
Abstract
Defensins are critical components of the innate immune system and play an important role in the integration of innate and adaptive immune responses. Although information on the immunomodulatory properties of peptidoglycan from bacteria is abundant, little is known about the β-defensin induction effect of peptidoglycan from the probiotic Lactobacillus. This study investigated the effect of intact peptidoglycan from L. rhamnosus MLGA on the induction of avian β-defensin 9 in chicken immune cells and intestinal explants. Peptidoglycan from Lactobacillus rhamnosus MLGA dose dependently promoted avian β-defensin 9 mRNA expression in chicken PBMCs, splenocytes, thymocytes, hepatocytes, and chicken embryo jejunum, ileum, and cecum explants and increased the capacity of PBMC or splenocyte lysates to inhibit the growth of Salmonella Enteritidis. In contrast to the effect of L. rhamnosus MLGA-derived peptidoglycan, peptidoglycan derived from pathogenic Staphylococcus aureus reduced avian β-defensin 9 mRNA expression in chicken PBMCs and splenocytes. The inducible effect of peptidoglycan from L. rhamnosus MLGA on avian β-defensin 9 expression in PBMCs and splenocytes was observed without activation of the expression of associated pro-inflammatory cytokines IL-1β, IL-8, and IL-12p40, whereas these cytokine expressions were suppressed by peptidoglycan hydrolysate obtained by lysozyme digestion. The results of the present study show the capability of peptidoglycan derived from L. rhamnosus MLGA to induce the antimicrobial peptide defensin while simultaneously avoiding the deleterious risks of an inflammatory response.
Introduction
Defensins are small cationic antimicrobial peptides with broad-spectrum direct microbicidal activities against bacteria, protozoa, enveloped virus, and fungi. 1 Defensins are recognized as important effector molecules of host innate immunity and are widely distributed in various tissues, especially in immune organs, the epithelium of skin, and throughout the digestive, respiratory, and urogenital tracts. 1 , 2 In addition to their direct microbicidal activity, defensins also show multiple biological activities, including anti-tumor and anti-inflammatory properties, neutralizing of endotoxins, wound healing, and chemoattracting lymphocytes, dendritic cells, and monocytes, and thus play an important role in the integration of the innate and adaptive immune responses against infections. 3 , 4 The defensin family of vertebrate animals can be divided into α-, β-, and θ-defensin subgroups based on the spacing pattern of cysteines, which form three conserved disulfide bridges. 5 Interestingly, only β-defensins (avian β-defensins, AvBDs) have been reported to exist in avian species so far. 2 The chicken genome encodes a total of 14 AvBDs (AvBD1–14), which are densely clustered on chicken chromosome 3q and expressed in a wide range of tissues. 6 , 7
Among the 14 AvBDs, AvBD9 is extensively expressed in various organs and tissues of chicken.6–8 AvBD9 displays broad-spectrum antibacterial activities against Clostridium perfringens, Staphylococcus aureus, Campylobacter jejuni, Enterobacter cloacae, Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Shigella sonnei, and shows a prominent fungicidal activity against both unicellular and multicellular fungi.8–10 In addition to the potent antimicrobial activity of AvBD9, this peptide exhibits low hemolytic activity and low toxicity against animal cells, 8 and has multiple functions in the host defense against infection, 11 suggesting that AvBD9 plays an important role in both innate and adaptive immunity. Several studies revealed that some dietary compounds, including vitamin D3 and short-chain fatty acids in particular, potently promote AvBD9 expression in different immune cell types and the intestines of chicken, and thus boost host immunity and disease resistance in chicken.11–13 In contrast, tissue-specific reduction of AvBD9 expression was observed in chickens in response to several microbial infections such as Salmonella enterica, infectious bronchitis virus, and Marek’s disease virus infection,14–16 indicating that suppressing AvBD9 expression may be a major immune evasion strategy used by pathogens. Therefore, dietary modulation of the synthesis of endogenous antimicrobial peptides such as AvBD9 may have the potential to be developed as a cost-effective, antibiotic-alternative approach to disease control and prevention for poultry production.
Peptidoglycan, a major component of Gram-positive bacterial cell walls (accounting for approximately 90% of their dry mass), plays crucial roles in bacterial growth and survival as well as in the modulation of host immune responses. 17 Peptidoglycan from probiotic lactobacilli has been shown to possess multiple biological activities including immunomodulatory,18–21 anti-tumor,22–24 and anti-infection, 25 among which the immunomodulatory properties of peptidoglycan have been especially well documented. It has been demonstrated that peptidoglycan from lactobacilli can regulate not only innate immunity but also adaptive immune responses. 26 , 27
Several studies have shown that specific probiotic Lactobacillus strains strongly induced mRNA expression and protein production of β-defensin in epithelial cells, thus enhancing host defense against infection.28–31 We previously reported that heat-killed Lactobacillus rhamnosus MLGA that was isolated from health chicken intestine enhances AvBD9 gene expression in primary intestinal epithelial cells of chicken embryos, 32 implying that specific heat-stable cellular components from this probiotic bacterium contribute to this beneficial effect. Although the immunoregulatory activity of peptidoglycan from Lactobacillus has been studied extensively, little is known about the β-defensin induction effect of peptidoglycan. Given peptidoglycan is the major component of the cell walls of Lactobacillus and there is similar immunological and biological activity between defensin and peptidoglycan, we hypothesized that peptidoglycan from specific probiotic Lactobacillus strains can stimulate defensin expression and thus improve immune defense against pathogens. Therefore, the objective of the present study was to investigate the effect of intact peptidoglycan derived from L. rhamnosus MLGA on the expression of a representative chicken β-defensin (AvBD9) in jejunum, ileum, and cecum explants as well as immune cells. In addition, pro-inflammatory cytokine expression in immune cells in response to peptidoglycan stimulation was determined to examine whether peptidoglycan derived from L. rhamnosus MLGA had an impact on triggering inflammatory response while stimulating AvBD9 expression.
Material and methods
Bacterial strains
The probiotic L. rhamnosus MLGA strain was isolated from the small intestine of a healthy chicken 32 and preserved in a glycerin tube at –80°C. L. rhamnosus MLGA was cultured statically in Mann-Rogosa-Sharpe broth (Solarbio, Beijing, China) overnight at 37°C under anaerobic conditions. The bacteria were grown until log phase and then harvested by centrifugation at 2200 g for 20 min and washed three times with sterile 0.9% saline solution (w/v = 1:10). The collected bacteria were used for peptidoglycan preparation. Salmonella Enteritidis (ATCC 13076) was cultured in Tryptic Soy Broth (TSB) medium (Baisi Biotechnology, Hangzhou, China) at 37°C for 8 h. This was followed by three passages every 8 h into fresh TSB for a total of 24 h, to ensure log phase growth. Bacterial cells were washed three times in sterile PBS, pH 7.4, by centrifugation at 1500 g for 15 min, quantified with a spectrophotometer at 625 nm using an established concentration curve, and diluted in sterile PBS as per required concentrations for the experiment. The concentration of S. Enteritidis was confirmed by plating on BGA plates.
Preparation of intact peptidoglycan from L. rhamnosus MLGA
Intact peptidoglycan was extracted from L. rhamnosus MLGA based on a trichloroacetic acid (TCA) method as described previously 33 with minor modifications. Briefly, the bacteria were dissolved in 10% TCA (w/v = 1:10) and incubated in a boiling bath for 1 h. The mixture was immediately cooled and then centrifuged at 13,000 g for 10 min. The sediment was washed with distilled water to remove TCA, then treated with a special solvent composed of acetic acid-sodium acetate buffer (0.5 M acetic acid and 0.2 M sodium acetate, pH 4.5), chloroform and methanol at a ratio of 4:5:10 (v/v/v) for 24 h. After centrifugation at 4000 g for 20 min, the insoluble resides were incubated in Tris-HCl (0.1 M, pH 7.5) containing 3500 U trypsin at 37°C in a shaking bath (140 rpm) for 12 h. The mixture was then centrifuged at 8000 g for 20 min. Finally, the sediment obtained was washed with sterile distilled water 4–5 times, lyophilized, and stored at –80°C for further use. The target product was analyzed and confirmed as intact peptidoglycan by means of transmission electron microscopy, amino acid composition analysis, and lysozyme digestion test (data not shown).
Preparation of L. rhamnosus MLGA-derived peptidoglycan hydrolysate by lysozyme
Lyophilized intact peptidoglycan extracted from L. rhamnosus MLGA was suspended in PBS (pH 6.8) to obtain a 10% (w/v) peptidoglycan solution. Chicken egg white lysozyme (Amresco Inc., Ohio, USA) was then added to the solution at a ratio of 1:100 (enzyme/peptidoglycan), followed by incubation for 48 h at 37°C with gentle shaking. After digestion, the reaction solution was heated in a boiling-water bath for 15 min to inactivate the lysozyme and then lyophilized.
Isolation, culture, and stimulation of immune cells and intestinal tissue explants
PBMCs, splenocytes, thymocytes, and hepatocytes were isolated from the anti-coagulated blood, spleens, thymuses, and livers of 3- to 4-wk-old Arbor Acres broiler chickens. Blood was collected in tubes containing heparin sodium from the wing vein. PBMCs were isolated from anti-coagulated blood through gradient centrifugation using Histopaque® 1077 (Sigma-Aldrich, St. Louis, MO, USA) following the manufacturer's procedure. Chickens were killed by cervical dislocation and soaked in 75% ethanol for 3 min. The spleens, livers, and thymuses were harvested aseptically, and rinsed in D-Hank’s balanced salt solution (HBSS) (Solarbio, Beijing, China) supplemented with 200 U/ml of penicillin (Solarbio) and 200 µg/ml of streptomycin (Solarbio). Spleens and thymuses were gently disrupted with the flat end of a 10 ml syringe plunger and passed through a cell strainer to obtain single-cell suspensions in HBSS. Splenocytes and thymocytes were collected by a density gradient centrifugation using Histopaque® 1077. Cells were washed three times with HBSS by centrifugation at 500 g at 4°C for 10 min and then re-suspended in complete RPMI 1640 medium containing 10% FBS, 50 mM of
All animal procedures reported herein were in line with the Laboratory Animal Welfare and Ethics Censorship and approved by the Laboratory Animal Ethics Committee of Jiangxi Agricultural University (Nanchang, Jiangxi, China).
Real-time quantitative RT-PCR
Total RNA was extracted from cells or gut explants using Trizol Reagent (TransGen Biotech, Beijing, China) following the manufacturer’s instructions. RNA purity was assessed by examining the absorbance ratio at 260 and 280 nm using a NanoDrop ND-100 microvolume spectrophotometer (Thermo Fisher Scientific), whereas integrity was verified by electrophoresis on 1% denaturing agarose gel. Reverse transcription was performed using an M-MLV Reverse Transcriptase Kit (ShineGene Bioetch, Shanghai, China) following the manufacturer’s instructions. Real-time RT-PCR was then conducted on FTC2000 Real-Time PCR Detection System (Funglyn Biotech Inc., Canada) in 50 µl reaction volumes containing Hotstart Fluo-PCR mix (ShineGene), primers, fluorescent probes (TaKaRa Biotechnology, Dalian, China), and cDNA. PCR cycling conditions were 94°C for 4 min, followed by 40 cycles of 94°C for 30 s, and 60°C for 1 min. The specificity of PCR reaction was confirmed by the melt curve analysis. The primers and probes used in quantitative real-time PCR reactions are shown in Table 1. The relative gene expression levels were quantified using the comparative 2–ΔΔCt method with the GAPDH gene as a reference for normalization. Results were presented as fold-expression change relative to the control group (medium alone). All the assays were performed in triplicate and repeated at least two times.
The primer and probe sequences of AvBD9 and cytokines for real-time PCR reactions.

AvBD9: avian β-defensin 9.
Antibacterial capacity of PBMCs and splenocytes treated with peptidoglycan from L. rhamnosus MLGA
The antibacterial activity of cell lysates was measured as previously described with slight modifications. 11 Briefly, PBMCs and splenocytes were cultured in antibiotic-free RPMI 1640 medium containing 0, 5, 10, 50, or 100 μg/ml of peptidoglycan from L. rhamnosus MLGA for 6 h at 37°C and 5% CO2. Cell culture supernatants were collected, and cells were then scraped, lysed with 1% Triton X-100, and centrifuged 12,000 g for 10 min at 4°C. Serial two-fold dilutions were then prepared from the cell lysate supernatants. Cell culture supernatants and cell lysate dilutions were incubated with 1 × 104 CFU of S. Enteritidis ATCC13076 in TSB medium for 9 h in a 96-well plate at 37°C. Bacterial turbidity was measured at OD590 nm using a Microplate reader (Multiskan MK3, Thermo Fisher Scientific, Shanghai, China). Different concentrations of peptidoglycan from L. rhamnosus MLGA were also directly incubated with S. Enteritidis in TSB medium and the OD590 nm was recorded after 9 h incubation.
Statistical analysis
Data were expressed as mean ± standard deviation (SD) from two to three independent experiments. Data were analyzed using one-way analysis of variance and unpaired two-tailed Student’s t-test was used to evaluate the statistical significance between treatments using SPSS 17.0 software. Differences were considered statistically significant when the P value was < 0.05.
Results
Induction of AvBD9 gene expression in primary chicken immune cells and intestinal tissue explants by peptidoglycan from L. rhamnosus MLGA
In a previous study we demonstrated that intact peptidoglycan from L. rhamnosus MLGA induces AvBD9 gene expression in primary chicken small intestinal epithelial cells in both time- and dose-dependent manners, with the maximal response occurring at 4 h post-stimulation at a concentration of 50 µg/ml. 34 To further examine whether L. rhamnosus MLGA-derived peptidoglycan is capable of stimulating AvBD9 expression in other cell types, especially immune cells, we performed both dose-response and time-course experiments in primary PBMCs, splenocytes, thymocytes, and hepatocytes. An obvious time-dependent increase in AvBD9 expression was elicited in PBMCs in response to treatment of 50 µg/ml peptidoglycan, peaking at 6 h post-stimulation and returning to the basal level after 48 h of incubation (Figure 1a). A maximal response of AvBD9 expression was also observed in splenocytes and hepatocytes following 6 h stimulation, although the magnitude of induction was much less than in PBMCs (Figure 1a). However, no time-dependent induction of AvBD9 was seen in thymocytes on stimulation with 50 µg/ml peptidoglycan (Figure 1a), suggestive of cell-specific regulation of AvBD9 expression by peptidoglycan from L. rhamnosus MLGA. The expression of AvBD9 mRNA was dose-dependently augmented in all tested cells upon peptidoglycan treatment for 6 h, and the induction of AvBD9 expression was more prominent in the PBMCs than in the other three cell types (Figure 1b). Similarly, L. rhamnosus MLGA-derived peptidoglycan treatment exhibited a dose-dependent induction of AvBD9 in jejunum, ileum, and cecum explants after 24 h stimulation, with a maximal effect seen at the concentration of 10 µg/ml in the jejunum and ileum explants, followed by a gradual decline as the peptidoglycan concentration was increased (Figure 1c).
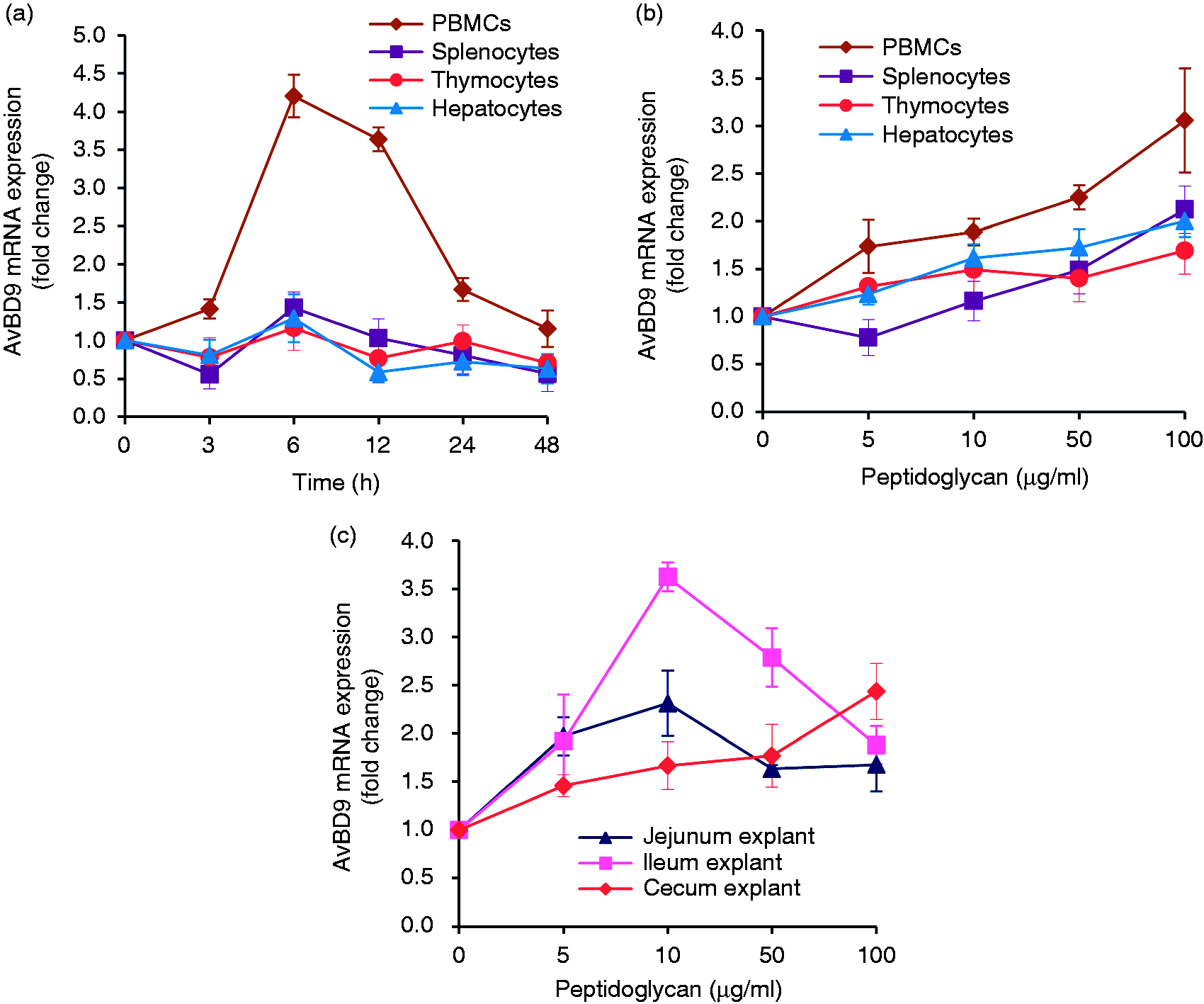
Intact peptidoglycan from L. rhamnosus MLGA induces avian β-defensin 9 (AvBD9) gene expression in different chicken cell types and intestinal tissue explants. Chicken primary cells were incubated with 50 µg/ml of peptidoglycan for indicated time points (a) or indicated concentrations of peptidoglycan for 6 h (b). (c) Chicken jejunal, ileal, and cecal explants were treated with indicated concentrations of peptidoglycan for 24 h. The AvBD9 mRNA expression was analyzed by real-time RT-PCR, and the relative gene expression was calculated using the comparative 2-ΔΔCt method with the GAPDH gene as a reference for normalization and presented as fold-expression change relative to the control group. Values are given as the mean ± SD of two to three independent experiments.
Peptidoglycan can be hydrolyzed by lysozyme (also named N-acetylmuramide glycanohydrolase or muramidase), which is found in monocytes, macrophages, epithelial secretions, and intestinal Paneth cells. 35 Furthermore, the beneficial immunomodulatory effects of peptidoglycan from probiotics can be diminished by treatment with N-acetylmuramidase in some cases. 23 We next examined the modulatory effect of peptidoglycan fragments released by lysozyme on AvBD9 expression. L. rhamnosus MLGA peptidoglycan hydrolysate at 100 µg/ml increased AvBD9 mRNA expression in PBMCs (Figure 2a) but not in splenocytes (Figure 2b), albeit at a lesser magnitude than intact peptidoglycan.
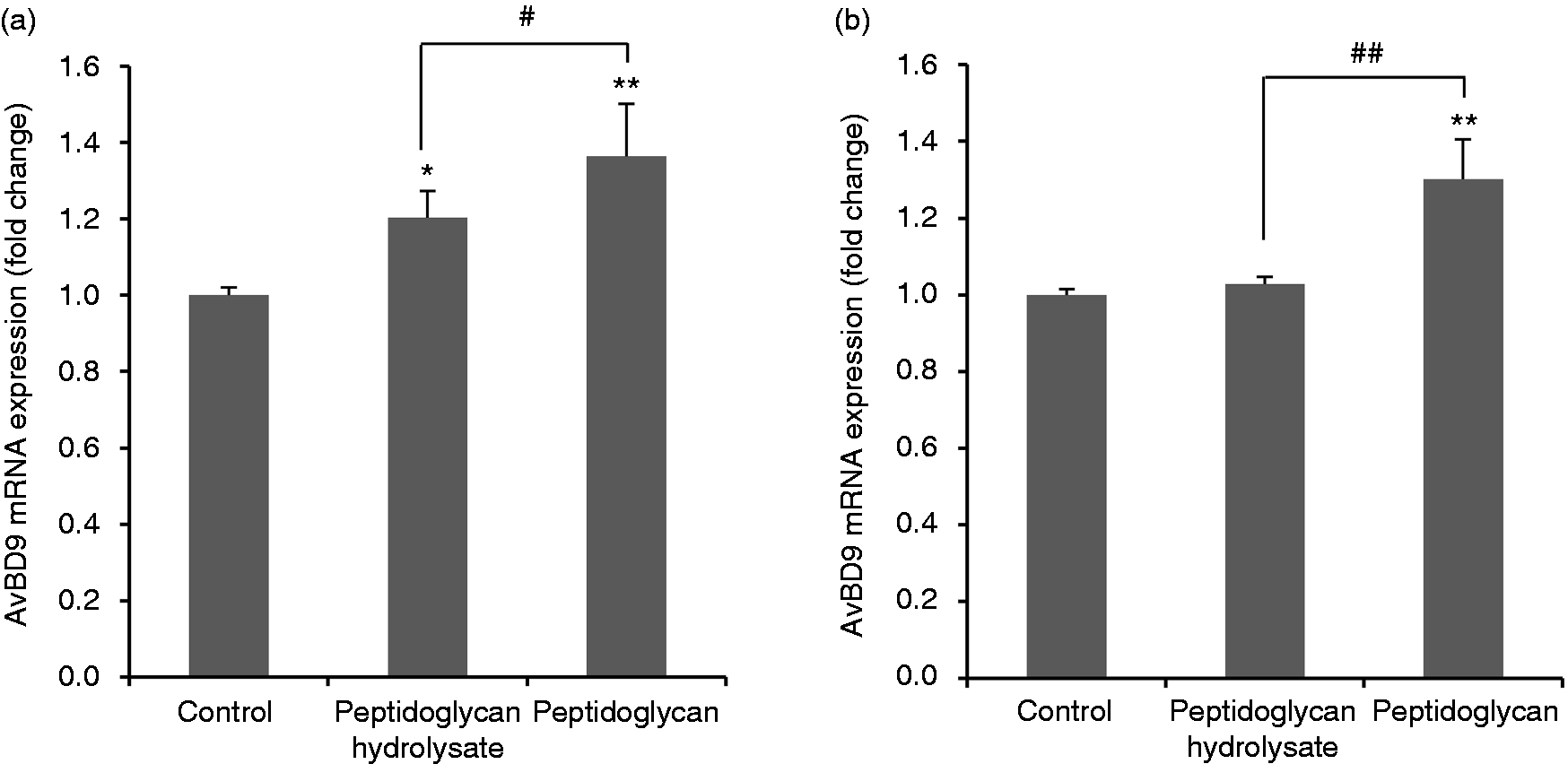
Avian β-defensin 9 (AvBD9) mRNA expression in primary chicken PBMCs (a) and splenocytes (b) in response to L. rhamnosus MLGA-derived intact peptidoglycan or its hydrolysate obtained by lysozyme digestion. Primary cells were incubated with 100 µg/ml of L. rhamnosus MLGA-derived intact peptidoglycan or its hydrolysate for 6 h. Real-time RT-PCR was performed to determine the AvBD9 mRNA expression, and the relative gene expression was calculated using the comparative 2-ΔΔCt method with the GAPDH gene as a reference for normalization and presented as fold-expression change relative to the control group. Values are given as the mean ± SD of two to three independent experiments. *P < 0.05, **P < 0.01 compared with control group; #P < 0.05, ##P < 0.01 between the indicated treatments.
Peptidoglycan from pathogenic bacterium suppresses AvBD9 expression in primary chicken PBMCs and splenocytes
Peptidoglycan is a major cell wall component of Gram-positive bacteria. However, subtle alterations in structural molecules of peptidoglycan in various bacteria cause different interactions between the host cells and microbes, and thus elicit differences in immunobiological effects. 27 , 36 Therefore, we tested the difference in the capacity to modulate AvBD9 expression between peptidoglycans from probiotic L. rhamnosus MLGA and pathogenic S. aureus. In contrast to peptidoglycan from L. rhamnosus MLGA, S. aureus peptidoglycan down-regulated AvBD9 mRNA expression in both PBMCs and splenocytes (Figure 3a and b, respectively).
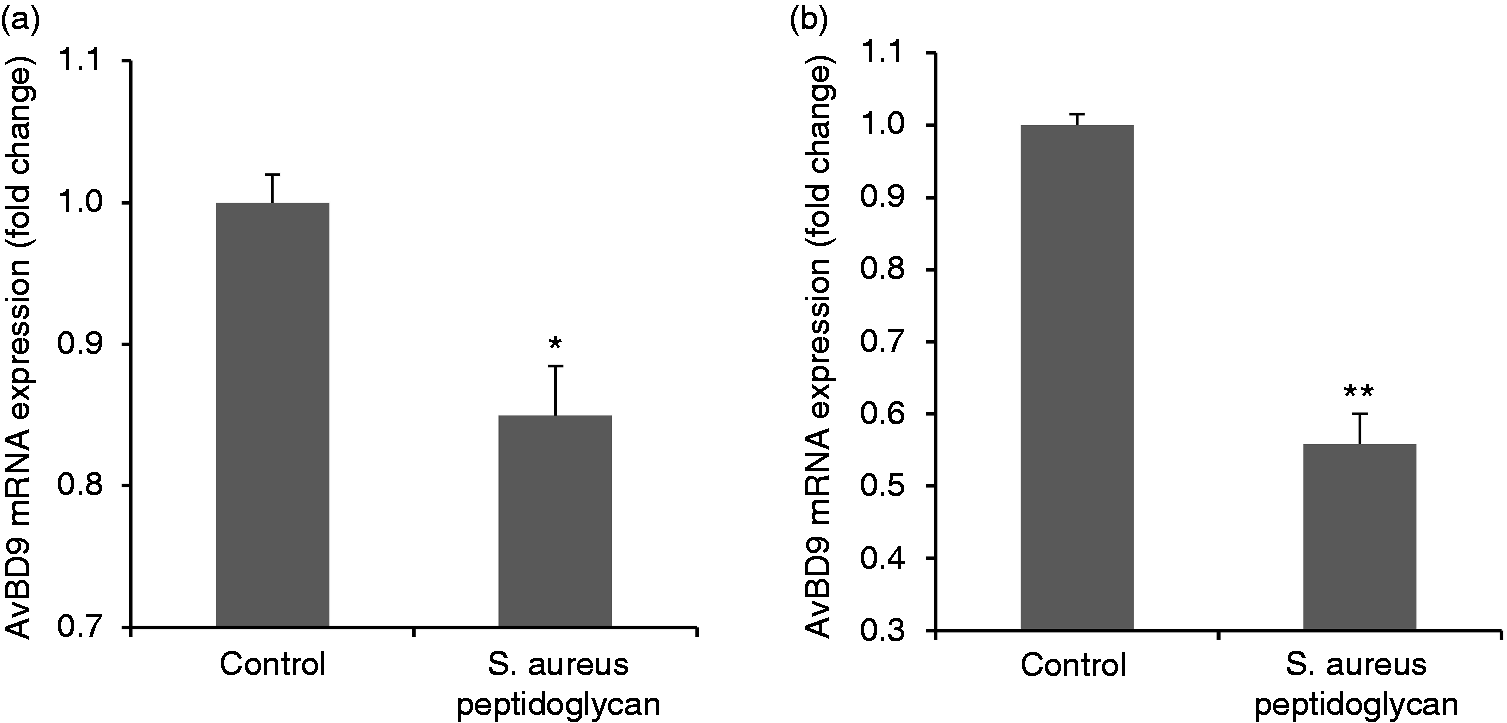
Peptidoglycan from pathogenic Staphylococcus aureus suppresses avian β-defensin 9 (AvBD9) expression in primary chicken PBMCs (a) and splenocytes (b). Cells were incubated with S. aureus-derived peptidoglycan at a concentration of 100 µg/ml for 6 h. Real-time RT-PCR was performed to determine the AvBD9 mRNA expression. Values are given as the mean ± SD of two to three independent experiments. *P < 0.05, **P < 0.01 compared with control group.
Peptidoglycan from L. rhamnosus MLGA triggers no inflammatory response in PBMCs and splenocytes
Pro-inflammatory cytokines are known to be potent inducers and up-regulators of defensins in humans 37 and chickens. 38 To examine whether the up-regulation of AvBD9 expression by L. rhamnosus MLGA-derived peptidoglycan or its hydrolysate by lysozyme was mediated by pro-inflammatory cytokines synthesized in response to peptidoglycan stimulation, we treated PBMCs and splenocytes with and without peptidoglycan or its hydrolysate for 6 h and analyzed the expressions of three representative pro-inflammatory cytokines including IL-1β, IL-8, and IL-12p40. Bacterial LPS from Escherichia coli 055:B5 (Solarbio) at 1 μg/ml was used as a positive control. Intact peptidoglycan showed no induction of all three cytokines both in PBMCs and in splenocytes (Figure 4). Moreover, IL-1β and IL-8 expression was significantly decreased in splenocytes by intact peptidoglycan at 100 and 50 μg/ml, respectively (Figure 4a and b). In contrast, IL-1β, IL-8, and IL-12p40 expressions were markedly induced in response to LPS (Figure 4). Interestingly, L. rhamnosus MLGA peptidoglycan hydrolysate significantly suppressed IL-1β expression in PBMCs and splenocytes (Figure 4a), IL-8 expression in splenocytes (Figure 4b), and IL-12p40 expression in PBMCs (Figure 4c). These results demonstrated the L. rhamnosus MLGA-derived peptidoglycan and its hydrolysate by lysozyme promote AvBD9 expression without being accompanied by inflammatory response, suggesting that AvBD9 expression induction by peptidoglycan from L. rhamnosus MLGA is pro-inflammatory cytokine-independent.
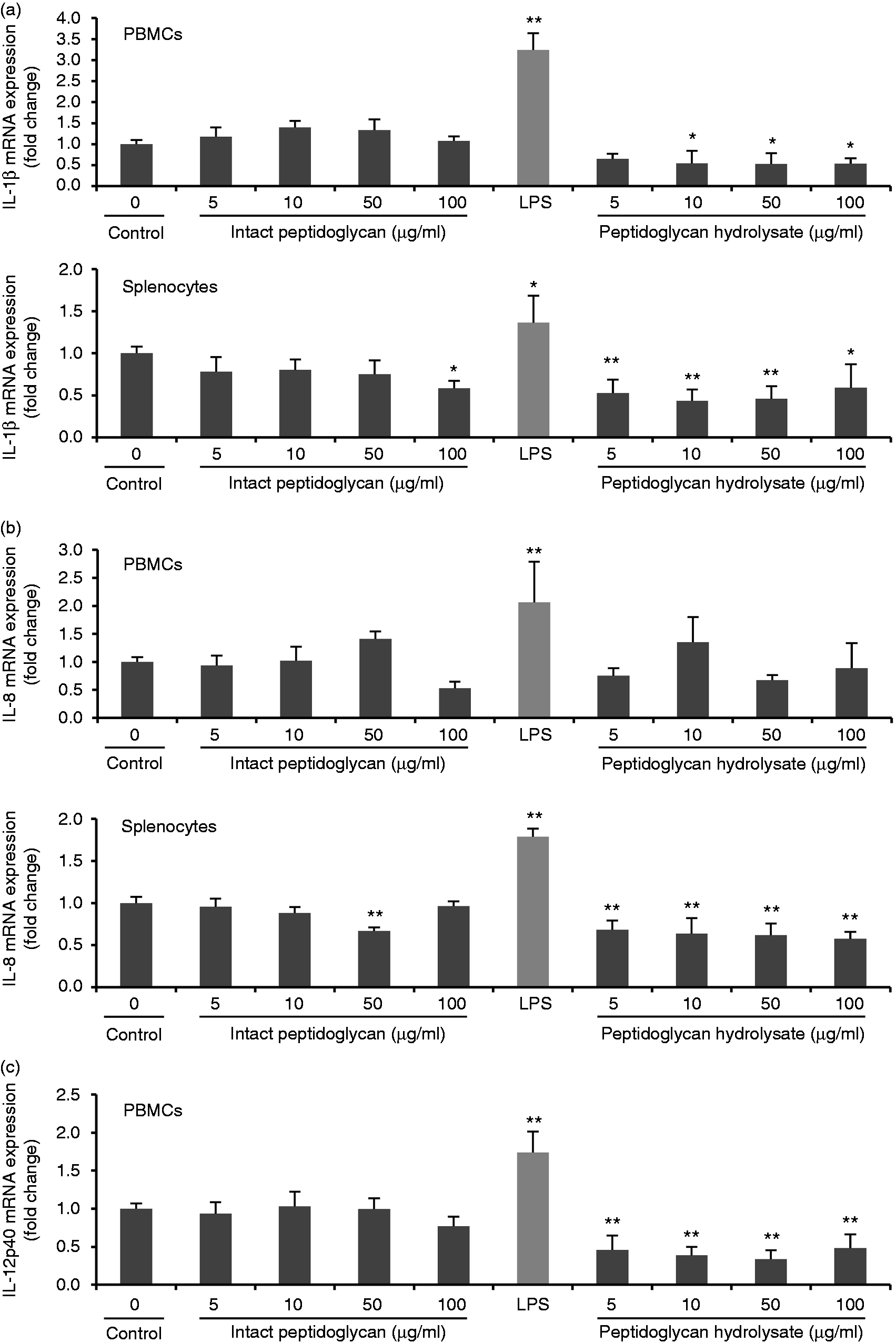
Peptidoglycan from L. rhamnosus MLGA triggers no inflammatory response in chicken PBMCs and splenocytes. Chicken PBMCs or splenocytes were stimulated with indicated concentrations of peptidoglycan or its hydrolysate by lysozyme, or 1 μg/ml LPS as a positive control for 6 h. Real-time RT-PCR was performed to determine the mRNA expressions of IL-1β (a), IL-8 (b), and IL-12p40 (c), and the relative gene expression was calculated using the comparative 2-ΔΔCt method with the GAPDH gene as a reference for normalization and presented as fold-expression change relative to the control group. Values are given as the mean ± SD of two to three independent experiments. *P < 0.05, **P < 0.01 compared with control group.
Peptidoglycan from L. rhamnosus MLGA increases antibacterial activity of chicken PBMCs and splenocytes against S. Enteritidis
It is known that chicken AvBD9 exerts broad-spectrum antibacterial activities against both Gram-positive and Gram-negative pathogenic strains. 8 , 9 Because L. rhamnosus MLGA-derived peptidoglycan significantly induced AvBD9 expression in chicken PBMCs and splenocytes, we next investigated the functional consequence of AvBD9 induction in PBMCs and splenocytes by peptidoglycan from L. rhamnosus MLGA. As shown in Figure 5, both PBMC (Figure 5a) and splenocyte (Figure 5b) lysates pretreated with peptidoglycan significantly inhibited the growth of S. Enteritidis in a dose-dependent manner. However, the supernatant from the co-culture of PBMCs (Figure 5c) or splenocytes (Figure 5d) with L. rhamnosus MLGA-derived peptidoglycan was not able to inhibit the growth of S. Enteritidis probably due to the low secreted AvBD9 into the extracellular medium. It was worth noting that L. rhamnosus MLGA-derived peptidoglycan itself showed no inhibitory effect on the growth of S. Enteritidis (Figure 5e).
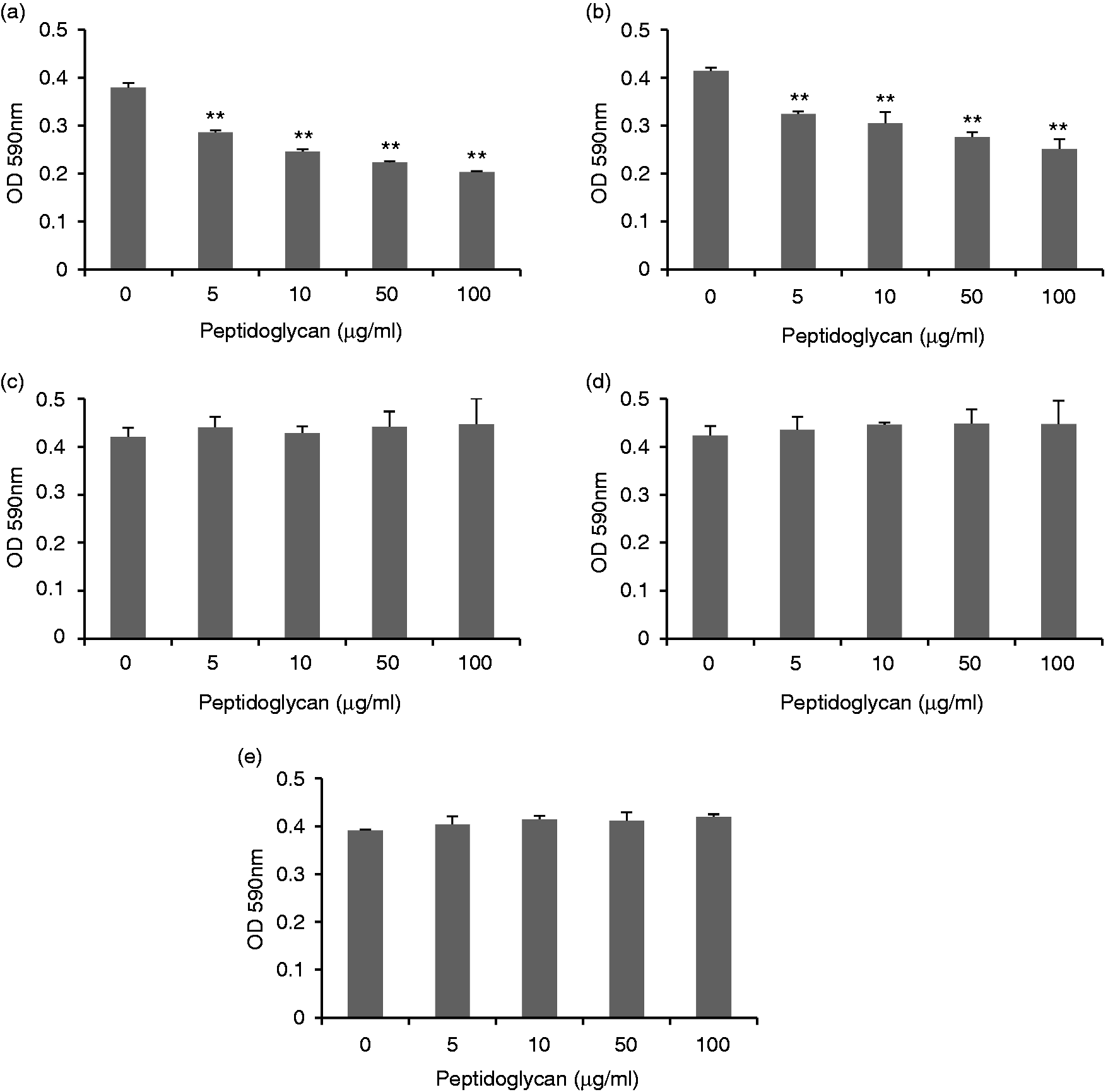
Peptidoglycan from L. rhamnosus MLGA increases antibacterial activity of chicken PBMCs (a) and splenocytes (b). Chicken PBMCs or splenocytes were treated with or without different concentrations of L. rhamnosus MLGA-derived peptidoglycan for 6 h. Cell lysates were them prepared and incubated with S. Enteritidis (ATCC 13076) for 9 h at 37°C. Bacterial turbidity at OD590 nm was measured as an indication of the bacterial density. S. Enteritidis was also directly incubated with different concentrations of L. rhamnosus MLGA-derived peptidoglycan in cell culture medium alone without chicken primary cells (c) or incubated with supernatants from co-culture of PBMCs or splenocytes with L. rhamnosus MLGA-derived peptidoglycan (d). Values are given as the mean ± SD of two to three independent experiments. **P < 0.01 compared with control group.
Discussion
Peptidoglycan is a dominant cell wall component of Lactobacillus bacteria and is constitutively released from the gut microbiota into the intestinal lumen, 36 , 39 and can be absorbed and then circulate throughout the host. 40 , 41 The immunomodulating effects of peptidoglycans from probiotic Lactobacilli that predominantly colonize the small intestine of the host on the intestinal immune system have been extensively studied. Increasing evidence demonstrates that peptidoglycan from commensal lactic acid bacteria not only affects gut homeostasis but also has substantial effects on systemic immune responses. 19 , 27 , 42 Nevertheless, the molecular mechanisms of immunomodulation induced by peptidoglycans are not completely understood. Our results showed that intact peptidoglycan from L. rhamnosus MLGA promotes AvBD9 mRNA expression with different potential in chicken embryo jejunum, ileum, and cecum explants, as well as PBMCs, splenocytes, thymocytes, and hepatocytes in a dose-dependent manner. Of note, peptidoglycan hydrolysates (fragments) obtained by lysozyme digestion also has the capacity to induce AvBD9 expression in PBMCs. Furthermore, we observed that L. rhamnosus MLGA-derived peptidoglycan treatment dose dependently enhanced the antibacterial activity of both PBMC and splenocyte lysates against S. Enteritidis, whereas L. rhamnosus MLGA-derived peptidoglycan itself showed no inhibitory effect on the growth of S. Enteritidis, implying that an enhancement in the antibacterial activity of the peptidoglycan-treated cell lysates is, in part at least, due to the augmentation of AvBD9 expression. Based on these results, we propose that the defensin-inducing potential of peptidoglycans from specific probiotic bacteria species in immune cells and gut intestinal tissue of the host is another manner and mechanism of action through which the probiotic strains exert their immunomodulatory and anti-infection effects on the host.
Intact peptidoglycan and peptidoglycan fragments can be recognized by specific receptors termed PRRs, including TLRs, NLRs, peptidoglycan recognition proteins, and C-type lectin-like receptors, which are expressed intracellularly or on the cell surface in a variety of cell types, including various epithelial cells and immune cells of the host.
43
,
44
Upon recognition of peptidoglycan or peptidoglycan-derived fragments, PRRs initiate downstream signaling cascade events that lead to the expression and secretion of a broad range of innate immune effector molecules, including cytokines, chemokines, interferons, host defense peptides such as defensins and cathelicidins, and to the onset of adaptive immune responses.
45
The role of TLRs and NLRs in mediating the induction of β-defensin expression by peptidoglycan or its fragments has been previously demonstrated.
34
,46–48 In the present study, we showed that peptidoglycan hydrolysates (fragments) also promoted AvBD9 expression in PBMCs albeit at a lesser magnitude than intact peptidoglycan from L. rhamnosus MLAG. However, the inducible expression of AvBD9 by peptidoglycan hydrolysates was diminished in splenocytes as compared to intact peptidoglycan. The difference in AvBD9 induction by peptidoglycan hydrolysates between PBMCs and splenocytes may be ascribed to the different expression profile of PRRs between these two kinds of immune cell populations. For example, cytosolic bacterial peptidoglycan sensor NOD1, an NLR family member, is expressed in a variety of different cell types and tissues and is activated by peptidoglycan fragments containing meso-diaminopimelic acid primarily found in Gram-negative bacteria.
49
In this sense, the peptidoglycan from Gram-positive L. rhamnosus is unable to be detected by NOD1. Conversely, NOD2 expression is restricted to hematopoietic cells and the epithelium of barrier tissues such as the skin, lungs and gastrointestinal tract.
50
NOD2 recognizes N-acetylmuramic acid-containing peptidoglycan fragments with N-acetylmuramyl-
Studies have shown that peptidoglycans from probiotic lactic acid bacteria stains specifically provoke a pro-inflammatory response,
18
,
52
or exhibit anti-inflammatory activity mainly depending on the subtle difference in the peptidoglycan structure.
23
,
24
,
36
,
53
Comparison of the structure of peptidoglycans from various kinds of microorganisms showed the difference in the third amino acid (
Although considered a broad-spectrum pattern-recognizing system, innate immunity can detect very subtle differences in Gram-positive walls. 57 Subtle structural modifications to peptidoglycan can influence the ability of the innate immune system to detect bacteria and can allow bacteria to evade or alter host defenses. 42 In the present study, we found that, in contrast to the effect of L. rhamnosus MLGA-derived peptidoglycan, peptidoglycan from the pathogenic S. aureus suppressed AvBD9 expression, which provides evidence that down-regulated expression of a specific β-defensin such as AvBD9 may be an immune escape strategy used by the specific pathogen in chicken infections.
Conclusion
In summary, this study revealed a new possible immunomodulatory mechanism by which the peptidoglycan from L. rhamnosus MLGA enhances the innate defense response through the up-regulation of β-defensin such as AvBD9 expression in the gut as well as immune cells from peripheral tissues of chicken. More importantly, L. rhamnosus MLGA-derived peptidoglycan triggers no inflammatory response in immune cells while promoting β-defensin gene expression. Moreover, this work points out the possible potential use of peptidoglycan from L. rhamnosus as an alternative to feed antibiotic growth promoters in poultry production. However, further work is needed to explore the potential of L. rhamnosus-derived peptidoglycan to enhance the expression of antimicrobial peptides in pathogen-challenged chickens.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (Grant Numbers: 31360555, 31560641) and Key Project of the Natural Science Foundation of Jiangxi Province ((Grant Number 20181ACB20015).