
Abstract
The stratum corneum tryptic enzyme kallikrein 5 (KLK5) is a serine protease that is involved in the cell renewal and maintenance of the skin barrier functions. The excessive activation of KLK5 causes an exacerbation of dermatoses, such as rosacea and atopic dermatitis. Brassica napus play a well-known role in the treatment of canola oil through their anti-oxidative and DNA protective properties. We aimed to investigate whether the bioactive ceramide modulate the KLK5 protease. The ceramides were evaluated using an enzymatic assay to measure the anti-KLK5 activity. Our study revealed that the ceramides modulate the KLK5 and 7 protease activity. Ceramides may affect the skin barrier and atopic function via the regulation of proteases.
Stratum corneum tryptic enzyme kallikrein 5 (KLK5) is a serine protease, expressed in the epidermis and involved in cell renewal and maintenance of the skin barrier function. 1,2 Kallikrein 5 plays a central role in the degradation of corneodesmosomes, which are the main adhesive structures in the cornified cell layer. Kallikrein 5 is also involved in the activation of other epithelial serine proteases such as kallikrein- and matriptase-family proteases. 3 Although KLK5 is important for the maintenance of skin homeostasis, its overactivation can impair the skin barrier function and contribute to the discovery of various dermatoses. 4 Several studies have reported certain triterpenoids that are bioactive natural products and suppress the serine protease activity. Plant-derived pentacyclic triterpenoids, such as oleanolic acid, ursolic acid, and β-boswellic acid, have been reported to inhibit the esterase activity in human neutrophils. 5 Brassica napus is one of the world’s most economically important products as an oilseed. Brassica napus has been used in China to treat benign prostatic hyperplasia. The previous studies showed that the ethyl acetate extract of B. napus has sterols, terpenoids, flavones, long chain hydrocarbons, and brassinolide. 6 Sphingolipids, like ceramides and cerebrosides, are important constituents of the cellular membrane, and are emerging as important second messengers for various cellular processes such as the cell cycle arrest, differentiation, senescence, and apoptosis. 7 Although B. napus is widely known for its actions in dietary supplements and cosmetics, its effects on atopic dermatitis are yet to be investigated. Therefore, we investigated the efficacy of a C24 ceramide from B. napus on KLK5 and KLK7 protease activity. The C24 ceramide (Figure 1) was obtained as an amorphous white powder, and the structure was determined to be (2S,3S,4R,8E)-2-[(2′R)-2′-hydroxytetracosenoilamino]-8-octadecene-1,3,4-triol). 8 Next, we examined the potential of the ceramide and ursolic acid to inhibit the KLK5 activity at a concentration of 10 µM (Table 1). Ceramide and ursolic acid (10 µM) inhibited the hydrolysis of a synthetic substrate of purified KLK5, demonstrating inhibition percentages of 38.2% and 54.1%, respectively. The positive control agent, leupeptin hemisulfate, completely inhibited KLK5 activity at 42.1 µM. The 50% inhibitory concentration (IC50) values reported for ceramide and ursolic acid were 13.8 and 8.5 µM, respectively. Notably, the inhibitory effect of the ceramide on KLK5 activity was dose dependent. We further investigated the selectivity of the ceramide and ursolic acid against KLK5 and KLK7 protease activity. As shown in Table 2, ursolic acid inhibited KLK5 and trypsin activities; the IC50 values were 5.6 and 12.8 µM, respectively. In contrast, the IC50 values against KLK7 and chymotrypsin C activities were >100 µM. Ursolic acid exhibited a moderate inhibitory effect on KLK7 and chymotrypsin C at a maximal concentration of 100 µM (37.8% and 9.2%, respectively). The ceramide demonstrated IC50 values of 10.2 and 15.4 µM against KLK5 and trypsin proteases, respectively, and exhibited a weak inhibitory effect on KLK7 and chymotrypsin C proteases (21.3% and 5.3%, respectively, at 100 µM). This is the first report to clarify the effects of naturally occurring ceramides on human KLK5 protease. The ceramide suppressed KLK5 activity. Furthermore, KLK5 is involved in the processing of cathelicidin-related antimicrobial peptides in epidermal keratinocytes. 9 Our findings suggest that ceramides regulated the proteolytic processing of KLK5 and inhibited KLK5 and trypsin activities but not KLK7 and chymotrypsin C activities. This indicated that these ceramides are more selective against trypsin-like serine proteases (KLK5) rather than chymotrypsin-like serine proteases (KLK7). As the antiprotease activity was detected at a relatively low concentration of <100 µM, and chymotrypsin-like serine protease (KLK7 and chymotrypsin C) activity was partially inhibited at 100 µM, the ceramides could possibly exhibit an antiprotease activity against other proteases at higher concentrations. Ursolic acid is used as an additive in cosmetic ingredients. Ceramides act on multiple target molecules, and it is plausible that the above effects of ceramides were modulated through target molecules such as PPAR-alpha and kinase enzymes. 10 Although the present study could not clarify and discover the relationship of ceramides with multiple molecular targets, well-designed future studies assessing the intermolecular interactions could promote the development of new KLK5 drug inhibitors. Our findings have demonstrated future possibilities and the necessity to conduct further research to elucidate the role of ceramides as therapeutic agents in refractory skin diseases such as rosacea and atopic dermatitis.
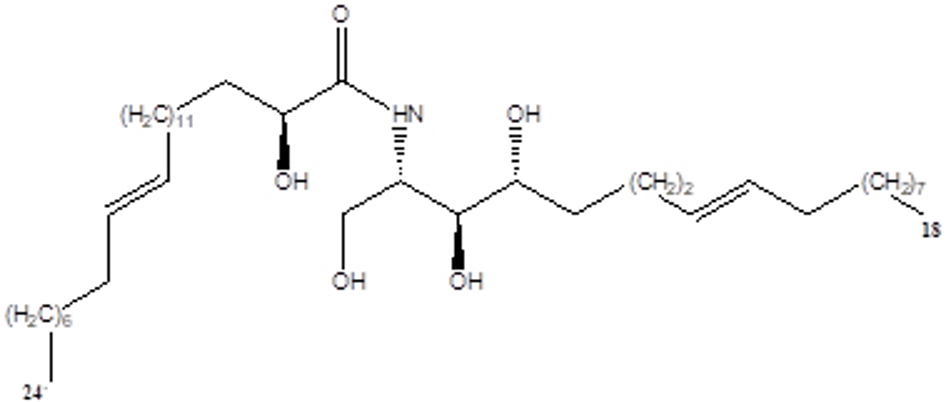
Chemical structures of ceramide from the seed of Brassica napus L.
Assay to Ceramide With Anti-Kallikrein 5 Activity.

Recombinant human kallikrein 5 (KLK5, 8.1 nM) was mixed with 10 µM test compounds and Boc-Val-Pro-Arg-AMC fluorogenic peptide (100 µM). After incubating for 5 minutes, the relative fluorescent unit (RFU) was measured at Ex 380 nm/Em 460 nm. A serine protease inhibitor, leupeptin hemisulfate (42.1 µM), was used as positive control. Ceramides showing an activity of more than 35% inhibition vs control were statistically significant at each concentration by Dunnett’s test. Data are presented as the mean ± SEM of triplicate tests. The percentage of inhibition was calculated using the following formula: (1 − (A − B)/(C − B)) × 100, where A is the test sample RFU, B the basal RFU without KLK5, and C the vehicle RFU.
aUrsolic acid is a commercially available standard.
bDexamethasone is a negative standard.
cLeupeptin hemisulfate salt is a positive standard.
Comparative Activities of Ursolic Acid and Tumulosic Acid Against Serine Proteases.
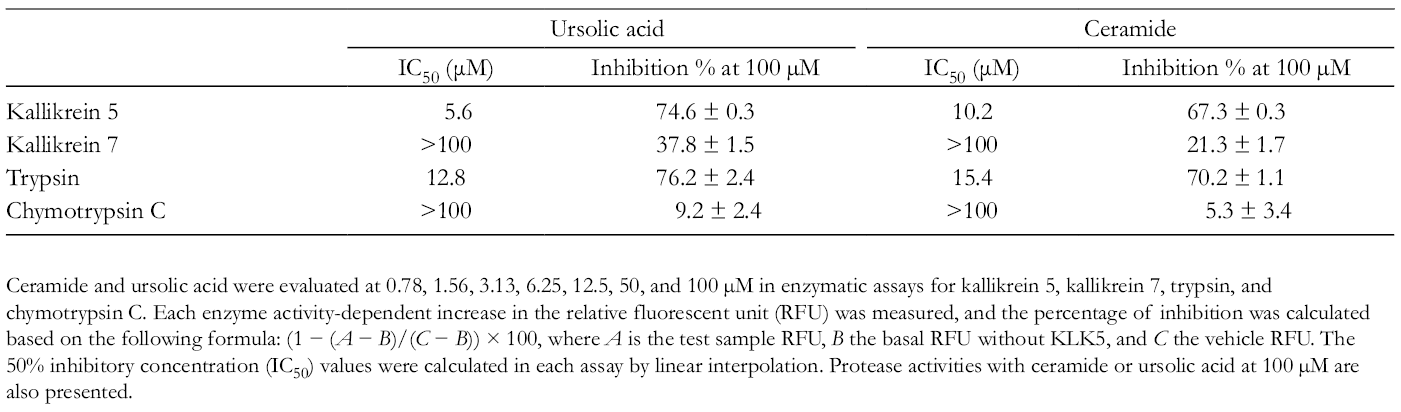
Ceramide and ursolic acid were evaluated at 0.78, 1.56, 3.13, 6.25, 12.5, 50, and 100 µM in enzymatic assays for kallikrein 5, kallikrein 7, trypsin, and chymotrypsin C. Each enzyme activity-dependent increase in the relative fluorescent unit (RFU) was measured, and the percentage of inhibition was calculated based on the following formula: (1 − (A − B)/(C − B)) × 100, where A is the test sample RFU, B the basal RFU without KLK5, and C the vehicle RFU. The 50% inhibitory concentration (IC50) values were calculated in each assay by linear interpolation. Protease activities with ceramide or ursolic acid at 100 µM are also presented.
Experimental
General
The seeds of B. napus L. were purchased in November 2018 in G-market, Seoul, South Korea. A voucher specimen (Skedrm20181105) has been deposited at the raw material room, SKEDERM cosmetic R&D center, South Korea. Nuclear magnetic resonance (NMR) spectra were recorded on a Varian Inova-400 FT-NMR spectrometer (CA, United States) with TMS as an internal standard, δ in ppm, J in Hz. High-resolution electrospray ionization mass spectrometry were measured on Bruker APEXII mass spectrometer in m/z. EI-MS were measured on a VG ZABHS mass spectrometer at 70 eV. Silica gel (200-300 mesh) were obtained from Merck Co. Ltd. Ursolic acid was purchased fromWako Pure Chemical Industries (Osaka, Japan). Dexamethasone (Wako Pure Chemical Industries), leupeptin hemisulfate salt and chymostatin (Sigma-Aldrich, St Louis, MO, United States) were purchased for use as reference regents.
Extraction and Isolation
The air-dried seeds of B. napus L. (2 kg) were extracted with 70% EtOH (3 × 20 L, 7 days each) at room temperature and the EtOH was removed under reduced pressure to give a residue (612 g), which was suspended in distilled water and extracted with n-hexane and EtOAc, respectively. The EtOAc extract (36 g) was subjected to column chromatography over silica gel (200-300 mesh, 2500 g) and eluted with CHCl3; CHCl3-MeOH (95/5), (90/10), (85/15), (80/20), (70/30), (50/50), (30/70), and (10/90); and MeOH to yield 10 fractions (Fr.1-Fr.10). Fr.2 to Fr.4 were subjected to a silica gel column eluting with CHCl3-MeOH (19/1) to give compound
Measurements of Enzymatic Activity
Kallikrein 5, KLK7, trypsin, and chymotrypsin C activities were evaluated according to the following methods supplied by R&D Systems Inc. (Minneapolis, MN, United States). The enzyme activity-dependent increase in the relative fluorescent unit (RFU) was measured, and the percentage of inhibition was calculated based on the following formula: (1 − (A − B)/(C − B)) × 100, where A is the RFU of test samples with enzymes, B the basal RFU without enzymes, and C the RFU of vehicle controls with enzymes.
To measure the KLK5 activity, enzymatic reaction was performed at room temperature in 100 mM NaH2PO4 buffer (pH 8.0) containing 0.25 µg/mL recombinant human KLK5 (R&D Systems Inc.), 100 mM of Boc-V-P-R-AMC Fluorogenic Peptide Substrate (R&D Systems Inc.), and 1.1% DMSO at final concentrations. Kallikrein 5 (final 8.1 nM) was preincubated with test samples for 5 minutes, followed by the addition of peptide substrate. After incubating for 5 minutes, RFU was measured at Ex 380 nm/Em 460 nm. Leupeptin hemisulfate (42.1 µM) was used as a positive control.
To measure the KLK7 activity, recombinant human pro-KLK7 (R&D Systems Inc.) was activated by bacterial thermolysin at 37°C for 2 hours just before the enzyme assay. Thereafter, enzymatic reaction was performed at room temperature in 50 mM Tris, 150 mM NaCl buffer (pH 8.5) containing 1 µg/mL activated-KLK7, 10 µM Mca-R-P-K-P-V-E-Nval-W-R-K (Dnp)-NH2 Fluorogenic Peptide Substrate II (R&D Systems Inc.), 150 mM NaCl, and 1.1% DMSO at final concentrations. Activated KLK7 (final 38.5 nM) was preincubated with test samples for 5 minutes, followed by the addition of peptide substrate. After incubating for 60 minutes, RFU was measured at Ex 320 nm/Em 405 nm. Chymostatin (10 µM) was used as a positive control.
To measure the trypsin activity, enzymatic reaction was performed at room temperature in 100 mM NaH2PO4 buffer (pH 8.0) containing 0.25 µg/mL recombinant human trypsin (Wako Pure Chemical Industries), 100 µM of Boc-V-P-R-AMC Fluorogenic Peptide Substrate, and 1.1% DMSO at final concentrations. Kallikrein 5 was preincubated with test samples for 5 minutes, followed by the addition of peptide substrate. After incubating for 1 minute, RFU was measured at Ex 380 nm/Em 460 nm. Leupeptin hemisulfate (42.1 µM) was used as a positive control.
To measure the chymotrypsin C activity, recombinant human pro-chymotrypsin C (R&D Systems Inc.) was activated by trypsin at 37°C for 1 hour just before the enzyme assay. Thereafter, the enzymatic reaction was performed at room temperature in 25 mM Tris, 0.5 mM CaCl2 buffer (pH 8.0) containing 1 µg/mL activated chymotrypsin C, 10 µM Suc-A-A-P-F-AMC (Bachem AG, Bubendorf, Switzerland) used as a fluorogenic substrate, and 1.1% DMSO at final concentrations. Activated chymotrypsin C (final 34.7 nM) was preincubated with test samples for 5 minutes, followed by the addition of peptide substrate. After incubating for 60 minutes, RFU was measured at Ex 380 nm/Em 460 nm. Chymostatin (10 µM) was used as a positive control.
Statistical Analysis
All of the data are presented as means ± standard deviation. Statistical analyses used the one-way analysis of variance followed by Dunnett’s test. P < 0.05 was considered to be statistically significant.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was carried out with the support of Classys INC. (Project title: Construction of high purity, concentration extracts techniques from natural materials).