
Abstract
Pancreatic cancer ranks among the most lethal digestive malignancies, exhibiting a steadily increasing incidence and mortality worldwide. Despite significant advances in cancer research, the 5-year survival rate remains below 10%, predominantly due to delayed diagnosis and limited therapeutic options. Concurrently, the gut microbiota—an integral component of host physiology—has emerged as a crucial player in the pathogenesis of pancreatic cancer. Mounting evidence indicates that alterations in gut microbial composition and function may influence tumor initiation, progression, and response to therapy. This review provides an in-depth examination of the intricate interplay between the gut microbiome and pancreatic cancer, highlighting potential diagnostic biomarkers and exploring microbiome-targeted therapeutic strategies to improve patient outcomes.
Plain language summary
Pancreatic cancer is a deadly disease with few early symptoms and limited treatment options. Recent research suggests that the bacteria living in our gut, known as gut microbiota, may play a role in the development and treatment of pancreatic cancer. This review looks at how changes in gut microbiota could influence the onset and progression of pancreatic cancer. We also explore new possibilities for using gut bacteria to help diagnose pancreatic cancer earlier and develop better treatment options. Understanding the relationship between gut microbiota and pancreatic cancer could lead to new strategies to improve patient outcomes and increase survival rates.
Introduction
Pancreatic cancer ranks among the most lethal malignancies of the digestive system, with pancreatic ductal adenocarcinoma (PDAC)—originating from the exocrine pancreas—comprising more than 85% of all malignant pancreatic tumors. 1 Globally, its incidence and mortality continue to rise. In the European Union (EU), for instance, projections suggest that by 2025 approximately 111,500 individuals will succumb to pancreatic cancer—an increase of nearly 50% compared with mortality rates reported in 2010—making pancreatic cancer the third leading cause of cancer-related deaths after lung and colorectal cancers. 2 In China, improving living standards and changing dietary habits have likewise contributed to a rising incidence and mortality rate, with the 5-year survival rate remaining below 10%, thereby posing a significant public health threat.1,3
The dismal survival rate of pancreatic cancer largely reflects its frequent late-stage diagnosis. Given its limited sensitivity to chemotherapy, early surgical intervention remains the primary strategy for improving patient outcomes. However, early-stage disease often presents with nonspecific or absent symptoms, and the lack of standardized screening for high-risk populations further impedes timely detection. Consequently, many patients are diagnosed at advanced stages, forfeiting the optimal surgical window. 4 Strengthening early diagnostic methods and interventions is thus critical to prolong survival and improve quality of life.
Alongside clinical challenges, recent evidence points to an intricate connection between gut microecology and the development of pancreatic cancer. The human gastrointestinal tract harbors approximately 100 trillion microorganisms, forming a complex microecological network essential for maintaining intestinal homeostasis, epithelial barrier integrity, and innate immunity.5,6 Disturbances in this network—characterized by a decline in beneficial bacteria and an increase in opportunistic pathogens—have been linked to pancreatic tumorigenesis. 4 Persistent immune responses and chronic inflammation triggered by dysbiosis can reshape the tumor microenvironment. Microbial metabolites entering the pancreas via the mesenteric veins may alter metabolic pathways, stimulate pro-inflammatory signals, and inhibit immune cell differentiation, thereby promoting tumor progression and metastasis.7,8 Although the precise mechanisms are not yet fully elucidated, existing evidence highlights the critical role of the gut microbiota in the pathogenesis and progression of pancreatic cancer (Figure 1).
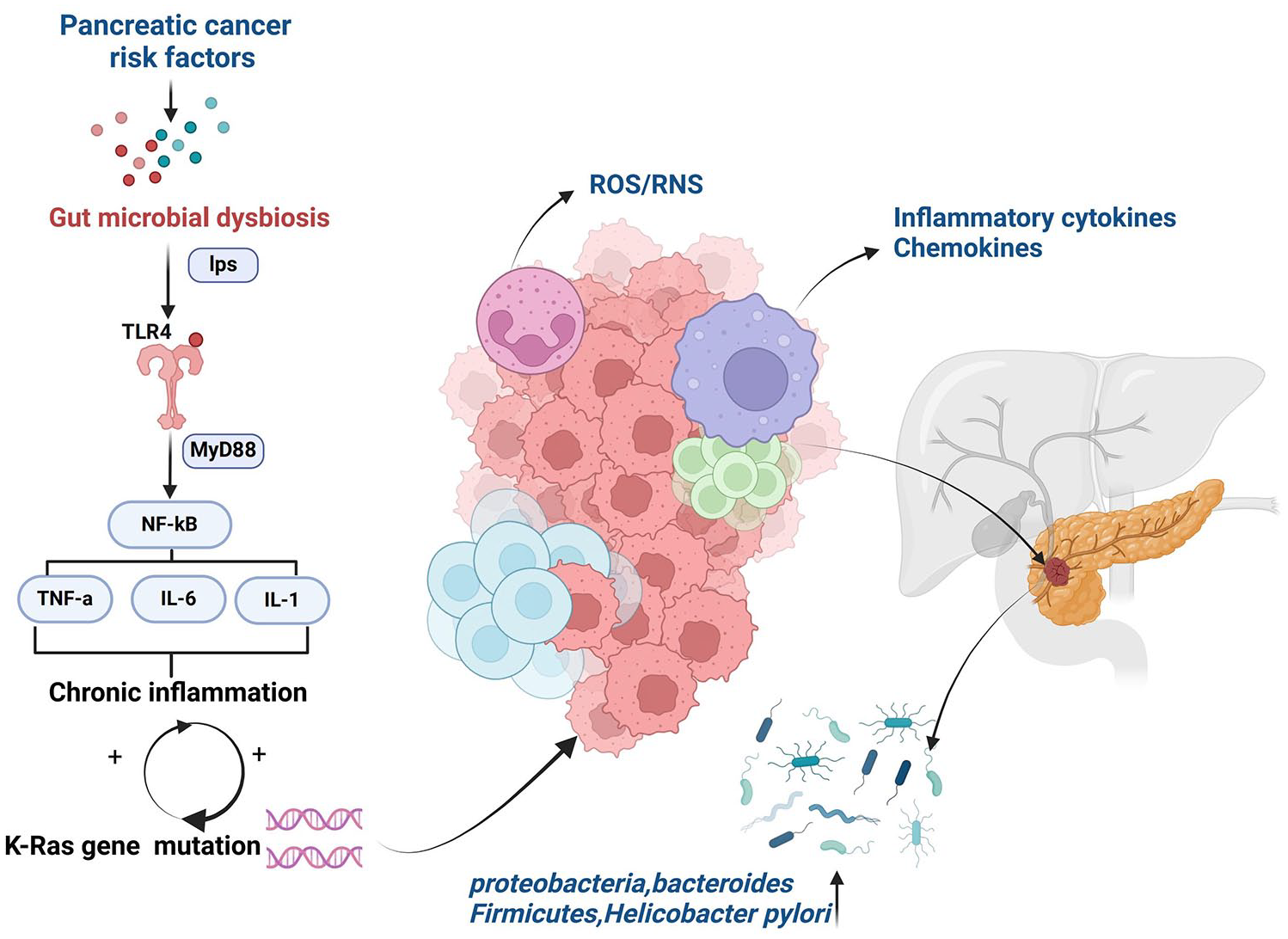
The influence of gut-microbiota-derived metabolites in the pathogenesis and progression of pancreatic cancer.
Taken together, these insights highlight the urgent need for enhanced early detection strategies and a deeper exploration of microbe-driven mechanisms, which may open new avenues for the prevention and treatment of pancreatic cancer.
The correlation between risk factors for pancreatic cancer and gut microbiota
Obesity
Obesity, elevated body mass index (BMI), and weight gain have been consistently linked to a heightened risk of pancreatic cancer. 9 Epidemiological evidence suggests that high-calorie, high-fat, and high-sugar diets significantly contribute to overweight or obesity, thereby increasing susceptibility to pancreatic malignancies. Notably, a BMI exceeding 35 constitutes a risk factor for both men and women. In addition, obesity and elevated BMI correlate with hyperglycemia, insulin resistance, increased circulating insulin and C-peptide, and diabetes—all of which are implicated in pancreatic carcinogenesis.
Although the precise mechanism underlying obesity-induced pancreatic cancer remains elusive, obesity is hallmarked by chronic inflammation and metabolic dysregulation. Increasing evidence points to gut microbiota dysbiosis as a crucial contributor to these metabolic disturbances. 10 Perturbations in the microbial community disrupt the normal host–microbe relationship, precipitating chronic inflammation and metabolic disorders, including obesity and its related pathologies. 11 Moreover, obesity is characterized by altered gut microbial diversity. The excessive release of lipopolysaccharide (LPS) from the gut microbiota in obesity often leads to metabolic endotoxemia. This low-grade chronic inflammation augments the secretion of proinflammatory cytokines and activates the nuclear factor-κB (NF-κB) pathway—collectively heightening the risk of pancreatic cancer.12,13
Obese individuals often exhibit elevated levels of pro-inflammatory adipokines (e.g., interleukin-6 and tumor necrosis factor-α), one principal pathway of which arises from the activation of innate immune responses.14 –16 Innate immunity relies on Toll-like receptors (TLRs), which recognize pathogen-associated molecular patterns such as lipopolysaccharides (LPS), viral CpG motifs, and zymogens. 17 In vitro studies indicate that LPS, originating from the gut, triggers TLR activation, subsequently driving the nuclear translocation of NF-κB. This process initiates the transcription of pro-inflammatory cytokines (e.g., IL-6, IL-1, and TNF-α) and culminates in both cytokine release and recruitment of pro-inflammatory mediators. 18 Sustained NF-κB activation can foster tumorigenesis by inhibiting apoptosis, promoting angiogenesis, enhancing invasiveness and metastasis of pancreatic cancer cells, and facilitating chemotherapy resistance. 19 These observations collectively underscore the critical role of obesity-associated inflammation and gut microbiota dysbiosis in promoting pancreatic cancer development and progression.
Diabetes
Type 2 diabetes (T2D) has emerged as a critical factor contributing to the rising incidence of pancreatic cancer. Approximately half of the patients diagnosed with pancreatic cancer already have pre-existing diabetes, with the majority of diabetes cases identified within 2–3 years prior to the detection of pancreatic malignancies.20,21 Both T2D and obesity share overlapping as well as distinct mechanisms that drive pancreatic tumorigenesis. For instance, T2D and obesity are often characterized by insulin resistance, hyperinsulinemia, and elevated insulin-like growth factor-1 (IGF-1) levels—all of which can promote oncogenic processes. Increased circulating insulin reduces insulin growth factor-binding proteins (IGFBP)-1 and -2, thereby raising bioavailable IGF-1 levels. Upon binding to its receptor (IGF-1R), IGF-1 can stimulate tumor cell proliferation, inhibit apoptosis, and enhance angiogenesis, collectively fueling cancer progression.22,23 Epidemiological data further indicate a positive correlation between diabetes and pancreatic cancer risk, with one study reporting a 22% increased risk for every additional 25 g/day of fructose consumption. 24
Beyond these metabolic factors, a growing body of literature supports a role for gut microbiota dysbiosis in diabetes, particularly in type 1 diabetes (T1D). Multiple studies document significant alterations in the gut microbial composition of T1D patients, including decreased Actinobacteria, Firmicutes, and an altered Firmicutes/Bacteroidetes ratio, alongside increased Bacteroidetes.25,26 At the genus level, Clostridium, Bacteroides, Lactobacillus, Bifidobacterium, Coccidia/Rectum group, and Prevotella are notably reduced in T1D children. 27 Similar trends of elevated Bacteroidetes have also been reported in T1D patients and individuals at high risk of T1D.25,28,29 By contrast, T2D patients generally exhibit moderate dysbiosis, with reduced butyrate-producing bacteria and an expansion of opportunistic pathogens, thereby heightening susceptibility to various diseases.30 –32 Some specific bacterial strains have been shown to modulate glycemic control in diabetic populations. 33 In addition to insulin resistance triggered by gut microbiota (GM) dysfunction, recent findings suggest that changes in microbial-derived metabolites—particularly acetate and butyrate—can further elevate pancreatic cancer risk. This effect may be mediated by chronic inflammation and endotoxemia, arising from impaired epithelial tight junctions in the intestinal mucosa, ultimately contributing to a tumor-favoring microenvironment.12,13
Probiotic interventions targeting GM composition may offer therapeutic benefits by mitigating low-grade inflammation associated with dysbiosis.34 –36 Certain probiotic strains can, for example, upregulate the anti-inflammatory cytokine IL-10, thereby suppressing pro-inflammatory mediators (e.g., IFN-γ, IL-2, and IL-1β) and potentially preventing the onset of diabetes.36,37 These findings underscore the intricate interplay among metabolic disorders, GM, and inflammatory pathways in the pathogenesis of pancreatic cancer.
Chronic pancreatitis
Chronic pancreatitis is a fibrosing inflammatory disease widely recognized as a significant risk factor for pancreatic cancer. Epidemiological studies indicate that patients with chronic pancreatitis face an approximately eightfold increase in pancreatic cancer incidence within 5 years of diagnosis. 38 The longer the duration of chronic pancreatitis, the higher the risk of disease onset. Given that GM dysbiosis is considered one of the potential drivers of chronic pancreatitis, its progression also contributes to the initiation and development of pancreatic cancer. Multiple investigations reveal reduced relative abundances of Enterococcus faecalis and Bromus rumen—particularly in individuals also presenting with diabetes—suggesting a notable decrease in GM diversity among patients with chronic pancreatitis.39,40 Overgrowth of gut microbes may foster a microenvironment conducive to pancreatic carcinogenesis; for example, small intestinal bacterial overgrowth can induce inflammation via lipopolysaccharide (LPS), thereby triggering K-RAS mutations—an effect especially pertinent in certain chronic pancreatitis patients. 41 Microbial infections can cause carcinogenic inflammation in the pancreas, whether locally or systemically, as the persistent inflammatory stimulation driven by these microbes is thought to initiate K-RAS activation. 42 In addition, murine models have demonstrated accelerated progression of pancreatic ductal adenocarcinoma in the presence of gastrointestinal microbiota, underscoring the potential influence of host gut flora on pancreatic disease. 42
Chronic pancreatitis, as well as other inflammatory disorders, is thus recognized as a potent risk factor for PDAC, with significantly higher incidence rates compared to unaffected populations. Convergent evidence from epidemiological data, preclinical research, and clinical studies consistently supports this association. Inflammatory responses play a pivotal role in pancreatic tumorigenesis: patients with hereditary autoimmune pancreatitis exhibit an approximate 40% lifetime risk of developing PDAC, whereas those with chronic pancreatitis have a 16-fold increased risk relative to controls.43,44 While several mechanistic pathways have been proposed to explain how GM contributes to pancreatic cancer, inflammation remains central to these processes. Animal studies show that germ-free mice display lower cancer incidence, and antibiotic-treated mice—characterized by reduced microbiota—also demonstrate lower susceptibility to tumor formation, likely due to diminished tumor-associated inflammation.
Moreover, the GM can regulate the host’s inflammatory response and influence an array of physiological functions. When dysregulated, these changes can lead to functional impairments and disease. Current evidence suggests that there is a correlation between metabolic abnormalities in chronic pancreatitis and disruptions in the GM; however, further research is needed to elucidate the precise nature of this relationship. Further mechanistic insights come from work by Daniluk et al., 45 who reported that inflammatory stimuli can elevate wild-type Ras activity to pathological levels. In mice expressing oncogenic K-Ras, persistent inflammatory stimuli exert prolonged effects on Ras, NF-κB, and Cox-2 signaling. Once oncogenic Ras is activated, inflammatory signals trigger a positive feedback loop involving NF-κB, thereby promoting the release of pro-inflammatory mediators and further amplifying Ras activity. 45 Under these conditions, mice expressing oncogenic K-Ras develop chronic pancreatic inflammation and pancreatic intraepithelial neoplasia, highlighting the critical role of inflammation in the progression to PDAC. 46
Diagnosis of pancreatic cancer using GM
The current diagnostic workflow for pancreatic cancer largely hinges on clinical manifestations, tumor markers, imaging examinations, and histopathological features. 47 However, the only FDA-approved biomarker for pancreatic ductal adenocarcinoma (PDAC), carbohydrate antigen 19-9 (CA19-9), exhibits limited sensitivity and specificity, restricting its utility to disease monitoring rather than population-wide screening. The lack of broadly applicable biomarkers or screening tools complicates early PDAC diagnosis. 48 Recent studies propose that a deeper understanding of the GM may facilitate the identification of novel biomarkers, an urgent need in PDAC detection.49,50
Microbiome analyses reveal that Firmicutes and Bacteroidetes predominate in both PDAC patients and healthy controls, whereas patients exhibit notably higher abundances of Proteobacteria, Synergistetes, and Euryarchaeota. Although Proteobacteria account for only about 8% of the GM in PDAC patients, they constitute nearly 50% of the microbial community in pancreatic tumor tissues. 42 Furthermore, Aykut et al. 51 demonstrated via ITS1 region amplification of the 18S rRNA gene that Ascomycota and Basidiomycota dominate the fungal communities in both the gut and tumor tissues of PDAC patients.
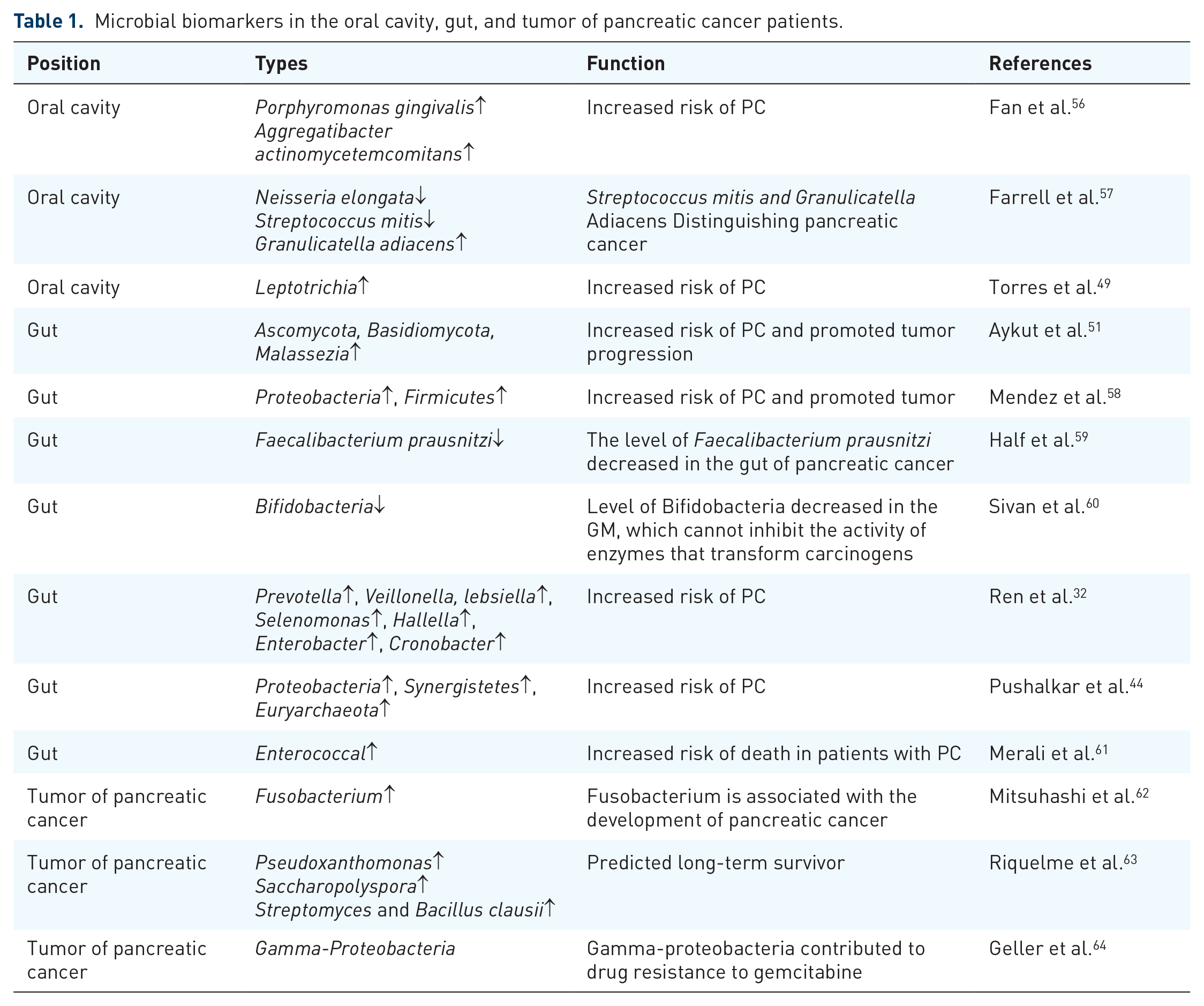
Saliva testing has also emerged as a potential noninvasive approach for PDAC detection, given that oral microbiota may migrate to the pancreas. 52 The presence of pathogenic oral bacteria can induce inflammatory responses, prompt the secretion of pro-inflammatory cytokines, promote oxidative stress, and cause DNA damage—collectively driving molecular alterations associated with tumorigenesis. Pancreatic inflammation and disturbances in GM may further enable the reflux of intestinal contents via the main pancreatic duct, introducing bacterial species into the pancreatic milieu and disrupting its normal microenvironment. 53 Notably, recent investigations employing 16S rRNA gene high-throughput sequencing underscore the potential of salivary RNA biomarkers in profiling oral microbiota.54,55 Compared with healthy individuals, PDAC patients exhibit elevated proportions of Treponema and reduced proportions of Porphyromonas in saliva, suggesting that these taxa may serve as novel biomarkers for pancreatic cancer diagnosis. A summary of the published data on enriched microbes at different sites in clinical studies of pancreatic cancer is included in Table 1. In this review, the unique microbiological characteristics of pancreatic cancer are examined from three perspectives: the oral microbiome, the intratumoral microbiome, and the metabolites produced by these microbial communities.
Microbial biomarkers in the oral cavity, gut, and tumor of pancreatic cancer patients.
Research on the oral microbiota and pancreatic cancer
Poor oral health has been associated with the risk of a number of medical conditions including atherosclerosis, diabetes, autoimmune conditions, obesity and digestive cancers, and lifestyle factors including smoking, alcohol consumption, and diet.Interestingly, several of these factors also are known or suspected risk factors for pancreatic cancer. 65 Numerous epidemiological studies and meta-analyses have evaluated specific oral health indicators—such as periodontal disease, tooth loss, and gingivitis—and their association with pancreatic cancer risk. Although these cohort studies vary in their methodologies and endpoints, their collective findings consistently indicate that poor oral health is linked to a higher risk of pancreatic cancer. 66
Since the initiation of the NIH Human Microbiome Project, research into the role of the host microbiome in cancer risk and prognosis has significantly expanded. This includes studies on the oral microbiome, which comprises over 700 bacterial species, in cancers associated with periodontal disease such as pancreatic cancer. 65 Until recently, the potential systemic effects of oral bacteria were not well understood. While oral bacteria had been primarily linked to cancers of the oral cavity, their potential association with cancers of more distant organs, particularly those within the digestive system, had only been hypothesized.
A study published in 2017 examined the oral microbiomes of 40 patients with newly diagnosed pancreatic cancer, 39 patients with intraductal papillary mucinous neoplasms (IPMNs), and 58 control individuals. Compared with the control group, pancreatic cancer patients exhibited higher levels of Firmicutes and several related taxa. However, after adjustment for multiple testing, the significance remained only at the phylum level. By contrast, control individuals had higher levels of Proteobacteria and associated taxa, including Gammaproteobacteria, Pasteurellales, Pasteurellaceae, Haemophilus, Haemophilus parainfluenzae, Betaproteobacteria, Neisseriales, Neisseriaceae, Neisseria, and Neisseria flavescens. 67
The epidemiological evidence supporting an association between periodontal disease and the risk of pancreatic cancer is compelling. These risk factors adversely affect the oral microbiome, leading to dysbiosis and an increased proportion of known pathogenic bacteria. Further investigation into the mechanisms by which periodontal disease contributes to the development of pancreatic cancer is necessary. In addition, the treatment of periodontal disease may have a beneficial effect on the prevention of pancreatic cancer.
The pancreatic tumor microbiome
Specific microbial communities within and surrounding pancreatic tumor tissues have been associated with tumorigenesis and progression. Fusobacterium, an opportunistic pathogen commonly colonizing the oral cavity and gut, has been detected in 8.8% of pancreatic cancer tissue samples.52,62,68 This suggests that oral microbiota may migrate to the digestive tract and pancreatic tissues through various transmission pathways. In pancreatic cystic tumors, bacterial DNA analysis revealed a significant enrichment of Fusobacterium.
During the tumorigenesis of pancreatic ductal adenocarcinoma, there is a successive alteration in microbial species across different bodily sites. Bacteria such as Porphyromonas gingivalis, Treponema denticola, P. stomatis, P. acidifaciens, P. endodontalis, and Filifactor alocis sequentially increase in pancreatic tissue as PDAC progresses. Conversely, probiotic-associated bacteria like Bifidobacterium breve and Lactiplantibacillus plantarum decline. In addition, the escalating trends of P. stomatis and P. acidifaciens observed in pancreatic tissue during PDAC tumorigenesis are mirrored in duodenal fluid and saliva. 69
Furthermore, microbial community compositions in pancreatic tissues differ between long-term survivors and short-term survivors of pancreatic cancer. P. gingivalis, P. endodontalis, and F. alocis progressively increase from chronic pancreatitis to stage IV PDAC in pancreatic tissues and are significantly enriched in the saliva of short-term survivors compared to long-term survivors. 69
The duodenal region of pancreatic cancer patients also exhibits distinct microbial communities compared to healthy controls. In the duodenal mucosa of pancreatic cancer patients, there is a higher abundance of Acinetobacter, Aquabacterium, Oceanobacillus, Rahnella, Massilia, Delftia, Deinococcus, and Sphingobium. By contrast, the duodenal microbiota of healthy individuals is enriched with Porphyromonas, Paenibacillus, Enhydrobacter, Escherichia, Shigella, and Pseudomonas. 70
These findings collectively indicate that the pancreatic tumor microbiome undergoes significant changes during PDAC development, with specific bacteria potentially playing roles in tumor progression, prognosis, and patient survival. Understanding these microbial dynamics is crucial for elucidating the mechanisms by which the microbiome influences pancreatic cancer and may offer novel avenues for prevention and therapy.
Impact of gut microbiota-derived metabolites on pancreatic tumors
The GM produces a variety of metabolites, some of which significantly influence tumorigenesis and tumor progression. Short-chain fatty acids (SCFAs) such as acetate, propionate, butyrate, and lactate can alter the pH within the colon and modulate the function of the gut immune system. These changes subsequently affect gene expression and cellular regulatory processes. 71
A reduction in propionate levels has been shown to decrease the abundance of mucosa-associated invariant T (MAIT) cells and regulatory T (Treg) cells, which are crucial for maintaining the integrity of the gut mucosal barrier. 72 Butyrate, another SCFA, interferes with histone modification and transcriptional regulation, impacting gene expression patterns related to cell proliferation and apoptosis. 73 Ren et al. 32 demonstrated that a decrease in butyrate-producing bacteria is closely linked to microbial dysbiosis in pancreatic cancer.
Bacteria and their secreted products within the tumor microenvironment exert profound effects on the development, treatment, and prognosis of pancreatic cancer. Currently recognized mechanisms by which the tumor microenvironment influences tumor growth include bile acid metabolism, immune modulation, and muscarinic receptor pathways. GM can translocate to pancreatic tumors, where they collaborate with intratumoral microbes to induce an immunosuppressive tumor microenvironment. This environment facilitates tumor growth and metastasis while limiting the efficacy of immune checkpoint inhibitors. For instance, Malassezia species are enriched in human pancreatic cancer tissues and can activate mannose-binding lectin (MBL), triggering the complement cascade and inflammatory immune responses. 51 The carbohydrate structures produced by Malassezia bind to MBL, leading to the activation of the C3 protein and subsequent complement-mediated inflammation. Complement activation stimulates cellular proliferation and migration, ultimately promoting tumor growth. 51
In addition, gut bacterial metabolites positively influence cancer therapy. Studies have reported that indole-3-acetic acid (3-IAA), a tryptophan metabolite produced by Bacteroides fragilis and Bacteroides polymorphus, is expressed at higher levels in pancreatic ductal adenocarcinoma patients who respond well to chemotherapy. 3-IAA is transported via the bloodstream to distal tumors, where it stimulates neutrophils to enhance the efficacy of chemotherapy, thereby aiding in cancer control. 74 Future therapeutic strategies could involve the direct supplementation of 3-IAA or adopting a high-tryptophan diet to bolster the effectiveness of chemotherapy in pancreatic cancer patients.
Given the vast diversity and complexity of the GM, the roles of its metabolites in tumorigenesis and tumor progression warrant further investigation. Understanding these interactions in greater detail may unveil novel therapeutic targets and strategies for the prevention and treatment of pancreatic cancer.
GM in pancreatic cancer treatment
The impact of GM and their metabolic products on tumor therapy has also garnered increasing attention. Sethi et al. investigated how oral antibiotic-induced alterations of the GM influence tumor development, revealing antitumor effects mediated by immune modulation—specifically, the promotion of Th1 cells secreting interferon-gamma (IFN-γ). Notably, these effects were not observed in animal models lacking mature T and B lymphocytes. Currently, in pancreatic cancer, gemcitabine-based chemotherapy remains the primary treatment modality; however, studies indicate that the microbiota significantly influences chemotherapy efficacy. 75 Geller et al. analyzed bacterial DNA in tumor samples from patients with pancreatic ductal adenocarcinoma (PDAC) and found that 76% of PDAC tumors contained detectable DNA of γ-Proteobacteria. In a colorectal cancer mouse model, these bacteria produced cytidine deaminase, which metabolized gemcitabine into its inactive form, thereby diminishing drug sensitivity. These findings suggest that combining antibiotics with gemcitabine-based chemotherapy may enhance therapeutic effectiveness 68 and provide an important adjunct to existing treatment strategies.
Fecal microbiota transplantation
Fecal microbiota transplantation (FMT) involves the transfer of healthy donor microbiota to replace disease-associated microbial communities, thereby restoring intestinal homeostasis. 76 Emerging evidence suggests that transplanting stool samples from particular patient groups into tumor-bearing mice—especially those receiving immune checkpoint inhibitors or antibiotic therapy—can augment immunotherapeutic efficacy. 77 Consequently, FMT represents a promising therapeutic avenue for treating both intestinal and extraintestinal pathologies by re-establishing a balanced gut microecology.
Recent studies provide further support for FMT’s potential in pancreatic cancer. In one investigation, fecal samples derived from healthy controls (HC), short-term survival patients (STS), and long-term survival patients (LTS) were transplanted into tumor-bearing mice, simulating a clinical FMT scenario. Notably, tumors in mice receiving LTS stool exhibited significantly reduced growth compared with those receiving STS or HC stool, suggesting that the microbiota of long-term surviving patients with pancreatic ductal adenocarcinoma (PDAC) harbor tumor-suppressive properties. By contrast, mice receiving STS donor stool developed larger tumors than those that received HC donor stool, implying that gut/tumor-associated microbes from PDAC patients may promote tumor progression. 63 These findings highlight FMT’s capability to modulate the GM, potentially preventing or slowing pancreatic tumor growth.
Despite these encouraging outcomes, the precise mechanisms underlying FMT’s clinical efficacy require further elucidation, necessitating large-scale prospective clinical trials to evaluate its safety and therapeutic potential. Moving forward, FMT may emerge as a valuable adjunct or standalone strategy for managing pancreatic cancer.
Immunotherapy
Immunotherapy harnesses and amplifies the host’s immune response to tumors, thereby inhibiting tumor growth and metastasis while preserving the functional integrity of normal tissues. 78 Emerging evidence highlights the pivotal role of GM in shaping the human immune system, including the modulation of immune checkpoints that can augment the efficacy of tumor immunotherapy.79 –81 Notably, Pushalkar et al. detected distinct gut- and tumor-associated microbial communities in a pancreatic cancer mouse model, suggesting that translocation of specific bacterial taxa from the intestine into the tumor microenvironment may occur. In an orthotopic pancreatic cancer mouse model, depletion of these bacteria using antibiotics effectively remodeled the tumor microenvironment, activated T cells, improved immune surveillance, and enhanced susceptibility to immunotherapy. 42 These findings underscore the potential of manipulating GM to improve immunotherapeutic outcomes in pancreatic cancer.
Further studies bolster this concept. Oral administration of Bifidobacterium has demonstrated antitumor efficacy comparable to anti-PD-L1 antibody therapy, and combining both interventions significantly reduced tumor volume. 60 The underlying mechanism appears to involve enhanced dendritic cell function, culminating in the activation and accumulation of CD8+ T cells within the tumor microenvironment. Collectively, these observations suggest that targeted microbiota modulation could serve as a vital adjunct to bolster cancer immunotherapy. 4
Probiotics and their derivatives
Probiotics, such as Bifidobacterium and Lactobacillus, are beneficial microorganisms naturally present in the gut that help maintain microbial balance and resist the detrimental effects of stomach acid and bile salts. Oral administration of probiotics has been reported to exert antitumor effects and enhance the efficacy of chemotherapy. For instance, Chen et al. observed that Lactobacillus supplementation improved gemcitabine efficacy in a murine model of pancreatic cancer, thereby increasing treatment tolerance. In addition, iron chelate derivatives of Lactobacillus casei can regulate the cell cycle via p53 activation, exhibiting antitumor potential in refractory and 5-fluorouracil-resistant pancreatic cancers.82,83
Meanwhile, commensal Bifidobacterium has emerged as a key regulator of gastrointestinal health. Oral administration of Bifidobacterium alone can attenuate tumor growth by enhancing antitumor immunity, preventing pathogenic colonization, and lowering local pH to maintain microbial homeostasis. 60 Diets rich in probiotics have likewise been correlated with a reduced incidence of pancreatitis, potentially lowering the risk of pancreatic cancer.84,85 Moreover, Bifidobacterium ferments glucose to produce SCFAs, including acetate, butyrate, and lactate, all of which confer notable benefits to gut health. Among these SCFAs, butyrate is particularly relevant to tumorigenesis and progression, although its role in pancreatic cancer remains less explored relative to lung cancer, colorectal cancer, and leukemia.86,87
Evidence suggests that sodium butyrate inhibits pancreatic cancer cell invasion and metastasis by downregulating integrin β4 expression.85,88 It further promotes tumor cell differentiation and induces the expression of certain tumor-associated antigens. 89 Mechanistically, butyrate may exert antineoplastic effects by inhibiting histone deacetylase (HDAC) and telomerase activity. 90 In addition, it induces Treg cell proliferation and the production of anti-inflammatory cytokines, reduces DNA oxidative damage, promotes apoptosis in damaged cells, and halts tumor cell proliferation. 91 Taken together, these findings underscore butyrate’s broad inhibitory impact on pancreatic tumor initiation, progression, and metastasis. Consequently, strategies aimed at modulating butyrate levels via GM interventions may hold promise for the prevention and treatment of pancreatic cancer.
In the future, integrating GM modulation with immunotherapy may represent a promising strategy for pancreatic cancer treatment. This approach includes using antibiotics to mitigate immunosuppressive effects and target specific drug-resistant bacterial strains, as well as employing FMT or tailored probiotics to enhance immunotherapeutic efficacy. Tumor-local immune responses and the GM undergo coordinated alterations as the tumor progresses. 92 Because immunological characteristics differ across tumor stages, pancreatic cancer immunotherapy regimens should be tailored accordingly. Early immunotherapy intervention combined with GM modulation holds the potential to achieve more favorable outcomes.
Impact of gut microbial diversity on prognosis in pancreatic cancer patients
Most patients with pancreatic cancer present at advanced stages, leading to a generally poor prognosis. Even among those who undergo surgical resection, the recurrence rate remains high, and the median survival is only 24 to 30 months.64,93,94 The factors influencing LTS in this population remain largely unknown. Notably, a cohort study by Riquelme et al. 95 revealed that GM may critically affect survival outcomes in pancreatic cancer. LTS exhibited significantly higher α-diversity in their tumor microbiota compared with STS, and patients with elevated α-diversity displayed a markedly improved overall survival relative to those with low α-diversity. Furthermore, while microbial diversity often correlates with favorable therapeutic responses, dysbiosis has been linked to poor treatment outcomes, chronic disease progression, and carcinogenesis. 80
In two independent cohorts of patients undergoing pancreatic cancer resection, systematic analyses of tumor microbiota diversity and composition similarly showed greater bacterial diversity among LTS. Specifically, the presence of Pseudomonas, Saccharopolyspora, and Streptomyces within the tumor microenvironment was predictive of extended survival. Complementary murine experiments indicated that alterations in GM could influence tumor microbiota and tumor growth. These findings suggest that bacterial colonization of pancreatic tumors by gut-derived species can reshape the intratumoral microbial landscape. The diversity and composition of the pancreatic cancer microbiome also appear to modulate immune infiltration, thereby affecting patient survival. In favorable prognostic groups, activation and recruitment of CD8+ T cells into the tumor environment are commonly observed. This immunologic phenotype may be shaped by a unique intratumoral microbiota, which, in turn, fosters a tumor microenvironment conducive to improved outcomes. Consequently, specific microbial signatures may serve as novel prognostic biomarkers for pancreatic cancer.96,97
This review is primarily based on published literature and lacks original data and clinical research supplementation. The precise mechanisms of GM in pancreatic cancer require further elucidation, as most existing studies remain at the basic research level. The feasibility of clinical applications warrants additional investigation. Although the potential of GM in treatment is discussed, its actual efficacy and safety must be validated through large-scale clinical trials.
Future research should clarify the mechanisms by which GM affects pancreatic cancer, especially through human studies that bridge the gap between basic science and clinical application. Current findings rely heavily on preclinical models, and standardized protocols are needed to ensure the consistency and safety of therapies like FMT and probiotics. In addition, personalized treatment strategies based on individual microbiota profiles should be developed to improve therapeutic outcomes. Large-scale clinical trials are essential to validate the efficacy and safety of these microbiota-based interventions in pancreatic cancer.
Conclusion
Pancreatic cancer is a highly malignant digestive system tumor, and there are currently no effective early diagnostic and treatment methods. GM plays a crucial role in the occurrence and development of pancreatic cancer, involving inflammatory states and immune responses. This review elucidates the impact of GM on pancreatic cancer and predicts its disease risk and survival status through microbiota analysis, proposing a new approach for the diagnosis and survival prediction of pancreatic cancer. FMT, probiotics, and immunotherapy represent new ideas for the diagnosis and treatment of pancreatic cancer. In the future, it is expected that changes in the microbiota of the oral cavity and gastrointestinal tract can be detected for the diagnosis of pancreatic cancer, and microbes can be utilized for immunotherapy to improve the cure rate and survival rate of patients with pancreatic cancer.
Footnotes
Acknowledgements
We would like to extend our gratitude to our colleagues and peers for their valuable discussions and feedback that contributed to the development of this review. We also appreciate the institutional support provided throughout the research process.