equal to 5) greater than 20% were removed, as were reads with an unknown base ("N" base) ratio greater than 5%. The clean readings were stored in FASTQ format. HISAT2 (v2.0.4) 27 was used to locate the clean reads against the reference genome (https://www.ncbi.nlm.nih.gov/genome/browse/#!/eukaryotes/87669/). Bowtie2 (v2.2.5) 28 was used to compare the clean reads with the reference coding gene set, and then the gene expression level was calculated by RSEM (v1.2.12). 29
Quantitative gene and various analyses based on gene expression levels (principal components, correlation, differential gene screening) were performed, and gene ontology (GO) functional significance enrichment analysis on the differentially expressed genes among the selected samples, pathway significant enrichment analysis, clustering, protein interaction network and transcription factors and other in-depth mining analysis.
Differentially expressed genes (DEGs) between groups were identified by the DEseq2, which is based on the principle of a negative binomial distribution. DEGs detection was carried out according to the method described by Michael et al. 30,31 All expressed genes were functionally annotated against the NCBI non-redundant protein (NR) database using the Blast algorithm with a cut-off of E-value ≤10−5. Genes were also subjected to classification and enrichment analysis of the GO functions and KEGG pathways. Gene ontology (GO) classification and Kyoto Encyclopedia of Genes and Genomes (KEGG) classification were performed using Blast2go software and KAAS (KEGG automatic annotation server), respectively. A hypergeometric test was used to identify the GO terms of a significant difference in DEGs between the 2 groups. The KEGG pathways enriched with DEGs compared to the reference set were also analyzed by the hypergeometric test.
Results and Discussions
Effects of Mixed Soybean Oil and Glucose on the Growth and Lipid Accumulation of M. Circinelloides
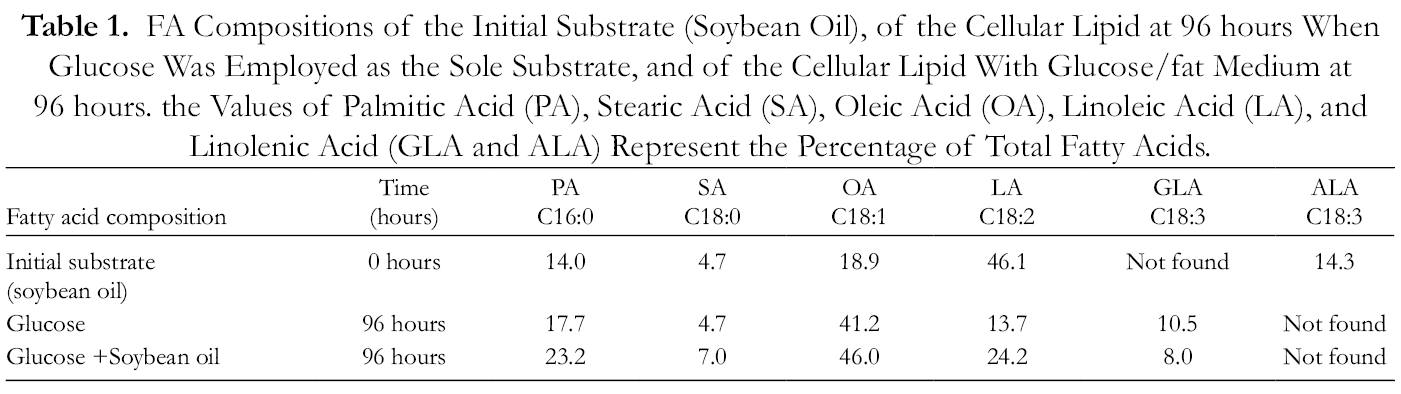
M. circinelloides WJ11 is a promising oleaginous fungus that can be cultivated on different carbon sources including waste oils or fats for high-valued oil production. To investigate the influence of exogenous plant oil as the carbon source on cell growth and lipid accumulation of M. circinelloides WJ11, soybean oil was tested at different concentrations by the shake flask method. It was found that a concentration of 24.3 g/L of soybean oil mixed with 35 g/L glucose led to the highest growth and lipid accumulation, and, therefore this concentration of oil and glucose was used for further experiments. The effect of mixed carbon sources on the growth and lipid accumulation of M. circinelloides was further studied in a fermentor. From the fermentation studies, it was observed that the cell dry weight (CDW) of the fungus grown in a medium with exogenous oil was higher than that of the glucose (control) (Figure 1(A)). The resultant dry biomass of M. circinelloides WJ11 after fermentation was examined for its total lipid content. The total lipid content of M. circinelloides WJ11 grown in the mixed medium was up to 43.8%, increased by 13.9% compared to glucose (Figure 1(B)). On glucose based media, this strain presented appreciable cell growth, with maximum biomass production after 48 hours of fermentation. After nearly 96 hours, total glucose exhaustion was observed in the culture medium. Additionally, when M. circinelloides was cultivated on an oil-glucose based media, this fungal strain was capable of producing high biomass, after 48 hours of fermentation. After nearly 48 hours, total glucose exhaustion was observed, while 96% of other available carbon sources (soybean oil) were utilized at ~96 hours of fermentation time. The addition of fatty materials employed as sole substrates or co-substrates used together with sugars has been shown to increase the cellular lipid content in various types of fungi. 32 -36 On the other hand, in some cases, modelling lipid production during growth on blends of sugars and fats has been successfully attempted. 36 Some earlier studies have indicated the potential of lipid production and the successful simulation and prediction of the process when M. circinelloides strains have been cultured on fatty materials that positively influence lipid accumulation. 37 The total lipid of M. circinelloides grown on glucose and glucose/oil blends was also subjected to gas chromatography for their fatty acid composition. From the GC results, the major fatty acids found were palmitic, stearic, oleic, linoleic, and γ-linolenic acids (Table 1). A major change in the profile of different fatty acids was observed in M. circinelloides grown on the glucose-oil mixed media. The content of most of the fatty acids, except GLA, increased; the highest increase was observed for linoleic acid. M. circinelloides has been reported to be an excellent SCO-producer, with high conversion yields of sugar/oil into lipid. Lipids produced on a glucose-oil media contain γ-linolenic acid (GLA), a polyunsaturated fatty acid (PUFA) odietary and pharmaceutical importance, though in low concentration as compared with glucose as the only carbon source. Fats and hydrophilic substrates have been used as carbon sources for cultivating Zygomycetes for the production of valuable fatty acids. 38 -40 In addition, our results are consistent with previous studies that have shown that oils such as coconut, palm, and other vegetable oils showed positive effects on fungal biomass and lipid accumulation. 41,42

Growth and lipid accumulation of WJ11 cultivated in different media. (A) The cell dry weight of strains. (B) The lipid content as a percentage of dry cell weight.
FA Compositions of the Initial Substrate (Soybean Oil), of the Cellular Lipid at 96 hours When Glucose Was Employed as the Sole Substrate, and of the Cellular Lipid With Glucose/fat Medium at 96 hours. the Values of Palmitic Acid (PA), Stearic Acid (SA), Oleic Acid (OA), Linoleic Acid (LA), and Linolenic Acid (GLA and ALA) Represent the Percentage of Total Fatty Acids.
RNA Sequencing, Differentially Expressed Genes (DEGs) and Functional Annotations
There have been many studies on the mechanism of lipid accumulation in M. circinelloides. 22 The genome of M. circinelloides WJ11 is completely available and comparative genomic approaches now provide an easy way to identify multiple genes that are expressed differentially. The cDNA library of fermentation samples after 24 hours has been sequenced. The obtained samples were subjected to clean reads; HISAT was used to align the clean reads to the reference genome sequence. A summary of the RNA-seq reads is shown in Supplemental Table S1. We have obtained 21.65 million and 21.67 million clean reads on average in the control group (glucose as carbon source) and treatment group (plant oil and glucose mixture as carbon source). Combined, these reads accounted for more than 96.7% of the total reads, indicating good sequencing quality. After filtering out the ribosomal RNA (rRNA), approximately 96.74% and 97.02% of reads from the control group and treatment group respectively were mapped to the reference genome.
Various carbon sources taken into the microbial cell go through different yet integrated pathways into the main anabolism toward TAGs synthesis. To study the gene regulation mechanism at the molecular level, the raw sequencing data of the sample were first purified and then aligned to the reference genome. Based on the RNA-seq analysis, 4646 differentially expressed genes (DEGs) were identified in the dataset of the treatment group compared to the control group (Figure 2(A)). From the results, it was observed that there were 2379 genes up-regulated and 2267 down-regulated in the treatment group compared with the control group (Figure 2(B)).

Distribution of differentially expressed genes (DEGs) between the exogenous oil treatment group and control group. (A) the volcano graph; (B) the distribution of up-regulated and down-regulated genes; (C) GO annotation and classification of DEGs; (D) Top 20 KEGG pathways in which DEGs are significantly enriched.
Gene ontology (GO) enrichment analyses for the DEGs were conducted. The results show GO functional classifications and annotations that elucidate the biological functions and overall distributions of all the DEGs. The GO analyses are classified into biological process, cellular component, and molecular function. These 3 main categories included 38 subcategories (Figure 2(C)). Due to its multiple functions, one gene could be annotated into numerous different categories. 43 In the biological processes category, metabolic process and cellular process were the 2 dominant classes, containing 967 and 1164 DEGs, respectively. In the cellular component category, cell (946 genes) and membrane (1294 genes) and membrane part (1249 genes) were the main classes, while binding (1568 genes) and catalytic activity (1693 genes) classes were prominent in the molecular function category.
KEGG pathway annotations and enrichment analyses were conducted for DEGs to identify the major changes in biological pathways involved in M. circinelloides WJ11 grown on different media. Three significant enrichment pathways (Q-value <0.05) for DEGs were identified: ribosome, MAPK signaling pathway-yeast, and longevity regulating pathway-multiple species. Also, several pathways with important functions appeared in the top 20 of the enriched KEGG pathways, such as glycolysis/gluconeogenesis, glycerophospholipid metabolism, glycerolipid metabolism, fatty acid metabolism, and fatty acid degradation (Figure 2(D)).
Key Pathways and DEGs Involved in Response to Exogenous Oil
Based on analyses of annotation, GO enrichment, and KEGG pathway enrichment for DEGs, we selected some important pathways of carbon metabolism like fatty acid metabolism, glycolysis, pentose phosphate pathway, and TCA cycle to illustrate further the multiple response mechanisms of M. circinelloides WJ11 to the exogenous oil at the transcriptional level. Glycolysis and pentose phosphate pathways are 2 important ones for the catabolic metabolism of glucose. The down-regulated DEGs associated with the glucose metabolism pathways are shown in Supplemental Table S2. From the results, it was found that many genes involved in glycolysis have been down-regulated (Figure 3). The down regulation of this pathway was probably due to the addition of exogenous oil. The expression of acetyl-CoA synthetase, 6-phosphofructokinase, alcohol dehydrogenase (NADP+), fructose-bisphosphate aldolase, and pyruvate kinase genes was significantly lower in the fungus cultured in soybean oil and glucose mixture as a carbon source. Among these genes, 2 encoding acetyl-CoA synthetase were down-regulated 22.03- and 21.93-fold, and one gene encoding 6-phosphofructokinase was down-regulated 21.74- fold. Also, the pentose phosphate pathway is another branch of glucose catabolism. Glucose is broken down by the pentose phosphate pathway to produce erythrose 4-phosphate. Glucose-6-phosphate dehydrogenase (G6PDH) and 6-phosphogluconate dehydrogenase (6PGDH) are the critical rate-limiting enzymes of the pathway. One gene encoding 6-phosphogluconate dehydrogenase was down-regulated 21.78- fold indicating that the cells use less glucose for de-novo lipid synthesis, but use exogenous fatty acids for ex-novo lipid synthesis. This result indirectly confirms that when fungi and yeasts are cultivated in mixed glucose and exogenous oil as the carbon source, the mechanism of lipid accumulation is different. 44,45 The strain grown in the glucose-oil mixed medium can use both carbon sources during the cell growth stage. Therefore exogenous oil not only provides a carbon skeleton for cell growth but also provides energy.

DEGs involved in the glycolysis pathway of M. circinelloides WJ 11 grown for 24 hours. Red color represents up-regulation, and green represents down-regulation of genes.
Differentially Expressed Genes Related to Lipid Accumulation
From the analysis of the pathway of glycerolipid metabolism, it was found that many genes have been up-regulated related to triglycerides synthesis (Figure 4). One gene encoding glycerol-3-phosphate O-acyltransferase was up-regulated 22.39-fold; this enzyme adds acyl groups to glycerol, the initial step for triacylglycerides synthesis. Besides, the up-regulation of genes was also detected for 2 enzymes involved in the conversion of diacylglycerides to triacylglycerides. These results confirmed that the addition of exogenous oil induces TAG synthesis, which in turn is responsible for increased lipid accumulation in WJ11. The genes of 2 enzymes involved in the intracellular degradation of triacylglycerides and diacylglycerides were down-regulated. Two genes encoding TAG lipase were down-regulated 21.76-and 21.70-fold. In addition, the gene encoding fatty acid synthase subunit beta (FAS2) was down-regulated; FAS2 catalyzes the formation of long-chain fatty acids from acetyl-CoA, malonyl-CoA and NADPH. However, the fatty acid synthesis gene (FabG) was up-regulated; FabG enzymes are members of the short-chain alcohol dehydrogenase/reductase (SDR) family that catalyze a wide range of NAD(H) and NADP(H) dependent oxidoreduction reactions. In the fatty acid degradation pathway, we also found that most of the genes were up-regulated. This study suggested that exogenous oil is hydrolyzed and assimilated by the fungus. The fatty acids produced from the hydrolysis of exogenous oil are oxidized for energy, as well as also being converted to triacylglycerides by an ex-novo pathway for storage.

DEGs involved in glycerolipid metabolism of M. circinelloides WJ 11 grown for 24 hours.
Conclusion
In the present study, lipid accumulation and gene expression of M. circinelloides WJ11 during batch cultivation on mixed plant oil-glucose as a carbon source was explored. This study highlights genes potentially involved in the ex-novo lipid synthesis of oleaginous fungi at the transcriptomic level. Overall growth and lipid accumulation of the treated strain were fast and high in the presence of exogenous oil as compared to glucose. During the early growth phase (24 hours), genes specific to TAG biosynthesis were up-regulated. This work provides a foundation for understanding the metabolic capabilities of M. circinelloides WJ11 necessary for growth and lipid accumulation on a complex carbon source (oil), with household waste or hydrophobic substrates being considered.
Supplemental Material
Table S1 - Supplemental material for Transcriptome Analysis of Oleaginous Fungus Mucor circinelloides WJ11 in Response to Exogenous Soybean Oil as Carbon Source
Supplemental material, Table S1, for Transcriptome Analysis of Oleaginous Fungus Mucor circinelloides WJ11 in Response to Exogenous Soybean Oil as Carbon Source by Caili Sun, Aabid Manzoor Shah, Junhuan Yang, Zongmin Wang, Lanlan Zhu and Yuanda Song in Natural Product Communications
Footnotes
Acknowledgments
The authors thank all members of the Colin Ratledge Center of Microbial Lipids, Shandong University of Technology, School of Agriculture Engineering and Food Sciences for their scientific assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the National Natural Science Foundation of China (Grants Nos. 31972851 and 31670064) and TaiShan Industrial Experts Programme (No. tscy20160101).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.