
Abstract
Exclusive enteral nutrition is established as an initial therapy to induce remission in active Crohn’s disease (CD), especially in children, but the mechanisms of action of this therapy are yet to be fully defined. Intestinal alkaline phosphatase (IAP), a recognised marker of enterocyte differentiation, is implicated in the innate gut immune response to enteric pathogens. Using the Caco-2 human adenocarcinoma cell line, this study showed that the incubation of human cells with a polymeric formula (PF) resulted in a dose-dependent increase in the expression of IAP on the cell surface. While further investigation is required to determine the pathway(s) involved, this finding suggests that cell surface-associated IAP may be an aspect of the gut’s innate immune response to pathogenic bacteria that is strengthened by PF in the setting of CD.
Keywords
Introduction
Crohn’s disease (CD) is a chronic condition involving the gastrointestinal tract that is characterised by an abnormal immune response to bacterial epitopes, resulting in inflammation and altered barrier function.1,2 These changes result in variable symptoms, with onset of disease from any age. 3 Currently, none of the available treatment approaches are curative.
Exclusive enteral nutrition (EEN), a liquid diet using elemental or polymeric formulae (PF), is increasingly accepted as a means to induce remission of active CD in adults and in children.4,5 In addition to reduction of disease activity and resolution of symptoms, EEN enhances mucosal healing, improves nutrition and avoids drug-related side effects. 6 However, while the clinical benefits of EEN are established, 7 the mechanisms of action of nutritional therapy are yet to be fully defined. This is important, because better understanding will lead to improved outcomes for patients that may include maintenance of remission. 8
Treatment of cultured colonic epithelial cells with PF (as used in EEN) is associated with increased anti-inflammatory activity and improved gut barrier function.9,10 PF also enhances the production of a carcinoembryonic antigen cell adhesion molecule (CEACAM6) on the surface of Caco-2 cells, which thereby bolsters innate immune responses by blocking the interaction of pathogenic bacteria with the intestinal epithelial barrier. 11 CEACAM6 is considered a marker of differentiated enterocytes, 12 and evidence that cells at the tips of the villi are less likely to be infected than undifferentiated cells in the crypts (despite a much greater exposure to bacteria) 13 supports the idea that differentiated cells contribute to innate immunity in the gut. Collectively, these findings suggest that PF may contribute to innate immunity by driving enterocyte differentiation. If so, such a finding is likely to be significant, given that a primary function of the epithelial cells that line the gastrointestinal tract is to provide a physical barrier that allows nutrient uptake and water absorption while keeping luminal bacteria out.
Enterocyte differentiation is associated with increased secretion of intestinal alkaline phosphatase (IAP). 14 IAP also contributes to gut mucosal defence by detoxification of LPS and reduced bacterial invasion.15,16 IAP is enriched on vesicles released from the tips of the microvilli. 17 As such, it is considered to act as a ‘secreted barrier’ in the gut lumen, where it is involved in the maintenance of normal gut microbial homeostasis. 18 The main aim of this study was to test the hypothesis that PF treatment of intestinal epithelial cells drives the proliferation and subsequent differentiation of enterocytes using IAP as a marker. A secondary aim was to determine if the effect of PF is mediated via the vitamin D receptor (VDR), based on evidence that this receptor is involved in maintaining the integrity of the intestinal epithelial barrier, 19 and that vitamin D can up-regulate the expression of IAP in these cells. 20
Materials and methods
Materials
Sodium butyrate (hereafter referred to as butyrate) and 1α,25-dihydroxyvitamin D3 [1,25(OH)2D3], the active form of vitamin D, which binds to the vitamin D receptor, 20 were sourced from Sigma (St Louis, MO, USA). PF (Osmolite) was sourced from Abbott Nutrition (Christchurch, New Zealand) and used at 20% (v/v) as previously described.9,10 Dulbecco’s PBS (D-PBS), Minimal Essential Medium (MEM) non-essential amino acids, penicillin/streptomycin and TrypLE™ Express were all obtained from Life Technologies (Auckland, New Zealand). Phosphatase substrate (p-nitrophenylphosphate; pNPP) and phosphatase standard (4-nitrophenol; pNP) were purchased from Sigma. Primary Abs were from Abcam (Melbourne, Australia; mouse anti-human IAP) and Sigma (mouse anti-GAPDH). The secondary Abs (HRP goat anti-mouse and FITC sheep anti-mouse) were from Pierce, Thermo Fisher Scientific and Silenius (Australia), respectively. BSA (AlbuMAX I Lipid rich BSA) was from Gibco (Invitrogen, Auckland, New Zealand). The VDR inhibitor TEI-9647 was synthesized at the Teijin Institute for Biomedical Research, Japan. All other reagents were from Sigma.
Cell culture
The Caco-2 (ATCC HTB-37) cell line used in this study was derived from a human colon adenocarcinoma and is widely used to model differentiating gut enterocytes.21,22 The cells were routinely cultured in MEM with Earle’s Salt and l-glutamine, supplemented with 10% (v/v) heat-inactivated FBS (Gibco), 0.1 mM MEM non-essential amino acids and 1% penicillin/streptomycin (all from Gibco). Cell culture was at 37℃ in a humidified atmosphere of 5% CO2 in air.
Cell viability
Caco-2 cells (1 × 105) were cultured in 24-well plates for 6–8 d [with and without the addition of 20% (v/v) PF at d 5]. The culture medium was removed and the remaining adherent cells were rinsed three times with D-PBS, with each wash added to the saved medium. The adherent cells were detached with TrypLE™ Express, the well washed with D-PBS, with both added to the pooled medium. A 1.5-ml aliquot of the final cell suspension was centrifuged at 500 g for 5 min and the supernatant discarded. The remaining pellet of cells was suspended in 150 µl of medium, an equal volume of trypan blue (in D-PBS, 0.025% final concentration) was added and the cells were loaded onto a haemocytometer (BLAU Brand, Germany). Cells were counted under the 40 × objective of a light microscope (Nikon, Tokyo, Japan) and viability was expressed as the number of trypan blue-negative (live) cells.
IAP activity
IAP activity was determined by measuring the change in absorbance as pNPP was hydrolysed to form yellow pNP. 14 Caco-2 cells were washed once with 1 ml PBS before being scraped into 300 μl cold collection buffer (0.1 M Tris-HCl, 150 mM NaCl; pH 8.0). Following vigorous pipetting to disperse cell fragments, 150 μl pNPP substrate solution [2.5 g/l pNPP (Sigma), 100 mM diethanolamine, 150 mM NaCl, 2 mM MgCl2; pH 9.5] was added to 50 µl cell lysate. After 8 min, 50 μl 0.5 M NaOH was added to the test samples to stop the reaction. The tubes were centrifuged for 3 min at 4000 g to pellet cell debris. Supernatant (200 μl) was removed and added to separate wells in a 96-well plate. Absorbance was read at 405 nm using a Spectramax-190 plate reader (Molecular Devices, Sunnyvale, CA, USA) and Softmax Pro V5.3 software (Molecular Devices) and converted to concentration after blank subtraction (duplicate sample incubated with 50 μl 0.5 M NaOH and 150 µl substrate solution without pNPP) with reference to a standard curve [50 µl pNP concentrations (0–700 µM) added to 150 µl pNPP solution and 50 µl NaOH 0.5 M]. Each assay included samples from duplicate wells and results were reported as pNP activity (µM).
Preparation of whole-cell lysates for immunoblotting
Caco-2 cells (1 × 105) were cultured in 24-well plates for 6–8 d [with and without the addition of 20% (v/v) PF at d 5]. Media was removed and the cells were washed once with PBS before being lifted with TrypLE™ Express. An equal volume of media was added to the cell suspension before it was centrifuged (800 g for 3 min). The resultant cell pellet was re-suspended in 100 μl RIPA buffer [50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% (v/v) NP-40, 1 × protease inhibitor (Complete Mini; Roche, Basel, Switzerland), 2% SDS, 2% mercaptoethanol, 5% glycerol v/v, 0.01% (w/v) bromophenol blue]. The samples were placed on ice for 30 min before centrifugation for 20 min at 30,000 g, to remove remaining cellular debris. The whole-cell lysate was stored at –20℃.
Immunoblot analysis of IAP expression in Caco-2 cells
SDS-PAGE-separated Caco-2 cell lysates were transferred to a polyvinylidene fluoride membrane (Bio-Rad, Hercules, CA, USA) by electroblottting and non-specific binding sites were blocked [5% (w/v) BSA in Tris-buffered saline with 0.05% Tween 20 (TBS-T)] for 1 h, at 25℃ (room temperature). The membrane was then probed overnight (18 h) at 4℃ with a mouse monoclonal anti-human IAP Ab (diluted 1/500), followed by HRP-conjugated goat anti-mouse IgG (diluted 1:5000) as secondary Ab. The Abs were diluted in 2% (w/v) BSA in TBS-T. The membrane was developed with a chemiluminescent substrate (ECL Plus Pierce Blotting Substrate; Thermo Fisher Scientific) and the blot photographed using a UVITEC Cambridge Gel Documentation system (UVITEC, Cambridge, UK). Blots were stripped and re-probed with GAPDH Ab. For quantitative analysis, bands were detected and evaluated densitometrically using UVI band software (UVITEC), normalised for the density of GAPDH. 20
Fluorescence microscopy
Caco-2 cells (1 × 105) were grown on ethanol-sterilised 16-mm round coverslips for 5 d. The medium was replaced, with and without the addition of 20% (v/v) PF, and the cells were cultured for up to 72 h longer. The supernatant was removed and the cells were fixed by adding 4% w/v paraformaldehyde for 45 min. Following two 5-min washes in PIPES buffer (60 mM PIPES; pH 6.8), potential non-specific binding sites on the cells were blocked by incubating the coverslips in blocking buffer [5% BSA (w/v) in PIPES] for 45 min at room temperature before overnight incubation at 4℃ in mouse anti-human IAP Ab, diluted in PIPES containing 2.5% (w/v) BSA, followed by a FITC-conjugated sheep anti-mouse secondary Ab for 2 h at room temperature [also diluted in PIPES buffer containing 2.5% (w/v) BSA]. Cell nuclei were stained with Hoechst 33342 (10 µg/ml, in D-PBS) at room temperature for 20 min. After four 10-min washes [PIPES containing 0.05% (v/v) Tween-20], followed by a final 10-min PIPES buffer only wash, the coverslips were mounted in p-phenylenediamine [0.1% (w/v) in 50 mM Tris-HCl; pH 6.8] and examined using epifluorescent microscopy on an AxioImager Z1 upright microscope with a 20 × EC Plan Neofluoar lens (Zeiss, Oberkochen, Germany). Images were taken with an Axiocam MRm camera, coupled to AxioVision software (Zeiss).
Statistical analysis
Results are the means ± SE of the means. One-way or two-way repeated measures ANOVA were used for analysis of the data sets. If the P-value was < 0.05, the ANOVA was followed by Tukey’s multiple comparisons post hoc test. Analysis was performed using Prism 6 (GraphPad Software Inc., La Jolla, CA, USA).
Results
Cell density
Caco-2 cell proliferation increased over 7 d of culture, with cell numbers significantly higher at all subsequent time points when compared with those at d 3 (P < 0.0001; Figure 1a). However, the rate of proliferation was observed to slow by d 5, and cell numbers plateaued at approximately 5 × 105 cells per well. This likely reflects contact inhibition as the cells start to form a confluent monolayer,
22
because the decrease in cell proliferation was not associated with an increase in the percentage of non-viable cells (Figure 1b). Together, these results [which are the mean ± SE of (a) three and (b) five independent experiments] informed the choice of using Caco-2 cells grown for 5 d as the time point to add PF.
Caco-2 cell numbers increase over time. (a) Trypan blue was added to Caco-2 cells before counting by haemocytometer with (b) the percentage of dead cells determined by trypan blue exclusion. Results are the mean ± SE of (a) three and (b) five independent experiments. Raw data were analysed by two-way repeated measures ANOVA. No significant differences were detected between the two concentrations of cells at any specific time point.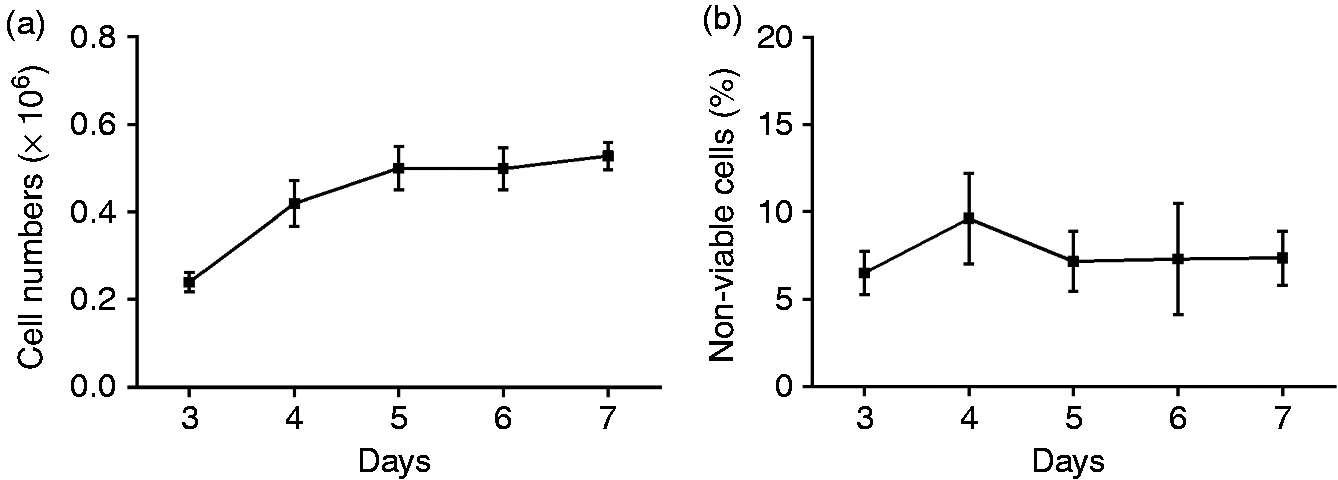
PF causes a decrease in cell number
The addition of 20% (v/v) PF to 5-d-old cultures of Caco-2 cells was found to have a marked effect on cell number over time (Figure 2a). Eight independent experiments revealed that cell numbers were found to decrease in the PF-treated wells, from 0.39 ± 0.05 at 24 h to 0.33 ± 0.02 at 72 h. In contrast, untreated cells continued to proliferate over the same period of time, increasing from 0.49 ± 0.04 to 0.67 ± 0.05. This difference was significant (P < 0.01) and, consequently, cell numbers were significantly higher in control wells at 48 and 72 h.
PF treatment increases cellular membrane associated IAP. Caco-2 cells were incubated with 20% (v/v) PF for up to 3 d. Cell (a) proliferation and (b) viability, and (c) IAP activity (in cell homogenates) were measured at 24 h intervals. Results are the mean ± SE of (a, b) eight and (c) three independent experiments. Raw data were analysed by two-way repeated measures ANOVA. Asterisks indicate a significant difference between the two cell treatments at fixed times. **P < 0.01, ***P < 0.001; ****P < 0.0001.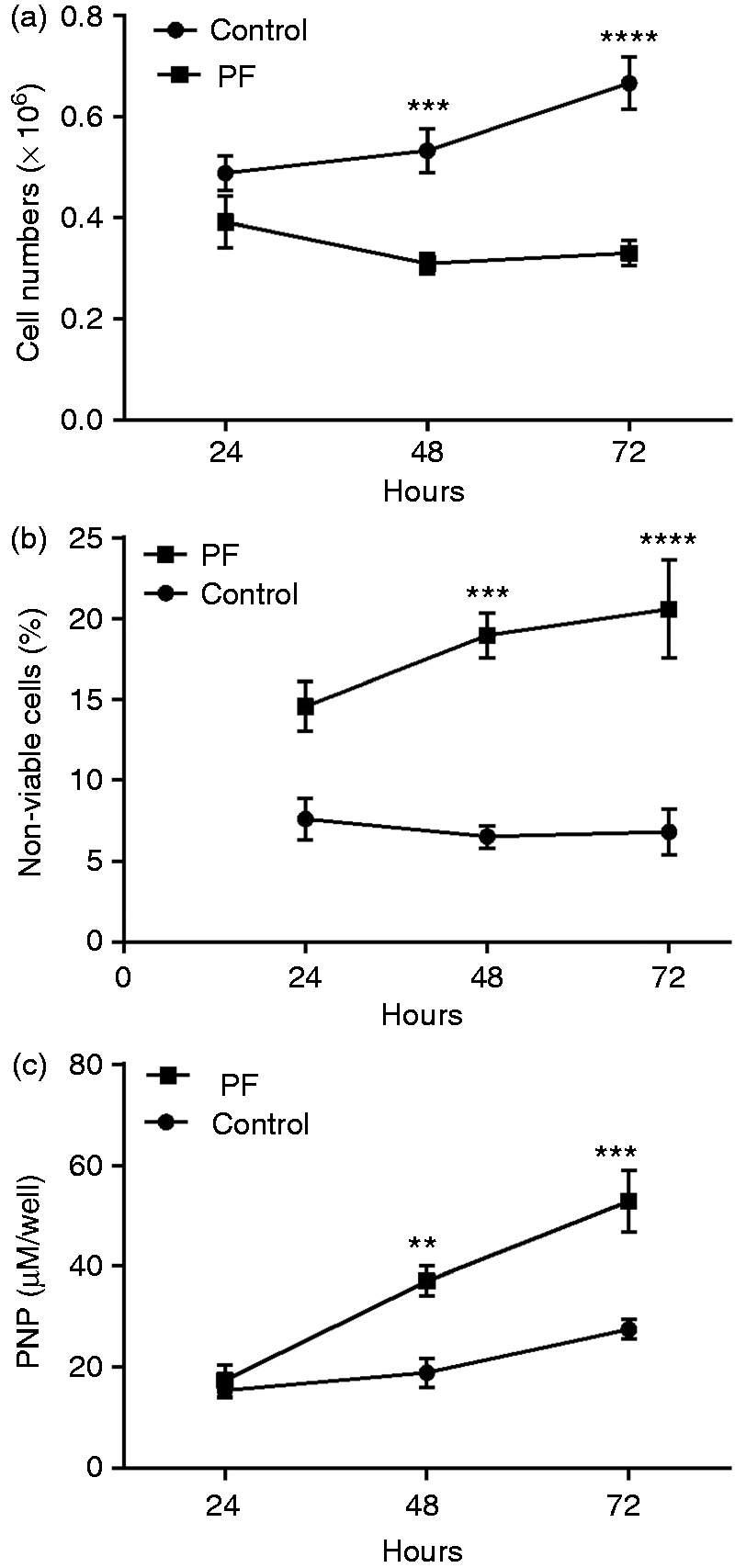
PF is associated with loss of cell viability
The observed decrease in Caco-2 cell numbers following the treatment of 5-d-old cultures with 20% (v/v) PF was explored further by determining the number of non-viable cells (assessed by trypan blue staining) in the same untreated and PF-treated cells (n = 8 independent experiments). Cell death in the untreated cells ranged from 6% to 8% and did not increase over time (Figure 2b). In contrast, treatment of the cells with 20% PF resulted in a significant increase in cell death over time (P < 0.05). Consequently, cell death was significantly higher in PF-treated wells at 48 h and 72 h.
PF is associated with increasing IAP activity
IAP activity is considered a marker of epithelial cell differentiation. 14 Levels of cellular IAP were measured across three independent experiments to determine if the observed effect of 20% (v/v) PF on cell growth was associated with enhanced differentiation as cell proliferation decreased. Whereas IAP activity in the untreated cells did not increase over time, treatment with 20% (v/v) PF caused a significant increase in Caco-2 cell IAP activity over time (P < 0.0001; Figure 2c). Consequently, IAP expression was significantly higher in PF-treated compared with untreated wells at 48 h and 72 h.
PF increases membrane expression of IAP
Lysates of 5-d-old Caco-2 cell monolayers incubated with 20% (v/v) PF for 48 and 72 h were probed with a monoclonal Ab against IAP and the density of IAP staining determined relative to the staining intensity of GAPDH in the same sample. PF treatment was observed to increase cell-associated IAP staining at both time points (1.361 ± 0.487 and 1.901 ± 0.509 at 48 h and 72 h, respectively) when compared with untreated controls (0.710 ± 0.111; Figure 3). However, analysis of the raw data from three independent experiments did not detect a significant increase in IAP staining at either time point.
PF causes an increase in IAP protein expression. Cells incubated with 20% (v/v) PF for 48 h and 72 h were lysed in RIPA buffer and probed for Abs specific for IAP. The blot was then re-probed with an anti-GAPDH Ab. Images are representative of three independent experiments.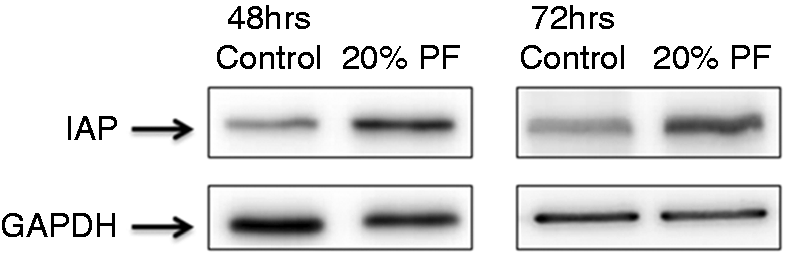
Caco-2 cells grown on coverslips for 5 d were given fresh medium with increasing concentrations (0, 5%, 10% and 20%) of PF for up to 72 h. Fluorescence microscopy of the fixed but not permeabilised cells following incubation with a monoclonal Ab against IAP revealed a marked difference in IAP protein expression between untreated and PF-treated cells that extended to a concentration-dependent effect of PF (Figure 4, which is representative of two independent experiments). In untreated Caco-2 cells, membrane-associated IAP expression was seen to be discrete and not evenly distributed (Figure 4a). With each increase in PF concentration, the IAP staining became more widespread and diffuse (Figure 4b–d). In contrast, PF-treated cells incubated without primary Ab showed no detectable fluorescence (not shown). Collectively, these findings suggested that PF has a demonstrable effect on cell differentiation. However, the mechanism(s) responsible for this effect remain unknown.
PF increases membrane-associated IAP. Caco-2 cells were (a) untreated, or treated with (b) 5%, (c) 10% or (d) 20% PF for 72 h before being stained for membrane-associated IAP. Magnification 20 × . Images are representative of two independent experiments.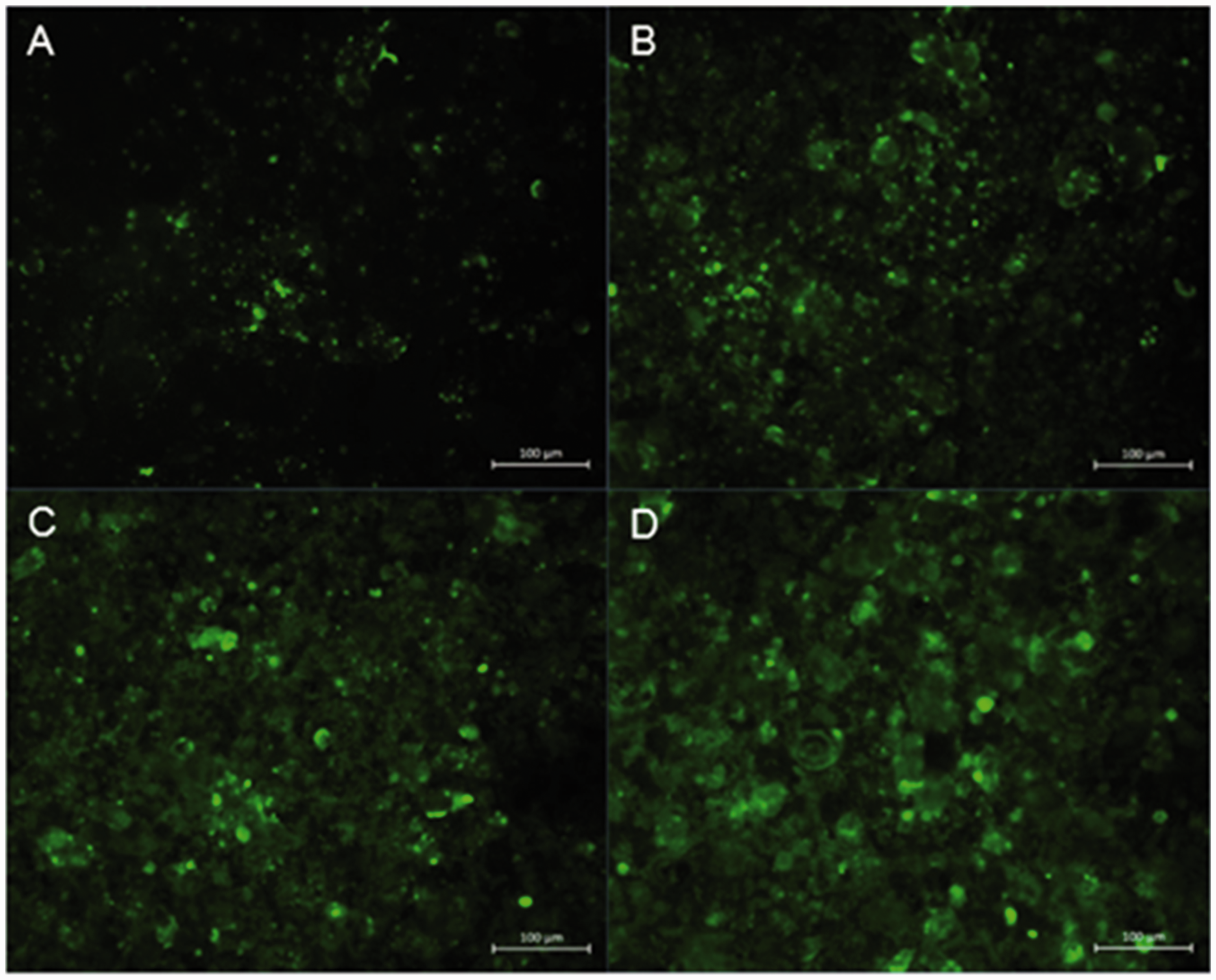
Butyrate and vitamin D increase IAP activity
Butyrate and 1,25(OH)2D3, an active form of vitamin D, were used as positive controls to test the hypothesis that the observed PF-associated effect on cell differentiation is mediated through the VDR.20,23 The addition of butyrate or 1,25(OH)2D3 to 5-d-old Caco-2 cell monolayers for 72 h induced IAP activity (Figure 5). The findings of three independent experiments revealed that butyrate had a significant dose-dependent effect on IAP activity (P < 0.05) and the highest concentration of butyrate (8 mM) induced significantly more IAP activity (3.4 μM of pNP per million cells) than 20% (v/v) PF (1.3 μM of pNP per million cells; P < 0.05). In contrast, 1,25(OH)2D3 had noticeably less effect on IAP activity, despite being added in concentrations that reportedly activate the VDR in cultured cell lines.
19
Dose-dependent effect of 1,25(OH)2D3 and butyrate on IAP activity. Caco-2 cells were exposed for 72 h and activity was measured in the cell lysate. PF [20% (v/v)] was used as a positive control. Results are ± SEM of three independent experiments. Raw data were analysed by repeated-measure ANOVA, and asterisks linked by bars indicate significant differences between groups. *P < 0.05.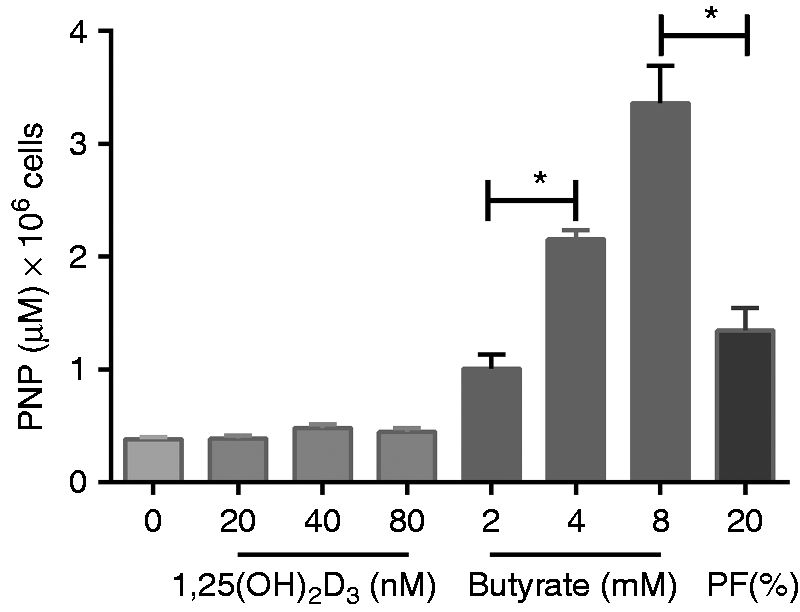
Despite inducing the highest levels of IAP activity, microscopy suggested that 8 mM butyrate was cytotoxic (not shown). However, the effect of 4 mM butyrate on the cells, assessed by counting cell numbers (0.17 ± 0.02 × 106) and viability (17.4% ± 3.7 non-viable cells) was found to be similar to the effect of 20% (v/v) PF added to Caco-2 cells for the same period of time (0.21 ± 0.04 × 106 cells and 21.3% ± 1.9 non-viable cells, respectively). Accordingly, 4 mM butyrate was used as a positive control for subsequent experiments where IAP release by 5-d-old monolayers of Caco-2 cells was assessed, with and without the addition of a VDR inhibitor (TEI-9647).
IAP-dependent effect of PF does not signal through the VDR
TEI-9647 inhibitor was provided at a concentration of 1 mM dissolved in ethanol with a recommended dilution of 1:1000 for cell culture experiments (Teijin Ltd, Japan). Here it was used at half (0.5 µM) and at double (2 µM) this concentration, and controls included cells exposed to a 1/500 dilution of ethanol in cell culture media, using the same experimental parameters across three independent experiments. Cell numbers and viability in these ethanol controls did not differ from untreated cells (not shown).
Treatment of Caco-2 cells with TEI-9647 was associated with a dose-dependent decrease in IAP activity in 4 mM butyrate and 80 nM 1,25(OH)2D3-stimulated cells (P < 0.05), with this effect most notable in the presence of butyrate (Figure 6a). In contrast, the VDR inhibitor (VDRI) had no effect on IAP levels in Caco-2 cells following exposure to 20% (v/v) PF (Figure 6a). 1,25(OH)2D3, butyrate and PF treatment were each observed to cause changes in Caco-2 proliferation and viability, with butyrate- and PF-treatment associated with a noticeable decrease (Figure 6b). The VDRI (used at 2 µM) was associated with a 50% reduction in butyrate-treated cell numbers (P < 0.01). A similar effect was not seen in the PF-treated cells co-incubated with the VDRI (Figure 6b). 1,25(OH)2D3 (80 nM) treatment had no effect on cell viability, whereas 4 mM butyrate and 20% (v/v) treatment each increased cell death to 16% and 21%, respectively. The PF-mediated increase in cell death was significant when compared with untreated controls (P < 0.01; Figure 6c). Addition of the VDRI had no apparent effect on the rate of cell death in untreated cells or cell treated with either 20% (v/v) PF or 80 nM 1,25(OH)2D3. However, increasing concentrations of TEI-9647 added to 4 mM butyrate treated cells notably reduced cell death, which is consistent with butyrate-induced cell death being mediated through the VDR (Figure 6c).
Effect of TEI-9647 on Caco-2 cell responses. Caco-2 cells were exposed for 72 h to 80 nM 1,25(OH)2D3, 4 mM butyrate and 20% (v/v) PF, without and with the addition of the VDRI at 0.5 µM and 2 µM. (a) IAP activity measured in the cell lysate, (b) cell numbers and (c) cell viability (determined by trypan blue exclusion). Results are ± SEM of three independent experiments. Raw data for each treatment group were analysed separately by repeated-measure ANOVA, and asterisks linked by bars indicate significant differences between the different concentrations of inhibitor within each group. *P < 0.05, **P < 0.01.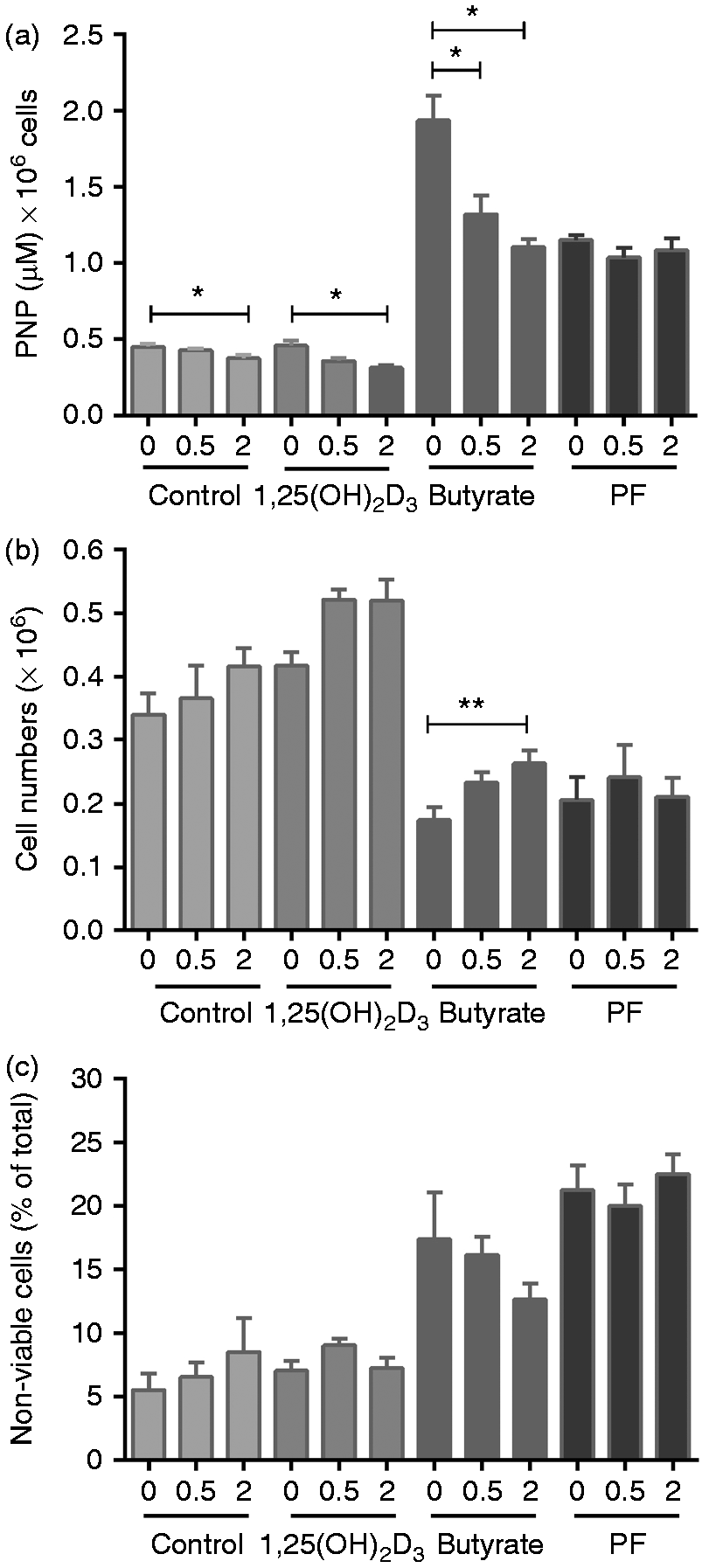
Discussion
Marked changes in phenotype are observed as intestinal epithelial cells move from the proliferative crypt toward the luminal interface where, as differentiated cells, they perform protective and absorptive roles. Differentiation is associated with the formation of mature tight junctions that help maintain barrier function, and is signalled by an increase in IAP activity. 23 Accordingly, the observation of lower-than-normal levels of IAP protein in the inflamed gut mucosa of children with CD suggests the normal processes required for enterocyte differentiation may be impaired. 24
PF enhanced the rate of Caco-2 cell proliferation and, as proliferation slowed, IAP levels increased in the treated cells, a finding that is consistent with reports of a role for PF in promoting and maintaining barrier function in the gut. 10 IAP also acts as a host defence protein by detoxifying bacterial products and regulating epithelial–microbial interactions at the gut wall. 17 Thus, a PF-associated increase in IAP expression would bolster this response. IAP is not the only protein released from the luminal surface of the differentiated cells that line the gut wall. Incubation of Caco-2 cells with 20% (v/v) PF doubles the release of CEACAM6 from the cell surface, reducing the ability of adherent-invasive Escherichia coli to associate with the gut epithelium. 11 Antimicrobial peptides that include members of the cathelicidin and defensin families also play a key role in host protection at this site. β-Defensin expression occurs in response to infection and inflammation, 25 whereas production of the bioactive cathelicidin peptide LL-37 is linked to enterocyte differentiation. 26 However, whether LL-37 expression in the small bowel increases in response to PF treatment remains to be determined. Evidence that differentiated enterocytes are less likely to become infected with invasive bacteria or become leaky to bacterial antigens in the presence of host inflammation also further supports the hypothesis that PF acts to enhance the innate immune response, 13 thereby protecting the gut from adherent and/or invasive bacteria.
The component(s) in PF that drive enterocyte differentiation with concomitant IAP expression are currently unknown, but one promising candidate was considered to be vitamin D. Low vitamin D and VDR levels are reported in patients with inflammatory bowel disease, 27 whereas long-term vitamin D supplementation is associated with fewer symptoms. 28 This positive effect associated with vitamin D may reflect enhanced barrier function, 29 as well as increased expression of IAP and cathelicidin.20,30 Collectively these findings strengthen the hypothesis that vitamin D may mediate the anti-inflammatory and/or pro-differentiation effects associated with enteral nutrition.
Almost all known actions of vitamin D are mediated through the VDR. 31 Butyrate, a short-chain fatty acid produced by colonic bacteria and considered to provide an important energy source for colonocytes, also signals through this receptor.20,32 Exposure of Caco-2 cells to butyrate also results in a dose-dependent increase in IAP activity, CEA protein concentration, cathelicidin expression and barrier function are related to enhanced crypt-to-villus differentiation.23,26 Taken together, these observations support the hypothesis that PF most likely also acts through the VDR.
A VDRI was used to test this hypothesis. TEI-9647 has been shown to antagonise the VDR in human osteoclasts and monocytes.33,34 However, there is no evidence to date regarding the ability of TEI-9647 to inhibit the VDR in Caco-2 cells. IAP expression, which increased following treatment of Caco-2 cells with butyrate or 1,25(OH)2D3, significantly decreased over time with the addition of TEI-9647. This supports earlier studies that find involvement of the vitamin D signalling pathway in colonic epithelial cell differentiation by butyrate.20,32 The response to the inhibitor was dose-dependent, and the effect was most notable when the higher concentration of TEI-9647 was added to cells stimulated with 4 mM butyrate, with IAP levels falling by approximately half. In marked contrast, the effect of 20% PF on IAP levels was not affected at either VDRI concentration.
Likewise, treatment with TEI-9647 blocked the butyrate-mediated effect on cell growth, and this effect was observed to be concentration-dependent. In contrast, VDRI treatment of Caco-2 cells did not override the inhibitory effect of 20% PF on cell proliferation. These findings suggest a number of things. Firstly, the shorter side chains may reduce the sensitivity of TEI-9647 as a VDR antagonist, 34 suggesting it is possible that TEI-9647, at the concentration used, failed to block PF-binding to the VDR. This remains to be tested. However, it is also possible that given the number of ingredients in PF, signalling may be via different receptor(s) and/or mechanisms. For example, EEN formulations that include eicosapentaenoic acid and docosahexaenoic acid are sources of resolvins that also induce IAP in Caco-2 cells, albeit via a different (ChemR23) receptor. 35 In contrast (or in addition), the effect of PF could also be mediated either through some calcium-dependent channel and/or through an independent calcium-sensing mechanism. This remains to be determined.
In summary, measurement of IAP activity as a marker of enterocyte differentiation revealed that protein expression and enzyme activity were increased in PF-treated Caco-2 cells. 1,25(OH)2D3 (an active form of vitamin D) and butyrate, both of which are known to promote enterocyte differentiation through the VDR, were used as positive controls to determine if PF also signals through this receptor. The VDRI used in the current study halved the effect of butyrate on cell proliferation and IAP activity in a dose-dependent fashion. The same inhibitor also caused a significant decrease in IAP activity in both 1,25(OH)2D3 treated and untreated cells. In contrast, the presence of TEI-9647 had no effect on PF-mediated cell proliferation and/or IAP activity, indicating that the accelerated differentiation of Caco-2 cells in the presence of PF is not primarily mediated through the VDR. Further work is now needed to translate these in vitro findings to the in vivo setting.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Canterbury Medical Research Foundation provided a grant-in-aid to JK to cover consumable costs for the study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.