
Abstract
Cryptocarya alba is an important tree species in the Chilean sclerophyllous forest. Its leaves and bark are used in traditional medicine to treat liver diseases and rheumatism. Analyses of the essential oil (EO) show serious discrepancies, and information on other constituents is limited. The aerial biomass of individual trees from 3 wild populations, some old trees, and farmed saplings were analyzed (n = 132). The EO profiles were studied by gas chromatography/mass spectrometry (GC/MS). The alkaloidal and polyphenol compositions were determined by ultra-high-performance liquid chromatography (UHPLC)/MS-MS. The total polyphenol content, the total flavonoid content, and the antioxidant capacity (diphenylpicrylhydrazyl, azinobisethylbenzothiazolinesulfonic acid, and ferric reducing antioxidant power: DPPH, ABTS, and FRAP respectively) were determined by standard methods. Significant differences were found at the individual and population levels in the contents of polyphenols, total flavonoids, antioxidant capacity, and specific alkaloids for leaves, bark, and wood. Farmed saplings grown under less light showed higher concentrations of higenamine, N-methylcoclaurine, N-methyllaurotetanine, and isocorydine, while those receiving more light were richer in laurolitsine, boldine, coclaurine, catechin, quercetin, epicatechin, quercitrin, and procyanidins. Important variations were found according to the season, age of the tree and of the leaves. The EO composition also varied considerably. These results support the idea that the natural variability of medicinal species is an important subject for study.
Cryptocarya alba (Mol.) Looser (Lauraceae) is an endemic evergreen tree of Chile known by the vernacular name “peumo.” It grows between the south of the Coquimbo Region (about 31°S) and the north of the Lake Region (about 40°S). It is most abundant in the center of its range, in the Mediterranean-type climate zone. 1 It can sprout from the roots after severe fires 2 and is able to survive damage caused mainly by European rabbits (Oryctolagus cuniculus). The trees cover an area in excess of 9 million hectares, forming mixed forests associated mainly with Lithraea caustica and Quillaja saponaria, and less frequently with Peumus boldus and Aristotelia chilensis. 3 Cryptocarya alba grows mainly in humid and shaded places like ravines where it may form pure stands. Nevertheless, it suffers from conservation problems in the drier northern part of its range. 1 It plays an important ecological role in the landscape. Many insects feed on its sap and wood; 5 indigenous rodent species and 5 bird species depend on its pale pink to red fruits, which are also a regular component of the diet of the Pseudalopex culpaeus fox. 1 This tree is used by humans for firewood and carpentry, and its leaves and bark have been used in different preparations for liver ailments, rheumatic pains, 4 and wound healing. 5 In at least one prehistoric cultural complex (500-1.000 CE), the leaves were smoked in clay pipes together with an unidentified wild tobacco (Nicotiana sp.). 6 The fruit has been collected since pre-Columbian times by the Mapuche people 7 and has potential as a functional food. 8
The Lauraceae family is a rich source of alkaloids, mainly of the 1-benzyltetrahydroisoquinoline and aporphine types. 9 Until a couple of years ago, C. alba leaves and bark were only known to contain reticuline, 4 but low concentrations of aporphines have been identified recently. 10 The fruit contains cryptofolione and a methoxylated derivative (α-pyrones). 11 The essential oil (EO) of the leaves analyzed presents variations. It was first analyzed in 1988, with terpinen-4-ol (14.1%), p-cymol (7.3%), and eucalyptol (6.6%) identified as major constituents. 12 A C. alba EO containing mainly α-terpineol (27.4%), eucalyptol (23.3%), and phellandrene (16.3%) exhibited antifungal activity. 13 Another sample containing terpinen-4-ol (28.1%), β-terpinene (23.1%), p-cymene (15.9%), and eucalyptol (18.9%) was found to be slightly bactericidal. 14 Yet another one, containing terpinen-4-ol (17.5%), 4-(3,3-dimethylbut-1-ynyl)-4-hydroxy-2,6,6-trimethyl-cyclohex-2-enone (12.8%), eucalyptol (7.9%), and p-cymene (7.1%) showed an insecticidal effect. 15 A very different C. alba EO, composed mainly of (E)-β-bergamotene (15.6%), viridiflorol (8.5%), and germacrene-D (7.6%), demonstrated activity as a corn weevil repellent. 16
Three common flavonoids and their glycosides, largely quercitrin, were isolated from the aerial parts of C. alba. 5 More than 30 phenolic compounds have been identified in these materials, where chlorogenic acid was highlighted. 7 In another recent study, the quercetin content practically doubled that of chlorogenic acid, and a poorly identified kaempferol derivative was also noteworthy. 17 In addition, catechin, epicatechin, procyanidins, and chlorogenic acids have been reported as leaf constituents, with large differences between different provenances and seasons. 18 Quercetin and chlorogenic acid appear to be major secondary metabolites of C. alba leaves. Quercetin is an abundant dietary flavonoid that is claimed to provide multiple health benefits. 19 Geographic latitude seems to affect the content of metabolites such as leaf EOs and alkaloids of P. boldus, 20,21 and the terpenes of Drimys winteri, 22 and may be the case in other Chilean species. 23
We have now examined the leaf, bark, and wood, assaying total polyphenols, total flavonoids, antioxidant potential, and separate alkaloids and polyphenols, and analyzed the leaf EO of individual C. alba trees from 3 natural populations and from a tree farm. We conclude that, in spite of their broad variation in secondary metabolite composition, the leaves are the most interesting organ of C. alba considering possible medicinal or industrial applications, followed by the bark and, finally, the wood. The content of leaf polyphenols varied from 54 to 131 mg gallic acid equivalents (GAE)/g dry weight (DW). The flavonoid content was between 8.5 and 21.9 mg
The alkaloid composition of the branches (wood and bark) is largely consistent with results obtained previously. 10 In 67% of the individuals, laurolitsine was the most abundant wood alkaloid, while laurotetanine and reticuline predominated in the bark. In 88% of the trees, reticuline was the major bark alkaloid, but exceptions were found in the southern population, with laurotetanine predominating in the bark of 6 individuals, and reticuline in the wood of another 7, while only 2 individuals had both high bark laurotetanine and wood reticuline. The population growing in the Metropolitan Region (R M) exhibited significant differences in the laurotetanine and laurolitsine contents of the wood with regard to the other 2 populations (Figure 1), and the Curanipe population (R VII) showed the lowest concentration of reticuline.

Concentration of alkaloids in wood of Cryptocarya alba. Different letters in columns correspond to significant differences according to Dunn’s test (P < 0.01).
Analysis of the bark of the R M trees gave significant differences in the contents of reticuline as compared with the other 2 populations. The lowest values for laurotetanine and boldine were found in the northernmost population (R V) (Figure 2). The content of laurotetanine in the bark was positively correlated with reticuline (R2 = 0.79, P < 0.0005) and N-methyllaurotetanine (R2 = 0.80, P < 0.0005), while laurolitsine was also correlated with boldine (R2 = 0.86; P < 0.0005) (Supplementary Table S2). In the wood, these alkaloid contents were also correlated, but to a lesser degree (Supplementary Table S3). The highest concentration of reticuline found in the bark of C. alba branches was 287 µg/g DW. Nevertheless, it is possible to find higher concentrations in this species, for example, in old trees, where some individuals had more than 500 µg/g DW. 10 Although higher concentrations have been reported in California poppy (Eschscholzia californica), 24 in the roots of Litsea cubeba, 25 and in the wood and bark of boldo (P. boldus), 26 a significant advantage of C. alba bark is that reticuline is the major alkaloid in this abundant species, making its purification simpler. Both reticuline enantiomers are intermediates in the biosynthesis of most isoquinoline alkaloids and are therefore very interesting as precursors in potential biotechnological processes. S(+)-reticuline is a dopamine receptor antagonist that produces catalepsy and a decrease in locomotor activity. 27,28 It also has GABAA activity 24 and is a powerful central nervous system depressor. 29 It inhibits platelet aggregation 30 and has anti-inflammatory and antinociceptive activity. 31

Concentration of alkaloids in bark of Cryptocarya alba. Different letters in columns correspond to significant differences according to Dunn’s test (P < 0.01).
In our earlier work on C. alba, the best results for leaves were obtained from young saplings (<3 years old). 10 Therefore, a detailed analysis of farmed individuals was deemed important. The amount of sunlight can influence the yield of some alkaloids so we compared subpopulations with different degrees of shading. In areas with less light (80% shade), we found higher concentrations of higenamine, N-methylcoclaurine, N-methyllaurotetanine, and isocorydine. In contrast, with 40% shade, laurolitsine, boldine, and coclaurine were more abundant (Figure 3). In the sunnier area, individuals grew larger leaves, but no differences were found in the yield of leaves per plant (Supplementary Table S5).

Concentration of alkaloids in leaves of Cryptocarya alba saplings grown under different degrees of shade. Different letters in columns correspond to significant differences according to *Tukey’s test (P < 0.01) and **Dunn’s test (P < 0.05).
The low concentrations of alkaloids found in C. alba leaves make it unlikely that these components are responsible for any of the reported activities, such as antioxidant capacity or antimutagenicity in Drosophila.
17
There is enough information to attribute greater relevance to phenolic compounds due to the abundance of chlorogenic acid, quercetin, and quercitrin. Gallic acid (
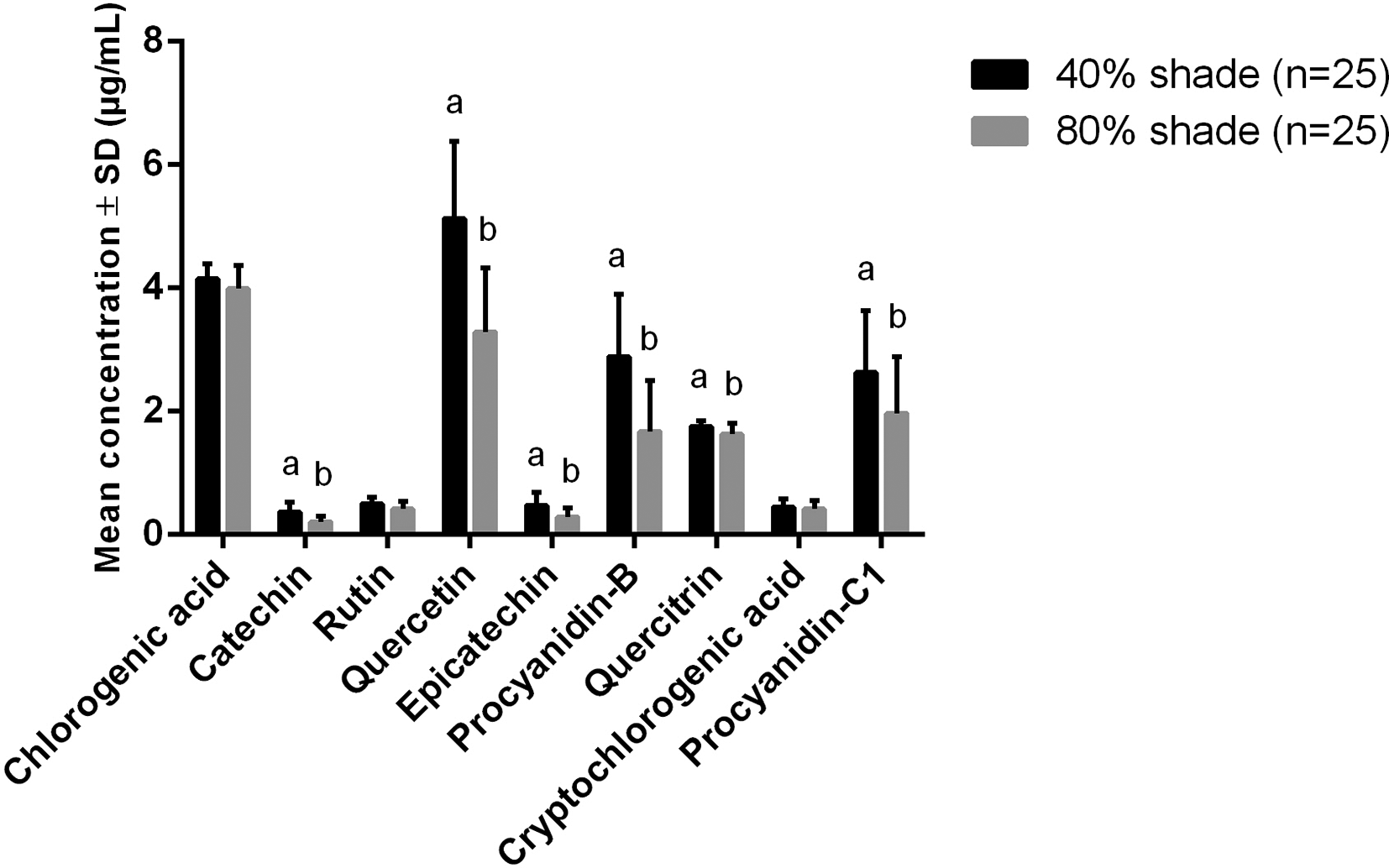
Phenolic compounds in the aqueous extract of Cryptocarya alba leaves from saplings grown under different degrees of shade. Different letters in columns correspond to significant differences according to Student’s t test (P < 0.05).
Fully developed C. alba leaves (adult leaves/old leaves) have a higher content of polyphenols, total flavonoids, and greater antioxidant capacity compared with soft new leaves (Supplementary Figure S6 and Table S4), with important differences for quercetin and procyanidins (Figure 5).

Phenolic compounds in the aqueous extracts of new and old leaves. Different letters in columns correspond to significant differences according to Student’s t test (P < 0.05).
Leaves of C. alba from the R M population showed significant variations in chlorogenic acid and quercitrin, but not other phenolics, according to the season of collection. If the age of the tree is considered, the leaves of long-lived individuals contain significantly more chlorogenic acid than those of younger trees in any season. Nevertheless, the higher (and therefore colder) location of the older trees might also be responsible for the difference found. Leaves of older trees also showed less variation. Important differences were also evidenced in the 8 sampling seasons for total phenolic compounds (TPC) (Supplementary Figure S7). Year-long fluctuations have been documented before for C. alba, with TPC and condensed tannins increasing in winter, and cis-chlorogenic acid in spring. 18 Although quercetin and rutin did not exhibit significant seasonal variation in the R M population, they varied considerably from one individual to another. Pacheco noted the total loss of cis-chlorogenic acid, epicatechin, caffeic acid, ferulic acid, and dihydroxybenzoic acid from C. alba in summer, 18 while in our analysis this constituent increased slightly but significantly in summer. Procyanidins were markedly higher in spring, decreasing with the advancing seasons (Figure 6).

Concentration of polyphenols in the aqueous extract of Cryptocarya alba leaves from 3 collections of younger trees in María Pinto and long-lived trees in Curacaví. Different letters in columns correspond to significant differences according to Dunn’s test (P < 0.05).
The secondary metabolites found in the highest amounts in bark and wood of C. alba were procyanidins. This agrees with the traditional use of C. alba bark to tan leather and for medicinal purposes, 4 even though the concentrations of these constituents may vary considerably. 18 These compounds have antimicrobial, antioxidant, and anti-inflammatory activities, 34 and procyanidin B2 has been shown to prevent cognitive decline and oxidative damage in mice. 35,36 In pigs, grape pomace procyanidins (mostly total a dimer and a trimer, possibly of procyanidin C2 improve antioxidant total status (TAS) and reduce lipid peroxidation (thiobarbituric acid reducing substances: TBARS). 37 Procyanidins are very low in the bark of long-lived trees, compared with full-grown trees of intermediate age, and also in the wood of one of these populations (Supplementary Figure S8). There are also considerably higher (average) amounts of these metabolites in the La Dormida pass (R V) population compared with the María Pinto (MR) population (Figure 7).

Phenolic compounds in the aqueous extracts of the bark of Cryptocarya alba. Different letters in columns correspond to significant differences according to * Dunn’s test (P < 0.05), **Unpaired t test (P < 0.05), and ***Mann-Whitney test (P < 0.05).
Cryptocarya alba is an important species in the Chilean sclerophyllous forest, and its aroma is often quite noticeable while walking in the woods. The volatile organic compounds emitted by C. alba are said to be composed mainly of eucalyptol, limonene, β-myrcene, α-pinene, and camphene. 38 Different reports on the leaf EO of C. alba show different major constituents, which may be terpinen-4-ol, α- terpineol, or β-bergamotene. Such differences, like those shown recently for C. mandioccana of the Brazilian Atlantic Forest, may be induced by environmental influences or may have a genetic basis, although the available information is insufficient to clarify this point. 39
Table 1 shows the most abundant and prevalent constituents of EO prepared from 8 different samples of leaves of C. alba growing in the city of Santiago (Chile). Sixteen compounds were present in at least 4 of the samples in a relative concentration ≥1%. In 6 of the 8 samples, sabinene was the first or second principal component (13.5 ± 2.8%), and in the other, 2 camphene (20.6% and 22.7 %) followed by β-eudesmol (6.1 ± 5.7%) and eucalyptol (5.4 ± 1.5%) were most abundant. The sabinene content was always higher than that recorded (6.8%) for a sample collected on the western limit of the Santiago valley (Lo Prado pass), while the eucalyptol content of this wild-growing material was similar (7.9%). 15
Main Constituents of the Leaf Essential Oil of Cryptocarya alba From Santiago City (Chile) (n = 8).

Ki = Kovats index; max = maximum; min = minimum; n = number of samples; Ref = references; RT = retention time; SD = standard deviation.
In conclusion, leaves are the most attractive component of the analyzed aerial biomass from a chemical viewpoint, with higher antioxidant capacity and higher phenolic contents. They are rich sources of quercetin, quercitrin, isorhamnetin, and chlorogenic acid. Our data suggest that the highly variable concentration of these compounds may be affected by the amount of light, the age of the tree, and collection season, all factors that should be considered for the quality control of this medicinal plant. The bark is rich in procyanidins and may be a useful source of (S)-reticuline, but the occurrence of these metabolites seems to depend on the age of the tree. At the population level, the great differences observed justify the continued study of some of these aspects and serious consideration of analytical quality control when harvesting material for medicinal purposes if reproducibility is an issue.
Experimental
Plant Material
Three wild populations of C. alba were selected in central Chile, one near the boundary between the Valparaíso Region (R V) and the Santiago Metropolitan Region, on La Dormida pass (Cuesta La Dormida, 33°03′S, 71°02′W, 820 m above sea level), one in María Pinto borough in the Metropolitan Region (R M) (33°26′S, 71°18′W, 230 masl), and a third near Curanipe in the Maule Region (R VII) (35°55′S, 72°38´W, 122 masl). Samples were collected from 25 individuals in each one of these populations, in January and February 2014 (the height of summer), taking materials from around each tree, which were then meticulously separated into leaves, wood, and bark, dried for 72 hours at 40°C, ground, and labeled. Leaves and bark of older trees (diameter at breast height, DBH >40 cm) were collected in the mountains in Curacaví borough (33°22′S, 70°58′W, 950 masl). More samples were collected from 100 saplings growing in an experimental farm in San Bernardo, on the outskirts of Santiago (33°39′S and 70°43′W, at an altitude of about 550 masl). Of the latter, 50 3.5-year-old individuals were sampled for alkaloid analysis and 50 4.5 year olds for polyphenol analysis. The species was identified by M. Sc. Gloria Rojas Villegas, and a voucher specimen was deposited at the Herbarium of the Museo Nacional de Historia Natural de Chile (No. 169 644 SGO).
Chemicals and Reference Standards
N-Methyl-laurotetanine was isolated from the leaves of Duguetia vallicola. 40 Racemic norreticuline, coclaurine, and N-methylcoclaurine were synthesized. 41 Racemic norcoclaurine (higenamine) was prepared by demethylation of coclaurine, 42 and (S)-glaucine was prepared from boldine by methylation with diazomethane. 43
Sample Preparation
Methanol Extracts
All dried plant materials (leaves, wood, and bark) were ground and passed through a 2-mm screen. A total of 1 ± 0.01 g aliquots were suspended in MeOH (100 mL), stirred for 2 hours at 60°C, and filtered through Whatman No. 1 paper. The solids collected were subjected to the same procedure 4 times. The combined extracts were concentrated in a rotary evaporator at 40°C under reduced pressure and stored until use in amber vials at -20°C. For analysis, they were resuspended in MeOH (20 mL).
Aqueous Extracts
A total of 1 ± 0.01 g of ground material (leaves, wood, or bark) was placed in a flask, and distilled water (150 mL) was added. The mixture was stirred briefly about every 45 seconds for 5 minutes, in a bath kept at 95°C. The extracts were gravity-filtered through Whatman No. 1 paper, and their volume adjusted to 150 mL.
Extraction of EOs
Samples of 400 ± 0.05 g of fresh C. alba leaves were collected in 3 different University campuses within not more than 10 km of each other in Santiago, Chile. The EO was obtained by hydrodistillation for 4 hours, dried with anhydrous sodium sulfate, and stored in amber vials at −20°C.
Quantification of TPC and Flavonoids
Polyphenol and flavonoid contents were quantified in accordance with the methods of Singleton and Rossi 44 and Chang et al. 45
Liquid Chromatography and Mass Spectrometry
All the samples were injected into an ultra-high pressure liquid chromatograph (Eksigent model EkspertUltraLC 100-XL) coupled to a triple quadrupole mass spectrometer in the electrospray mode (ESI) (ABSciex Triple Quad 4500). A Phenomenex Synergi™ Fusion-RP 80 Å (50 mm 2.0 mm, 4 µm) column was employed injecting 10 µL aliquots. The LC-MS/MS system was controlled by Analyst 1.6.2 and the data were processed using Multiquant 3.0.
Quantification of Alkaloids
The mobile phase was aqueous formic acid 0.1% v/v (eluent A) and acetonitrile (eluent B) at a flow rate of 0.3 mL/min. The gradient program 26 was as follows: (time, min/%B) 3/3%, 13/15%, 16/20%, 17/3%, and 18/3%. The mass spectrometer parameters were GS1 N2, 40 psi; GS2 N2, 50 psi; IS, 3500 V, TEMP, 650°C; and CURT N2, 25 psi. Calibration curves were built with concentrations in the 0.1-0.8 µg/mL range.
Quantification of Polyphenol
The mobile phase was aqueous formic acid 0.1% v/v (eluent A) and acetonitrile (eluent B) at a flow rate of 0.5 mL/min. The gradient program 34 was as follows: (time, min/%B) 1/5%, 10/40%, 11/100%, 13/5%, and 14/5%.
Gas Chromatographic Analysis
The GC/MS analysis was performed using a Shimadzu model GCMSQP 2010 Ultra gas chromatograph (Shimadzu, Kyoto, Japan), with an Rtx-5MS Crossbond 5% diphenyl-95% dimethyl polysiloxane (Restek, Bellefonte, PA, USA) capillary GC column (30-m length, 0.25 mm I.D., and 0.25 mm film thickness). The GC was operated in the splitless injection mode and the injection volume was 5 µL. The column temperature was held at 35°C for 5 minutes, raised at a rate of 10°C/min to 200°C, and kept for 10 minutes at 200°C. The carrier gas was He at a flow rate of 1.3 mL/min.
Antioxidant Capacity
DPPH Assay
A solution of 150 µM DPPH in MeOH was prepared and incubated in the dark for 2 hours. Then, a 20 µL aliquot of each extract was added to 2980 µL of DPPH solution, and after 30 minutes, the absorbance was read at 520 nm. 46 The percentage of inhibition was determined according to [(Ao – Ae)/Ao] × 100, where Ao is the absorbance without extract and Ae is the absorbance with extract. The calibration curve was built using Trolox (y = 1538.5x + 4.6594; R² = 0.9987). The results are expressed in μmol Trolox equivalent.
ABTS Assay
The ABTS+ radical was prepared by the reaction of ABTS (7 mM) and potassium persulfate (2.45 mM), incubated at room temperature in the dark for 16 hours. The radical solution was diluted with EtOH to obtain an absorbance value of 0.70 (±0.02) at 734 nm. 47 A 50 µL aliquot of each individual extract was added and allowed to react for 7 minutes and the absorbance was then read at 734 nm. The percentage of inhibition was determined according to [(Ao − Ae) / Ao] × 100, where Ao is the absorbance without extract and Ae is the absorbance with extract. The calibration curve was built using Trolox (y = 5.343x – 0.1006; R² = 0.9985). The results were expressed in µmol Trolox equivalent.
FRAP Assay
The FRAP assay was carried out according to the method of Benzie and Strain. The ferric complex was formed by mixing the following reagents: 25 mL sodium acetate buffer (300 mM, pH 3.6), 2.5 mL solution TPTZ (10 mM), and 2.5 mL ferric chloride (20 mM) in a ratio of 10:1:1. A 20 µL aliquot of each individual extract was added, and after 40 minutes at 37°C, the absorbance was read at 593 nm The calibration curve was built using Trolox (y = 0.1208x + 0.0277; R² = 0.9994). The results were expressed in µmol Trolox equivalent. 48
Data Analysis
The distribution of each variable was examined using the Shapiro-Wilk goodness of fit test. To compare the alkaloid concentrations between different collection points, in wood, bark, and leaves, and also to compare the leaf phenolics (40%-80% shade, new/old leaves, from María Pinto and Curacaví), Kruskal-Wallis and Dunn’s multiple comparison tests were used. Dunn’s test, unpaired t-test, and Mann-Whitney test were used in the analysis of bark phenolics. The association between different secondary metabolites was studied with Pearson’s correlation test for normal distributions of the variables and with Spearman’s test when the distribution was not normal. In all statistical tests, significance was set at α < 0.05. STATA 10.1, SPSS 17, and GraphPad Prism 6 statistical packages were used.
Supporting Information
Total polyphenol content, total flavonoid content, and antioxidant capacity of the leaves, wood, and bark of 3 populations (Supplementary Figure S1-S5), correlations between wood metabolites (Supplementary Table S1), bark and wood alkaloids (Supplementary Table S2 and S3), differences in polyphenolic and flavonoid contents, and antioxidant capacity of new and old leaves (Supplementary Figure S6 and Table S4), variation in total polyphenol and total flavonoid contents in 8 collection periods (Supplementary Figure S7), polyphenolic concentration in wood (Supplementary Figure S8), dimensions of the leaves of 3.5-year-old farmed C. alba saplings with 40% or 80% shading (Supplementary Table S5), UHPLC-MS/MS calibration curve for alkaloids (Supplementary Table S6) and polyphenols (Supplementary Table S7), EO profile of a sample of C. alba leaves, and GC-MS parameters of the identified compounds (Supplementary Table S8) are available.
Supplemental Material
Supplementary data - Supplemental material for Variation of Secondary Metabolites in the Aerial Biomass of Cryptocarya alba
Supplemental material, Supplementary data, for Variation of Secondary Metabolites in the Aerial Biomass of Cryptocarya alba by Ady Giordano, Gonzalo Fuentes-Barros, Sebastián Castro-Saavedra, Antonia González-Cooper, Cristian Suárez-Rozas, Julio Salas-Norambuena, Williams Acevedo-Fuentes, Felipe Leyton, Cristian Tirapegui, Javier Echeverría, Sandra Claros, and Bruce K. Cassels in Natural Product Communications
Footnotes
Acknowledgments
Thanks are due to Dr Edwin G. Pérez of the Facultad de Química of Pontificia Universidad Católica de Chile for a gift of N-methyllaurotetanine.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Fondo de Investigación del Bosque Nativo (No. 055/2013) and a FONDEQUIP grant (No. EQM 150102).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.