
Abstract
Low concentration of LPS can be detected in healthy mammals without triggering systemic inflammation. Here we analysed the influence of the mycotoxin deoxynivalenol (DON) on very low LPS concentrations and the role of DON in the physiology of pigs challenged with high artificial LPS dosage mimicking septic shock. Pigs were fed for 29 d with DON-contaminated (4.59 mg/kg feed) or control feed. Samples of control animals showed 6.6 ± 13.5 pg/ml LPS in portal and 3.1 ± 7.6 pg/ml LPS in jugular serum samples. In the DON fed group, 3.4 ± 7.2 pg/ml and 0.6 ± 0.8 pg/ml were detected. The differences were statistically not significant, indicating that DON is not a trigger for enhanced LPS transfer into the blood circulation. Next, pigs were challenged with 7.5 µg LPS/kg body mass via portal or jugular route. The application route did not significantly influence the LPS concentration. We expected higher circulating LPS concentrations in the presence of DON due to the additional stress of liver metabolism and reduced liver capacity to remove LPS from circulation. This scenario is supported by tendency. In summary, we found that DON is unlikely to influence LPS transfer in the gut; DON likely reduces the capacity for LPS removal in septic shock conditions.
Introduction
The intestinal barrier separates the gut lumen harbouring large amounts of commensal, and sometimes pathological, bacteria from the internal side of the body – a compartment which is essentially kept free of bacteria or other microorganisms. In comparison to the ‘inside’ of a healthy organism, the gut microbiota harbours infinite numbers of microorganisms and, among their breakdown products, endotoxins. 1 This extremely steep gradient of bacteria and endotoxins is maintained by i) the passive tight junctions system of the epithelial layer working as a passive ‘fence’ and ii) the activity of the innate and adaptive immune system selectively removing extraneous structures escaping the passive border. 2 The appearance of bacteria and endotoxins in the circulation is normally considered as an accidental, pathological situation. 3 In principal, LPS found in dimensions of 10–20 kDa are too large to cross the intact epithelial layer on the paracellular pathway. 4 Under normal circumstances, the intestinal epithelium is injured and recovers during the normal course of digestion and absorption of a meal. 5 It has been shown that endotoxins can also cross the intestinal border mediated by chylomicrons, which increase as a consequence of high-fat diets.1,6 This mechanism is a transcellular pathway; however, other physiological paracellular pathways, such as those activated by zonulin, have also been described. 7 The appearance of LPS beyond the intestinal border triggers immunological responses aimed at scavenging potential invaders or their breakdown products. Before microorganisms or their breakdown products can be removed by immune cells, the undesirable components must be identified and defence mechanisms must be targeted to the intruders without damaging own body structures. In addition to bacterial-derived protein structures (protein Ags), bacterial cell envelope structures (LPS) are the most important indicators used for triggering an immune response. 8 It has been shown that LPS can be detected in non-pathological situations. In a recent review, Gnauck et al. summarized the available data and found that most LPS concentrations in healthy humans were below 10–20 pg/ml. 9
Deoxynivalenol (DON) is a trichothecene mycotoxin produced by Fusarium spp., a common contaminant of various cereals used for feed and food. DON reduces feed intake and mass gain in pigs, acts as an inhibitor of protein synthesis and as an immunomodulator. Moreover, it has been shown in vitro that high dosages of DON affect the tight junctions in the porcine epithelial cells. 10 In the present experiments we therefore analysed i) LPS entry in barrows fed with DON-contaminated feed and ii) the kinetics of infused LPS in jugular and portal blood samples. The investigation was carried out in the framework of a larger animal experiment analysing further aspects of DON feeding and LPS challenge.11–15
Materials and methods
Animal experiment
The animal trial was performed in the Friedrich-Loeffler-Institute (Braunschweig, Germany), approved by the ethical committee of the Lower Saxony State Office for Consumer Protection and Food Safety (file number 33.4-42502-04-13/1274) and conducted according to the European Community regulations concerning the protection of experimental animals and the guidelines of the German Animal Welfare Act. This trial is part of a large project, and data on animal health and physiology are already published elsewhere.11,13–15 Portal and jugular samples of 18 control and 19 DON-fed animals were obtained. In brief, animals (German land race barrows, 10–11 wk old, initial mass: 25.8 ± 3.7 kg, final mass 29 d later: 40.8 ± 0.4 kg) received either control (CON) or DON-contaminated diet (4.59 mg/kg feed) for 4 wk. Pigs were fed 700 g (air-dry matter, ADM) twice daily, provided as slurry. The main components of the diet were barley (533 g/kg ADM), maize (150 g/kg ADM, where 75 g/kg were replaced by DON-contaminated maize for DON groups), soybean meal (200 g/kg ADM), rapeseed (50 g/kg ADM) and soybean oil (20 g/kg ADM). 15 At d 27 of the experiment pigs were surgically equipped with post-hepatic catheters in venae jugularis interna et externa and arteria carotis externa and pre-hepatically in vena lienalis and vena portae hepatis in order to facilitate simultaneous infusion and blood sampling. At d 29, samples of jugular and portal catheters were analysed for LPS content 30 min before feeding. For challenge experiments, LPS (7.5 µg/kg body mass (BM) dissolved in 0.9% saline, Escherichia coli O111:B4, product number L2630, Sigma-Aldrich, St. Louis, MO, USA) or saline (CON) was infused 15 min after morning feeding via vena jugularis externa (post-hepatic administration) and vena lienalis (pre-hepatic administration), respectively, for 1 h. Thus, two dietary groups (CON vs. DON) and three infusion regimens (NaCl, LPSportal, LPSjugular) resulted in six experimental groups. Pigs from each diet were assigned randomly to one of the three infusion regimens. The first abbreviation denotes diet and the second the infusion regimen: CON_CONju-CONpo, CON_LPSju-CONpo, CON_CONju-LPSpo, DON_CONju-CONpo, DON_LPSju-CONpo, DON_CONju-LPSpo. Samples were taken from pre-systemic (portal) and systemic (jugular) catheters 30 min before and 15, 60, 90 and 180 min after start of LPS infusion. The infusion was terminated after 60 min. Time kinetics of experimental groups without LPS challenge were not analysed further. An overview of the schedule is given in Figure 1.
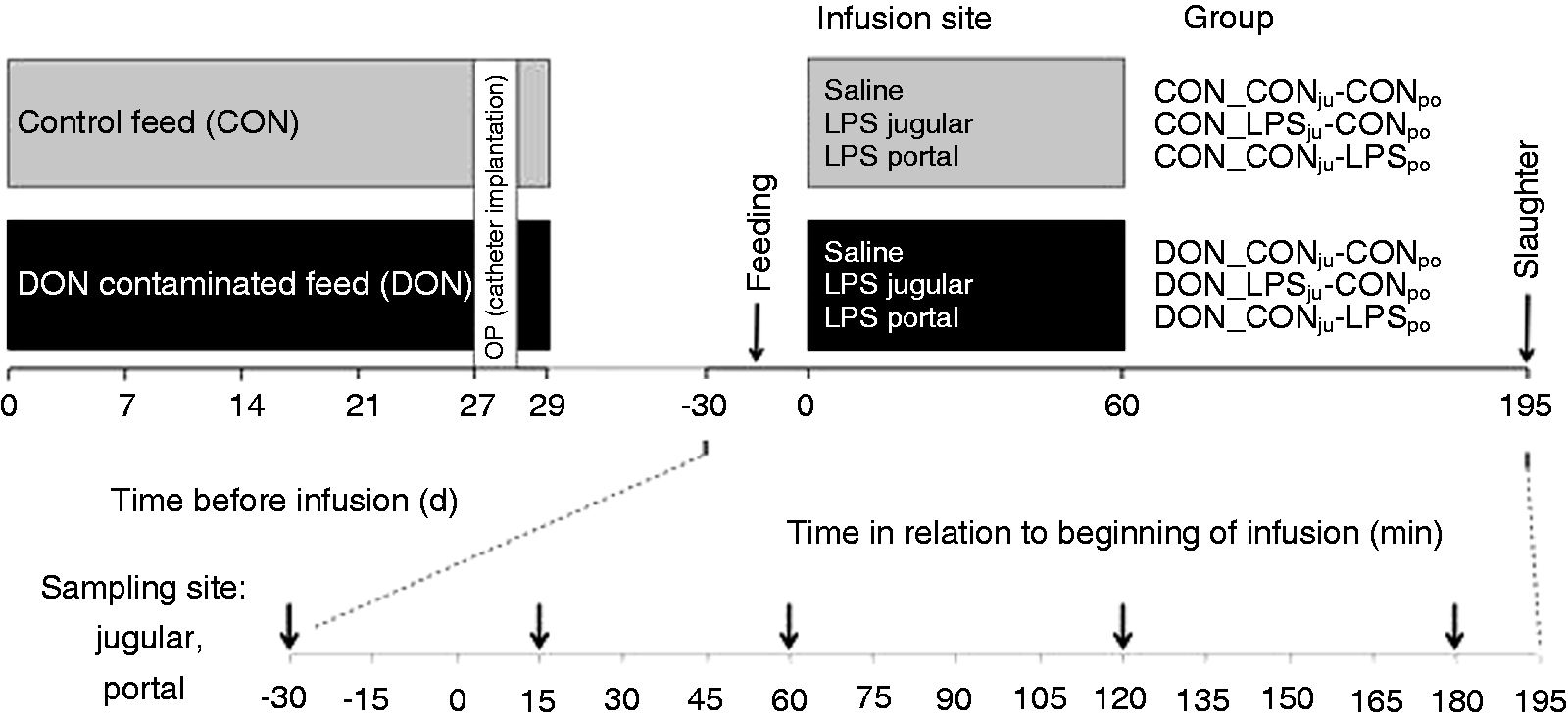
Time schedule of the animal experiment. Barrows received either a control feed (CON, n = 18) or a DON-contaminated feed (DON: 4.59 mg/kg feed, n = 19) for 4 wk; at the end of the experiment, E. coli LPS (7.5 µg/kg BM) or 0.9% saline was infused into jugular (ju) or portal (po) region. Blood samples were taken as indicated presystemic (portal) and systemic (jugular).
Serum and plasma preparation
Blood was taken via the portal and jugular catheter 30 min before LPS/saline infusion and at 15, 60, 120, and 180 min post infusionem (S-Monovette®, Sarstedt, Nümbrecht, Germany). Serum samples were obtained from whole blood by clotting for 60 min at room temperature (21°C) and then for another 30 min at 30°C. Samples were centrifuged at 2123 g for 15 min (15°C), and serum was stored at −80°C until further processing. Heparinized plasma samples were collected in Li-Heparin Monovetten® (Sarstedt). Samples were stored at −80°C until further processing.
LPS assay
Serum LPS content of available samples was analysed with a kinetic limulus amebocyte lysate (LAL) assay (Charles River Endosafe® Endochrome-K™, R1708K, Wilmington, MA) according to the manufacturer’s protocol. Briefly, the test was done in 96-well format at 37°C with a final test volume of 200 μl. All materials used were of certified endotoxin-free quality. Standards and samples were diluted in glass tubes treated for 4 h at 200°C. Control standard endotoxin was diluted with endotoxin-free water to construct a calibration curve up to 10 EU/ml corresponding to 1000 pg/ml. The lowest calculated concentration of the calibration curve was 0.001 pg/ml. Serum and plasma samples were diluted (1:10) with endotoxin-free water and heated for 10 min to 70°C. Reconstituted LAL reagent was prepared according to the recommendations of the individual kit certificate. Standard/sample (100 µl) was mixed at room temperature with 100 µl LAL reagent. The assay was started by warming to 37°C in the plate reader (Tecan infinite 200, Männedorf, Switzerland). The reaction was monitored at 405 nm at 30-s intervals for 2 h (200 cycles). According to the recommendations of the manufacturer, the time span required to reach a threshold of 0.2 was used as readout. Sample concentration was calculated versus linear log-log calibration curve for r >0.980. Otherwise samples were interpolated linearly between neighbouring standards. Samples taken 30 min before the acute phase of the animal experiments starts were routinely spiked with 1 pg/ml standard LPS. Recovery was found between 30% and 250% (manufacturer recommendation: 50%–200%). The lowest concentration that was regularly detectable was 0.05 pg/ml (limit of detection, LOD).
Statistical methods
The Shapiro-Wilk normality test was used for analysing data distribution. Comparisons of two data groups were analysed by one-sided t-test or Mann-Whitney U-Test for non-Gaussian distributed data (heteroscedastic, one-sided). Comparisons covering more than two groups were analysed by ANOVA (normally distributed) or Kruskal-Wallis method (non-Gaussian distributed data). Methods are indicated in figure legends.
Results
In a first step we analysed the LPS level in corresponding serum and heparin samples. Figure 2a shows mean values of 11 samples taken 30 min before jugular LPS challenge. These samples represent the lowest concentration range found in this investigation in healthy animals without experimental LPS challenge. The mean LPS concentrations were 0.99 and 1.16 pg/ml (± 0.46 and 0.43 pg/ml SEM) for serum and plasma source, respectively, and statistically not different (t-test). We further analysed serum and heparin samples 60 min after start of the LPS infusion (Figure 2b). The mean detected LPS concentrations are 358 and 329 pg/ml (± 143 and 138 pg/ml SEM) for serum and plasma of the corresponding samples. The detected concentrations of the samples ranged from 1.7 pg/ml to 1720 pg/ml. We did not find statistical significant differences between serum and heparin plasma samples as a source for LPS measurements (t-test).
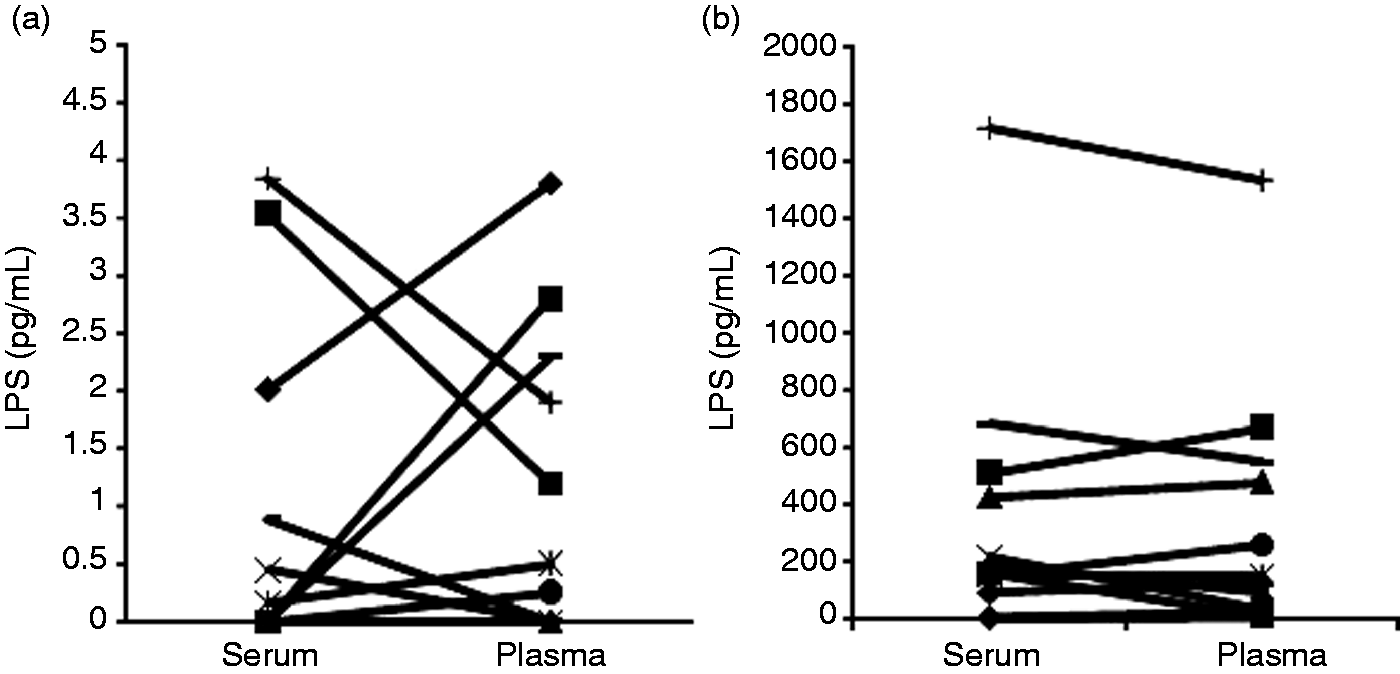
Quantification of LPS in porcine serum and plasma. (a) Serum and corresponding plasma samples (n = 11) of non-challenged (–30 min) pigs were analysed by LAL assay. (b) Serum and corresponding plasma samples (n = 12) of LPS challenged (60 min after beginning of infusion) pigs were analysed by LAL assay. Mean values were statistically not significant between serum and plasma samples (one-sided, heteroscedastic t-test, P > 0.05).
Blood samples were taken from portal and jugular catheter of DON-fed and CON-fed animals 30 min before infusion of LPS, and initial LPS level was quantified in serum (Figure 3). In the CON-fed group (n = 18), portal serum LPS was 6.6 ± 13.5 pg/ml and 3.1 ± 7.6 pg/ml in jugular samples. Results for the DON-fed group (n = 19) were 3.4 ± 7.2 pg/ml portal and 0.6 ± 0.8 pg/ml jugular. Data did not show normal distribution and differences in LPS contents were not significant (Kruskal-Wallis, SD). Prominent high LPS values (above mean value) detected in portal sampling were not linked to prominent increased values at jugular sampling in the same animal (except pig T68: 27.3 pg/ml portal and 31.0 pg/ml jugular) and vice versa. Numerically higher LPS concentrations were found in portal serum (efferent blood stream from intestine) in comparison to jugular samples in both CON and DON-fed animals. Due to the strong variability within the groups, these differences were statistically not significant.
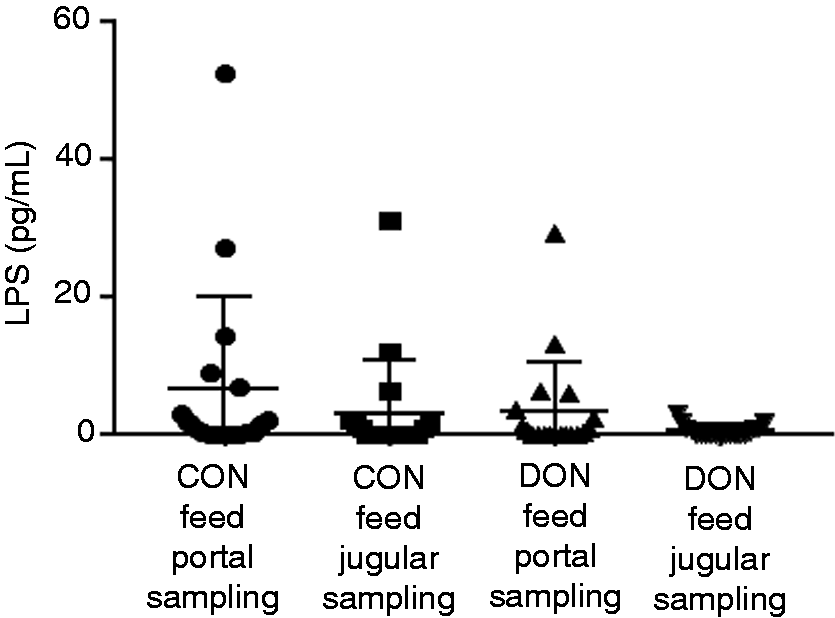
LPS content in portal and jugular serum of CON- and DON feed animals 30 min before infusion. Mean value and standard deviation are indicated. The groups are not significant different (Kruskal-Wallis, ± SD).
In the second part of the investigation, DON and CON-fed animals (13 and 12) were challenged with an LPS-infusion for 1 h. The final dosage of LPS was 7.5 µg LPS/kg body mass. LPS was applied via portal (CON_CONju-LPSpo, n = 6; DON_CONju-LPSpo, n = 6) or jugular catheter (CON_LPSju-CONpo, n = 6; DON_LPSju-CONpo, n = 7). Samples were taken from the corresponding catheter not used for infusion. In Figure 4, individual LPS kinetics are shown for both configurations. In general, in most animals, the application of LPS led to a continuous increase in serum samples up to the end of application at 60 min. The initial LPS concentration was measured 30 min before application of LPS starts. As these values are small in comparison to most LPS contents measured during application, we set initial measurements as start points at time 0 min. The measured LPS levels are highly variable in different animals independent of experimental configuration or feeding group. The LPS levels found in two animals (T20, CON_LPSju-CONpo; T14, DON_LPSju-CONpo) were barely above the LPS level detected in unchallenged animals. Other animals (e.g. T11, DON_LPSju-CONpo; T63 and T54, DON_CONju-LPSpo) showed LPS level that were more than 1000-times higher than the untreated initial contents. Peak concentrations after 60 min are between 1.7 and 1720 pg/ml. In 21 of 25 animals the maximal LPS levels were found at 60 min, and a distinct decrease was seen 30 min after the end of infusion at time 90 min. After 180 min the LPS concentration remained elevated in different degrees in 23 of 25 animals.
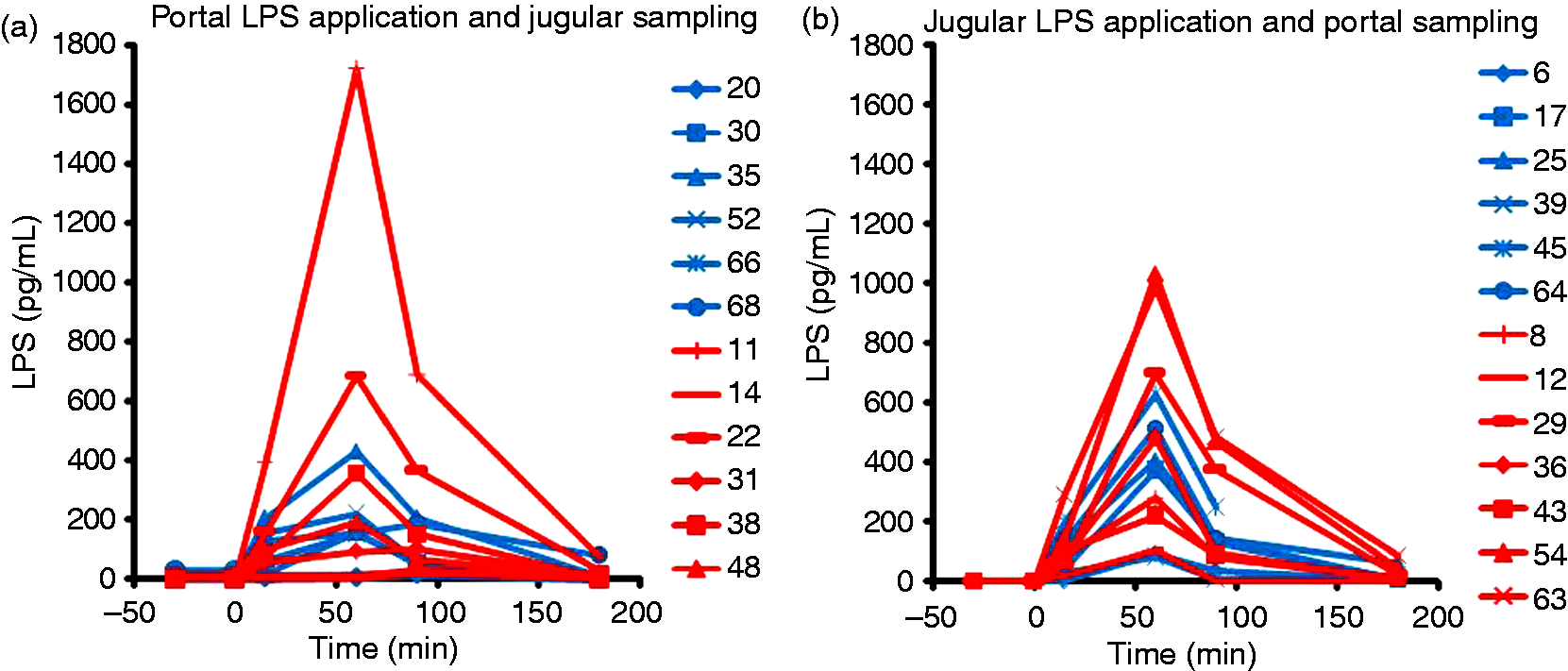
Blood LPS-kinetic of individual pigs challenged by LPS infusion. (a) LPS was applied via portal catheter and samples were taken from jugular catheter. Blue traces represent CON-fed animals, red traces DON-fed. (b) LPS was applied via jugular catheter and samples were taken from the portal side. Blue traces represent CON-fed animals, red traces DON-fed. Infusion started at t = 0 min. The low values at t = –30 min represent the untreated LPS-level before feeding and infusion. Numbers in legend indicate individual animals.
The kinetic data of the LPS challenge experiment shown above were integrated, and the LPS content in circulation was calculated as area under curve (AUC). Data passed the Shapiro-Wilk normality test, and ANOVA indicated no significant differences between groups (Figure 5a). However, the scatter plot in Figure 5a also showed that integral LPS values of five animals of the DON-fed groups were higher than the highest value in the CON-fed group. Therefore, data of CON- and DON-fed were cumulated independent of the application scheme (Figure 5b). Data were normally distributed and a tendency (P = 0.056) to higher integral LPS was found in DON-feed group.
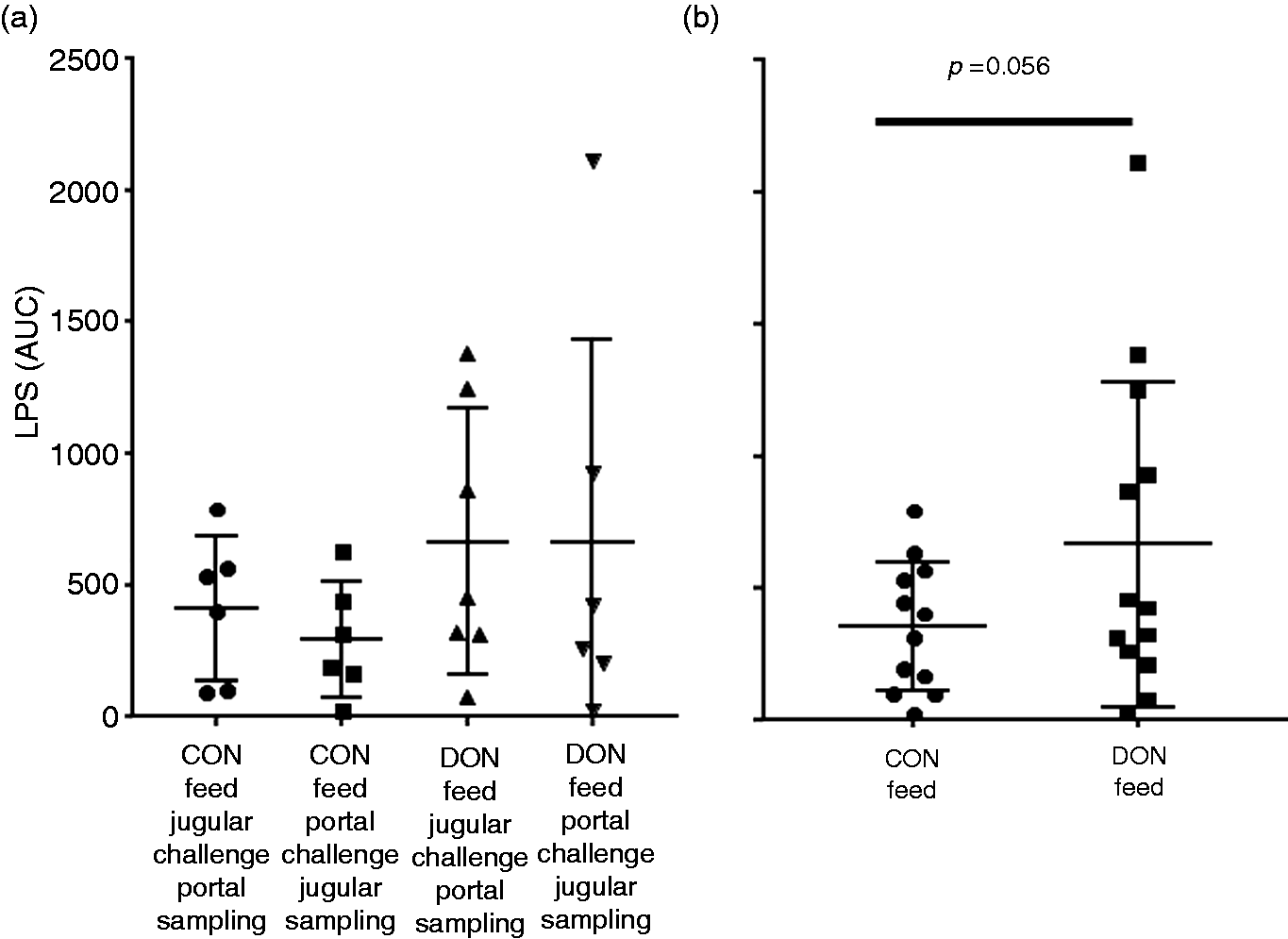
Integral LPS signal (area under the curve, AUC) in CON and DON-fed pigs challenged with 7.5 µg/kg LPS. (a) Individual groups. Feeding, challenge and sampling scheme as indicated. Mean values are not statistically significant (ANOVA). (b) Cumulated data from (A). Integral LPS is higher in DON-fed groups by tendency (one-sided, heteroscedastic t-test, P > 0.05).
Discussion
Detection and quantification of LPS in biological fluids such as serum or plasma is a challenge. The currently available LAL assay is a highly sensitive tool in standard solutions. As an example, commercially available tests allow quantification between 0.005 and 50 EU/ml in non-interfering fluids. In biologic fluids such as whole blood or the corresponding fractions the LAL assay can interfere with blood components or agents used for preparations of fractions. 16 Moreover, LPS can be attached to the surface of fibrin fibres and trapped in the fibrin clot. 17 In our hands, we did not find significant differences comparing detected LPS concentration between serum and heparin sources. One reason could be that the majority of LPS (97%) is bound to HDL, LDL and VLDL.6,18 Lipoproteins themselves can be quantified in serum and we concluded that the difference between both sources in respect of LPS detection is negligible.
In previous in vitro investigations, we could show that the mycotoxin DON affects the epithelial layer by reducing junctional components and consequently increased permeability measured as reduced transepithelial resistance. 19 Due to this observation, we hypothesised that the presence of DON in vivo increased the transfer of LPS across the epithelial barrier.
LPS are derived from the outer membrane of Gram-negative bacteria and trigger the innate immune system in low concentrations when occurring within the circulation. It has been suggested that LPS is transported by colon epithelial cells.20,21 Only a few measurements have so far been made in the portal circulation, which is the principal entry route from the digestive system. For example, in mouse portal vein, concentrations of 10 pg/ml (median) were reported. 22 Comparable concentrations (0.9 EU/ml, approximately 9 pg/ml) were found in rat. After ischemia, the intestinal border showed massive denudation and LPS concentration in the portal vein increased by a factor of 2.5. 23 Different factors influence the intestinal permeability. LPS transfer from the intestinal lumen to the circulation occurs frequently even in non-pathological situations. 24 It has been shown that food intake alone increases LPS level in healthy adults, and depends on meal composition. 25 Especially high-fat-containing food is linked with LPS transfer and the terms postprandial endotoxemia/inflammation were coined to describe this mechanism.26,27 In humans, a whole clutch of relevant diseases, including type II diabetes and obesity, are likely associated with inflammatory events triggered by gut-born LPS and microorganisms. 28 However, other feed components may also modulate LPS transfer and scavenge. In goats, it was found that high grain feed content was linked to an increased plasma LPS content (113–135 pg/ml) in comparison with low grain feed (47–59 pg/ml). 29 The intestinal epithelium is injured and restitutes during the normal course of digestion and absorption of a meal.5,30 Restitution is achieved by migration of epithelium covering the free basal membrane area. 31 The LPS levels found in this investigation in pig (portal 6.6 ± 13.5 pg/ml and jugular 3.1 ± 7.6 pg/m) were low and comparable to those reported from humans. 9 Clinically relevant is the fact that intestinal ischemia enhances LPS transfer across the barrier, predominately by paracellular permeability and epithelial destruction. 32 The transport can also be influenced by metabolic stress and butyrate. 33
In contrast to our hypothesis, we did not find an increased LPS load in DON-fed animals. As a consequence, an increased latent LPS level in blood circulation is unlikely be a mechanism relevant in DON toxicity.
LPS kinetics in response to infused LPS exhibited large individual variations (Figure 4). The maximal LPS concentration varied between individuals by a factor of 1000. As the infusion stream of LPS was constant, the distribution and/or scavenge capacity responsible for the removal of LPS from the circulation was likely the essential modulator of these kinetics. First in line when LPS entered via the intestine has reached the blood stream is the liver. 34 In non-pathological situations, the majority of LPS will scavenged and a significant lower LPS load in than found in the subsequent systemic circulation. 35 LPS neutralization is also mediated by innate immunity antimicrobial peptides, which bind to LPS and inhibit the immunological, pre-dominantly macrophage mediated response. 36 It has been shown in mice that the structure and length of different LPS types is relevant for the clearance of LPS and bacteria. 37
In a previous investigation, we showed small but significant modifications of liver function in DON-fed pigs. 13 Data from the previous investigation suggests that the DON burden of the liver may reduce the available capacities to scavenge LPS from circulation. The tendentious increase of integral LPS present in the bloodstream is in line with the concept of moderate hampered liver function.
Conclusion
The intestinal tract is an important entry route of LPS into the circulation. In vitro, DON exposure of intestinal epithelial cells resulted in increased paracellular permeability. However, our data in vivo did not support this for the transfer of LPS. A strong tendency suggests that DON acts as an additional stress factor reducing the liver capacity to scavenge LPS after challenge. These findings underline the complex modulatory effects of DON in the organism.
Footnotes
Acknowledgements
The authors would like to thank Romina Wolter for their excellent technical support in sample analysis.
Declaration of conflicting interests
The author(s) declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported financially by Deutsche Forschungsgemeinschaft (DFG DA 558/1-4 and RO 743/3-3).