
Abstract
The present study aimed to investigate the effects of sodium butyrate on the intestinal barrier and mast cell activation, as well as inflammatory mediator production, and determine whether mitogen-activated protein kinase signaling pathways are involved in these processes. A total of 72 piglets, weaned at 28 ± 1 d age, were allotted to two dietary treatments (control vs. 450 mg/kg sodium butyrate) for 2 wk. The results showed that supplemental sodium butyrate increased daily gain, improved intestinal morphology, as indicated by greater villus height and villus height:crypt depth ratio, and intestinal barrier function reflected by increased transepithelial electrical resistance and decreased paracellular flux of dextran (4 kDa). Moreover, sodium butyrate reduced the percentage of degranulated mast cells and its inflammatory mediator content (histamine, tryptase, TNF-α and IL-6) in the jejunum mucosa. Sodium butyrate also decreased the expression of mast cell-specific tryptase, TNF-α and IL-6 mRNA. Sodium butyrate significantly decreased the phosphorylated ratio of JNK whereas not affecting the phosphorylated ratios of ERK and p38. The results indicated that the protective effects of sodium butyrate on intestinal integrity were closely related to inhibition of mast cell activation and inflammatory mediator production, and that the JNK signaling pathway was likely involved in this process.
Keywords
Introduction
Weaning is the most significant event in the life of pigs as they are abruptly forced to adapt to nutritional, immunological and psychological disruptions. 1 Stresses associated with early weaning usually result in growth retardation, post-weaning diarrhea and impaired intestinal barrier of piglets.2–4 It is well known that sodium butyrate is involved in promoting growth, preventing diarrhea and restoring mucosal barrier integrity in piglets.5,6 Generally speaking, the role of sodium butyrate in intestinal integrity is primarily owing to its ability to provide energy for intestinal epithelial cells. 7 Recently, several studies have shown that sodium butyrate exerts anti-inflammatory effects in vitro.8–10 However, whether the beneficial role of sodium butyrate in intestinal integrity of weaned pigs is related to alleviation of intestinal inflammation remains unknown. Moreover, the underlying mechanism also needs to be further investigated.
Intestinal mucosal mast cells reside in the lamina propria underneath the epithelium and play a key role in intestinal inflammation.11,12 Mast cells contain large amounts of preformed compounds commonly referred to as mast cell inflammatory mediators, such as protease, histamine and cytokines.13,14 Once the mast cells are activated, these mediators are released throughout gastrointestinal tracts and cause epithelial barrier dysfunction.15–17 A few recent studies have reported that sodium butyrate inhibits mast cell activation and production of its mediator in vitro.18,19 It would be of interest to investigate whether sodium butyrate influences mast cell activation in vivo. However, no data are available regarding the effect of sodium butyrate on mast cell activation in weaned pigs.
MAPK pathways transduce signals from a diverse array of extracellular stimuli.20,21 Three principal members—ERK, p38 MAPK and c-Jun NH2–terminal kinase (JNK)—comprise this superfamily. 22 Interestingly, Masuda et al. 23 and Zhang et al. 19 reported that in murine bone marrow-derived mast cells, MAPK signaling pathways are involved in the production of mast cell inflammatory mediators; therefore, it is imperative that the effect of sodium butyrate on MAPK activation in weaned pigs is investigated.
Accordingly, we hypothesized that sodium butyrate would improve intestinal barrier function by influencing mast cell activation and inflammatory mediator production through modulation of MAPK signaling pathways. This study was conducted to assess the impact of sodium butyrate on intestinal barrier function and to determine whether MAPK signaling pathways are involved in the protective role of sodium butyrate on intestinal integrity.
Materials and methods
Animals and experiment design
Ingredient and composition of diets on an as-fed basis.
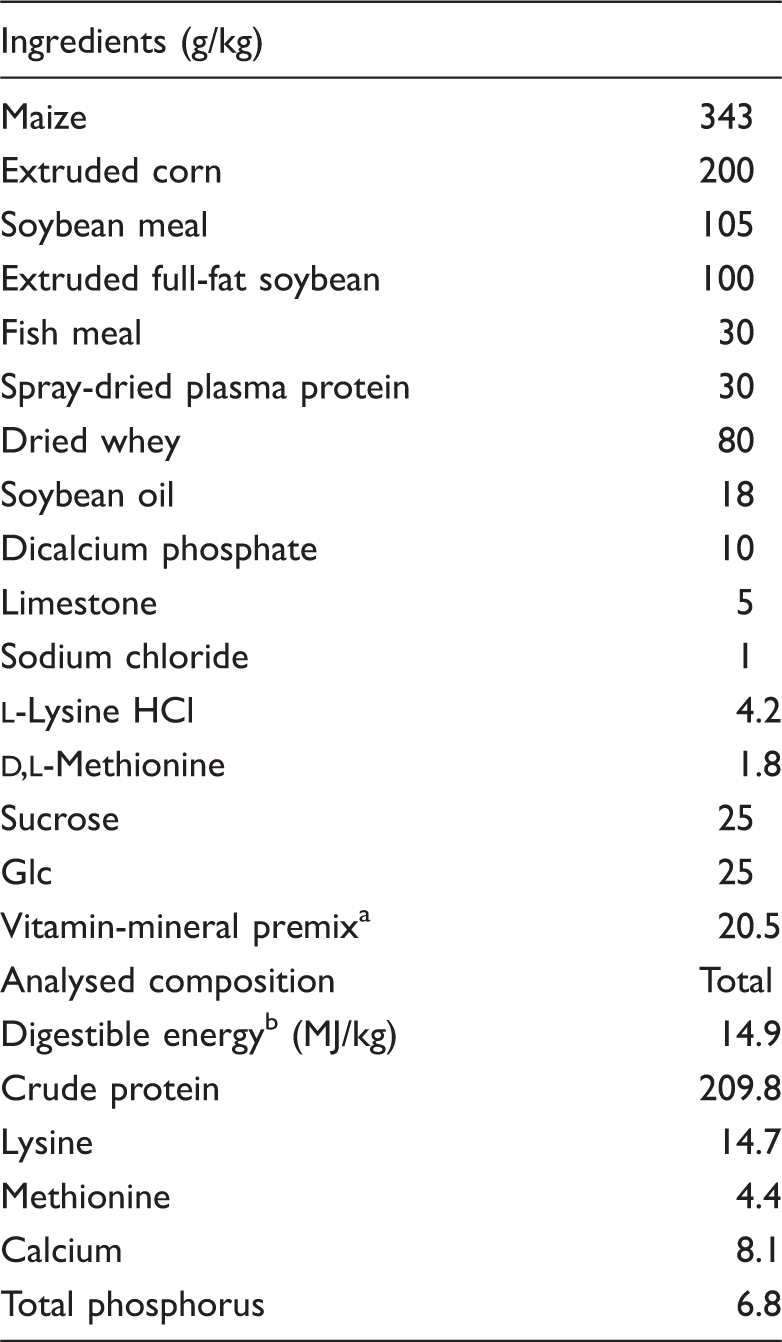
Provided the following per kg of diet: vitamin A, 8000 IU; vitamin D, 2000 IU; vitamin E, 30 IU; vitamin K3, 1.5 mg; vitamin B1, 1.6 mg; vitaminB6, 1.5 mg; vitamin B12, 12µg; niacin, 20 mg; d-pantothenic acid, 15 mg; Zn, 80 mg; Fe, 100 mg; Cu, 20 mg; Mn, 25 mg; I, 0.3 mg; Se, 0.2 mg. bDigestible energy was calculated from data provided by Feed Database in China (2011).
Sample collection
After the feeding trial (d 14 post-weaning), six piglets from each treatment (one pig from every pen) were euthanized with a dose of sodium pentobarbital (200 mg/kg of body mass) according to Chen et al. 24 Specimens (1 cm) of the proximal jejunum were fixed in 10% formalin for morphology measurements. Adjacent specimens were prepared for Ussing chamber studies. The mucosa samples from remaining jejunum were collected, rapidly frozen in liquid nitrogen and stored at −80℃ until analysis.
Intestinal morphology
Segments for morphological study were embedded in paraffin, sectioned and stained with hematoxylin and eosin. Villus height and crypt depth were determined with an image processing and analysis system (Leica Imaging Systems, Cambridge, UK).
Ussing chamber experiments
Transepithelial electrical resistance (TER) and fluorescein isothiocyanate dextran 4 kDa (FD4) were measured in an Ussing chamber system. Segments of the jejunum were stripped from the seromuscular layer in oxygenated Ringer’s solution and then mounted in an EasyMount Ussing chamber system (model VCC MC6; Physiologic Instruments, San Diego, CA, USA) as described previously. 4 In brief, the clamps were connected to Acquire and Analyse software (Physiologic Instruments) for automatic data collection. After a 15-min equilibration period on Ussing chambers, TER was recorded at 15-min intervals over a 1-h period. FD4 (Sigma-Aldrich, St. Louis, MO, USA) was added to the mucosal side at the final concentration of 0.4 mg/ml. The concentration of FD4 was measured by a fluorescence microplate reader (FLx800; BioTek Instruments, Winooski, VT, USA).
Mast cell counting
Sections of jejunum were prepared and then stained with toluidine blue. Sections were viewed at × 20 objective and data were presented as percentage of degranulated mast cells. The degranulated mast cells included mast cells that released > 50% of intracellular granules around the cell. 25 Mast cell counts were conducted on five different fields per slide and six slides per treatment. All cell counts were performed by at least two reviewers who were blinded to experimental treatments.
Mast cell inflammatory mediator analysis by ELISA
Jejunum mucosa was homogenized in PBS and the supernatant was collected. Samples were then diluted 1:10 in PBS and assayed for histamine, tryptase, TNF-α, IL-6 and IFN-γ using a commercial porcine ELISA assay (R&D Systems, Minneapolis, MN, USA). 26
mRNA expression analysis by real-time PCR
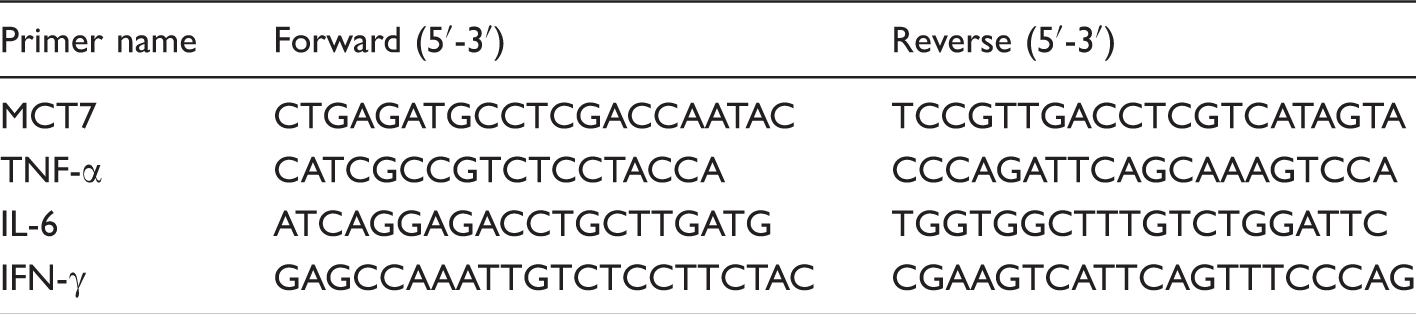
Primer sequences used for real-time PCR.
Protein expression analysis by Western blot
The method for Western blot analysis was the same as the procedures outlined by Hu et al. 4 In brief, after electrophoresis, the proteins were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA). The membranes were incubated with primary Ab at 4℃ for 12 h, then with the secondary Ab for 1 h at room temperature (25–27℃). The primary Abs [p38, phospho-p38, JNK, phospho-JNK (p-JNK), ERK, phospho-ERK (p-ERK)] and the secondary Ab (HRP-conjugated anti-rabbit Ab) were all purchased from Cell Signaling Technology (Danvers, MA, USA). Western blot was done with an enhanced chemiluminescence detection kit (Amersham, Arlington Heights, IL, USA), photographed by a ChemiScope 3400 (Clinx Science Instruments, Shanghai, China) and analyzed using Quantity One software. The values were calculated as the ratios of the phosphorylation levels and the total levels of MAPKs (JNK, p38, ERK).
Statistical analysis
Data were analyzed using SPSS 9.0 statistical package (IBM, Armonk, NY, USA). Results are expressed as mean ± SD. Differences between means were tested using Student’s t-test. Differences were considered significant at P < 0.05.
Results
Growth performance
Effects of sodium butyrate on growth performance of weaned pigs.

Data are mean ± SD (n = 6).
Differences were considered significant at P < 0.05.
Intestinal morphology and barrier function
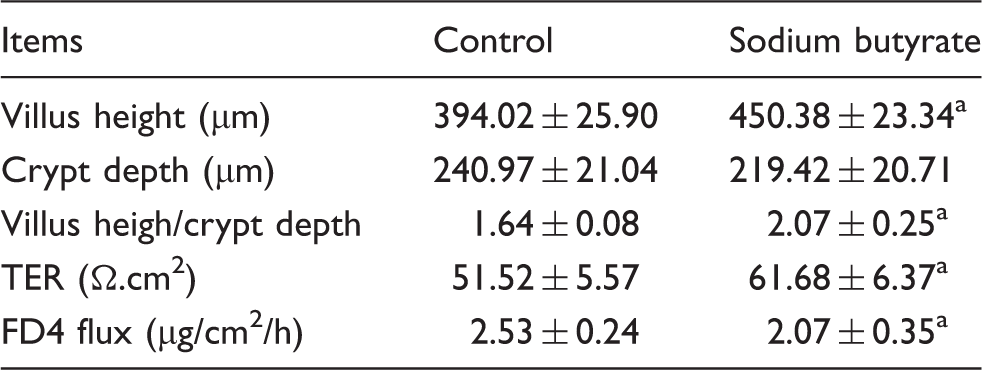
Effects of sodium butyrate on intestinal morphology and barrier function in jejunum of weaned pigs.
Data are mean ± SD (n = 6).
Differences were considered significant at P < 0.05.
Mast cell degranulation
Figure 1 shows the effects of sodium butyrate on mast cell degranulation in intestinal mucosa of piglets. In comparison with the control, dietary supplementation with sodium butyrate significantly reduced the percentage of degranulated mast cells (P < 0.05). However, the total mast cell numbers in jejunum mucosa showed no (P > 0.05) difference between the two groups.
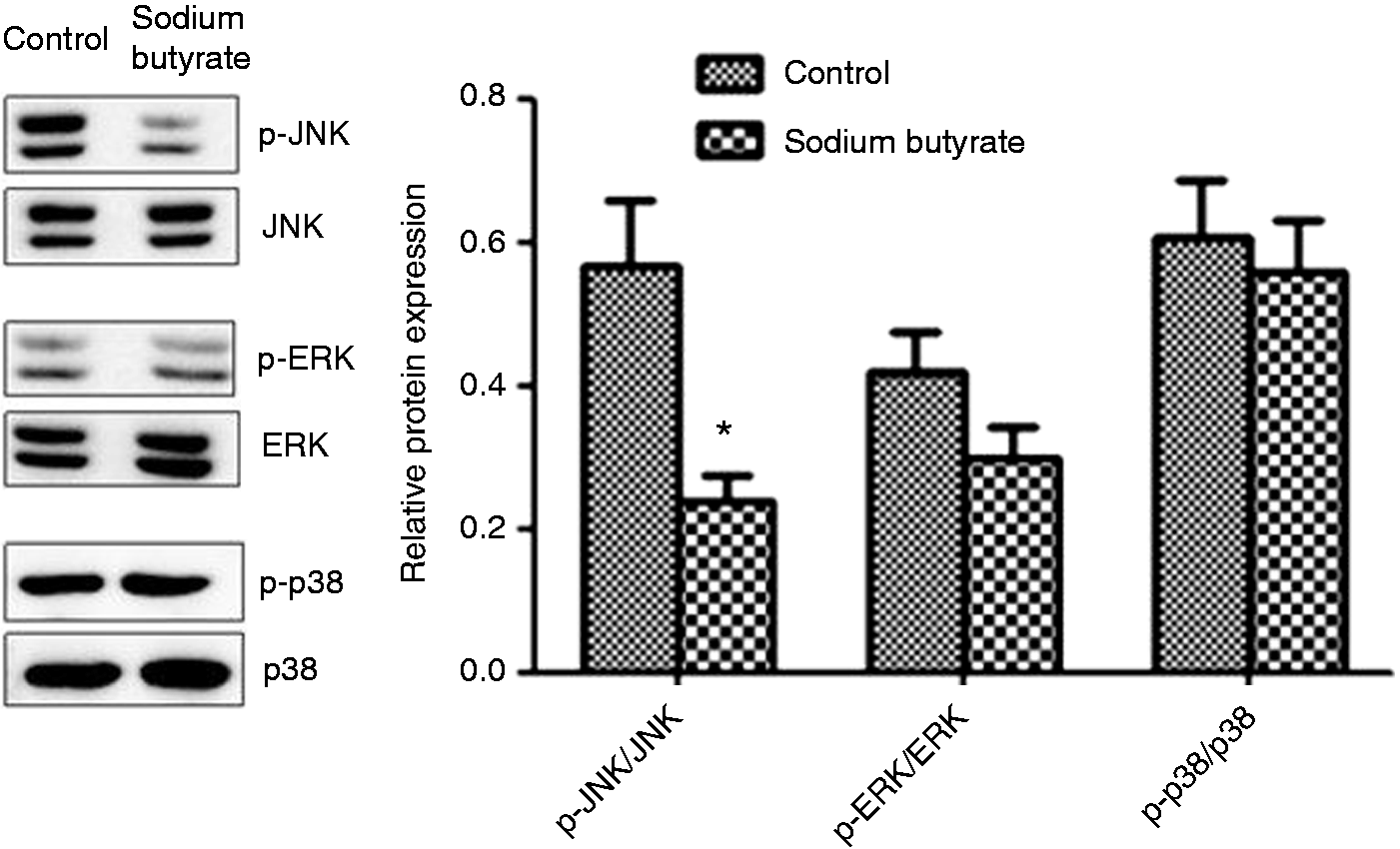
Effects of sodium butyrate on mast cell degranulation in jejunum mucosa of weaned pigs. Values are means ± SD (n = 6). *Differences were considered significant at P < 0.05. Effects of sodium butyrate on MAPK signal pathways in the jejunal mucosa of weaned pigs. The three MAPKs are JNK, p38 and ERK. The bands are representative blots from one of six pigs. The values are calculated as the ratios of their phosphorylation levels (p-JNK, p-p38, p-ERK) and the total levels of MAPKs. *Differences were considered significant at P < 0.05.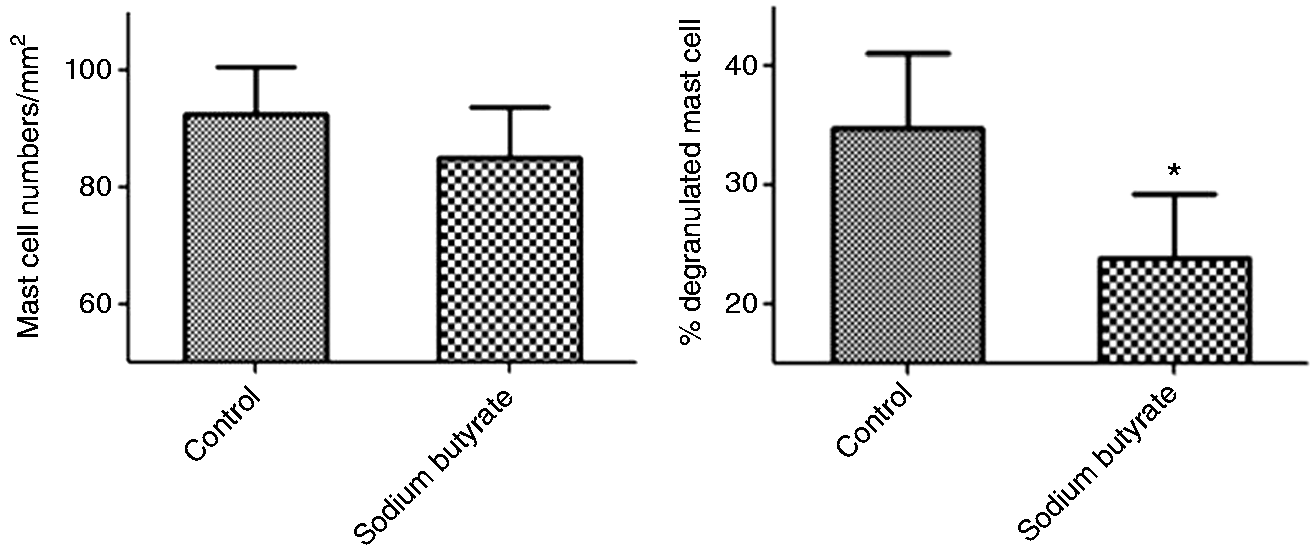
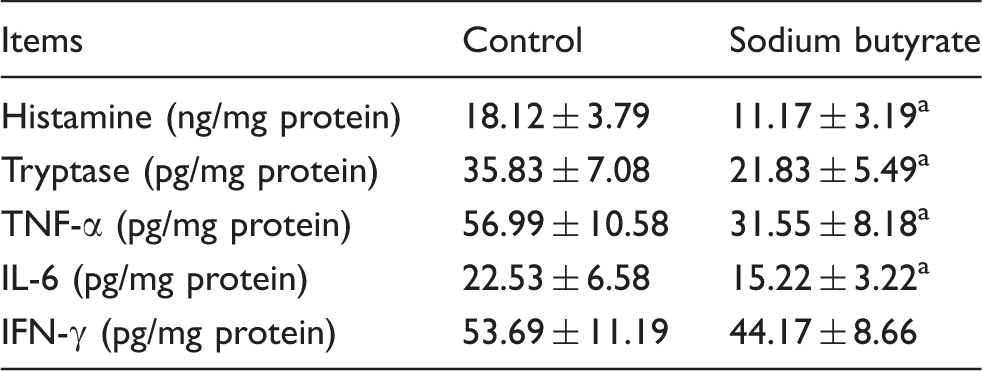
Mast cell inflammatory mediators content
Effects of sodium butyrate on mast cell inflammatory mediators content in jejunum mucosa of weaned pigs.
Data are mean ± SD (n = 6).
Differences were considered significant at P < 0.05.
Intestinal mRNA expression of mast cell inflammatory mediators
Effect of sodium butyrate on relative mRNA expression of mast cell inflammatory mediators in jejunal mucosa of weaned pigs. a
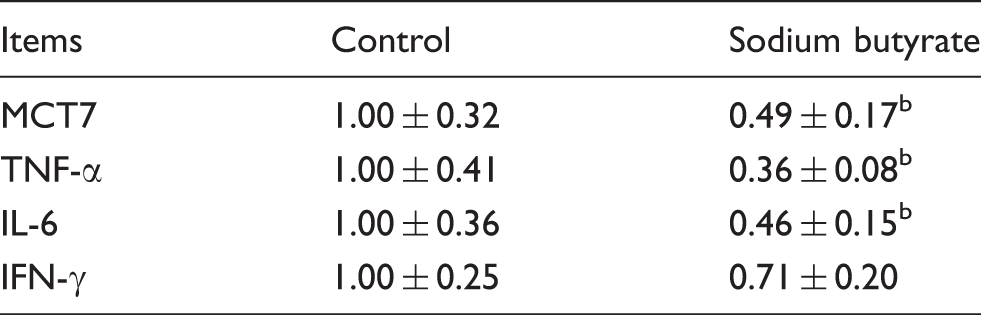
The 2-ΔΔCt method was used to analyze the relative expression (fold changes), calculated relative to the control group. Values are mean ± SD (n = 6).
Differences were considered significant at P < 0.05.
MAPK signaling pathways
Figure 1 shows the effects of sodium butyrate on the three MAPK signaling pathways (JNK, p38, ERK). In comparison with the control, supplemental sodium butyrate significantly decreased the phosphorylated ratio of JNK (p-JNK/JNK) (P< 0.05), whereas it did not significantly affect the phosphorylated ratios of ERK and p38 (p-ERK/ERK and p-p38/p38) (P > 0.05).
Discussion
Growth retardation is a common problem in weaned piglets. The present result, that dietary addition of sodium butyrate increased daily gain in weaned pigs, is consistent with previous research. 28 Moreover, sodium butyrate increased villus height and villus height:crypt depth ratio, which is consistent with the result of Lu et al. 29 The gastrointestinal tract is not only fundamental to the uptake of nutrients, but it also acts as a physical barrier. Disruption of the intestinal barrier facilitates luminal antigens to penetrate sub-epithelial tissues, resulting in a mucosal and systemic inflammatory response.3,30 Therefore, we evaluated the effect of sodium butyrate on the intestinal barrier of weaned piglets, using the Ussing chamber technique. A decreased TER and increased FD4 flux reflect increased paracellular permeability and impaired intestinal barrier. The present study showed that pigs fed with sodium butyrate showed higher TER and lower FD4 flux in jejunum, which indicated an improvement in intestinal barrier function. Similarly, using the lactulose to mannitol differential absorption test, Huang et al. 31 reported that sodium butyrate (1g/kg body mass) decreased the intestinal permeability of weaned piglets. The present study demonstrates that supplemental sodium butyrate improves growth performance, ameliorates weaning-associated intestinal injury and enhances intestinal barrier function.
Weaning-associated intestinal inflammation has negative effects on intestinal integrity and epithelial function in piglets.4,32 It has been reported that mast cell activation and inflammatory mediator release play a major role in the intestinal barrier dysfunction during the post-weaning period. 1 Mast cells are considered to be an important cell type and play central roles in mediating intestinal inflammation. 33 Upon activation, preformed mediators stored in granules are released immediately, resulting in increase of intestinal permeability.34,35 So far, only a few studies in vitro have reported that butyrate inhibits mast cell degranulation.18,36 This study, for the first time, demonstrated that dietary supplementation with sodium butyrate decreased the percentage of degranulated mast cell in weaned pigs. In addition, sodium butyrate reduced the content of mast cell inflammatory mediators in mucosa of weaned pigs, including histamine, tryptase, TNF-α and IL-6. These results are supported by previous observations in murine mast cell line CPII clone 12, where butyrate inhibited both degranulation and TNF-α release. 36 However, in mouse bone marrow-derived mast cells, Zhang et al. 19 found that butyrate suppresses FcɛRI-dependent cytokine release but had no effect on degranulation. The discrepancies might be related to the difference in cell models. The present study shows that dietary supplementation with sodium butyrate inhibited mast cell activation and release of its inflammatory mediators, thereby maintaining intestinal barrier function.
Several studies have shown that pro-inflammatory cytokines have negative effects on intestinal integrity and epithelial function.27,37 Controlling the release of intestinal pro-inflammatory cytokines may have potential benefits in alleviating gut disorders. 38 Previous studies reported that butyrate inhibited the inflammatory cytokines mRNA levels in mast cell in vitro.18,19 Therefore, we further investigated the effect of sodium butyrate on the relative mRNA expression of mast cell inflammatory mediators in the jejunal mucosa of weaned pigs. The results showed that dietary addition of sodium butyrate decreased the mRNA levels of mast cell-specific tryptase (MCT7), TNF-α and IL-6 in jejunal mucosa. It could be suggested that sodium butyrate improves intestinal integrity partially by inhibiting the mRNA expression of mast cell mediators in weaned pigs.
It has been reported that the MAPK signaling pathways regulate cytokine production in activated mast cells.23,39 To our knowledge, only a few studies in cell models in vitro have investigated the influence of sodium butyrate on MAPK signaling pathways.19,36 Diakos et al. 36 reported in murine mast cell line CPII clone 12 that butyrate treatment inhibited phosphorylation of JNK but not of p38 or ERK after dinitrophenyl-albumin challenge. In contrast to previous results, Zhang et al. 19 reported that sodium butyrate simultaneously inhibited the three MAPK signaling pathways in murine bone marrow-derived mast cells after trinitrophenol BSA stimulation. However, there has been no study regarding the effects of sodium butyrate on MAPK signaling pathways in vivo. In the present study, for the first time, we found that supplemental sodium butyrate significantly decreased the relative protein levels for phosphorylated JNK but did not affect phosphorylated ERK and p38 of jejunum mucosa in weaned pigs. Taken together, it is likely that the role of sodium butyrate in improving intestinal barrier function of weaned pigs is related to inhibition of JNK signaling pathway. However, the exact role of this signaling pathway remains to be elucidated. In a follow-up study, the specific inhibitors of JNK signaling pathway may be used to investigate whether inhibition of JNK can affect the beneficial role of sodium butyrate on mast cell degranulation and the intestinal barrier of weaned piglets.
In summary, dietary supplementation with sodium butyrate ameliorated weaning-associated growth retardation and intestinal injury, and inhibited mast cell activation and production of inflammatory mediators. The JNK signaling pathway is involved in the beneficial role of sodium butyrate on mast cell degranulation and intestinal barrier of weaned piglets.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by National Key R & D Program (2016YFD0501210), Zhejiang Province Key R & D Project (2015C02022), Zhejiang Province Major Science and Technology Project (2015C03006), Special Fund for Agro-scientific Research in the Public Interest (201403047), Dabeinong Funds for Discipline Development and Talent Training in Zhejiang University.