
Abstract
Histiocytic ulcerative colitis (HUC) is a chronic enteropathy which most notably occurs in Boxer dogs and French bulldogs. The inflamed mucosa is hallmarked by large, foamy, periodic acid–Schiff (PAS)-positive macrophages infiltrating the colonic mucosa. As little is known about their origin and phenotype, an immunohistochemical study was performed using different macrophage markers. Generally, canine colonic macrophages showed high expression of ionised calcium-binding adaptor molecule 1 and MHC class II. In canine HUC, macrophages revealed up-regulation of lysozyme and L1 Ag but decreased CD163 expression compared with controls, suggesting them to be pro-inflammatory cells, whereas the healthy colonic mucosa was characterised by an anti-inflammatory macrophage phenotype. In addition, PAS reaction was used to discriminate macrophage subpopulations. PAS– macrophages displayed higher expression of L1 Ag and CD64, whereas PAS+ cells, which were only present in HUC patients, were characterised by increased expression of lysozyme, inducible nitric oxide synthase and CD204. This indicates PAS+ cells to be mature macrophages contributing to the inflammatory process, which are most likely maintained by differentiation of immature PAS– macrophages continuously recruited from blood monocytes. In summary, macrophage heterogeneity in canine HUC probably illustrates their different maturation states and functions compared with the healthy animals.
Introduction
Histiocytic ulcerative colitis (HUC) is a chronic enteropathy in dogs that is characterised by colonic lamina propria (LP) and submucosal infiltration with salient large periodic acid–Schiff (PAS)-positive (PAS+) macrophages.1,2 This entity is also known as granulomatous colitis and was initially described in Boxer dogs, suggesting a breed-specific disease in hereditarily susceptible dogs.2,3 However, HUC is also reported in other breeds, including French bulldogs,4–6 English bulldogs, 7 Mastiff, Alaskan Malamute, Doberman Pinscher 8 and Beagles. 9 Typically, young dogs under 4 yr are affected.1,2 Clinically apparent symptoms among dogs are mainly haematochezia, diarrhoea, mucous faeces, inappetence and mass loss.1,2 Although the aetiology has been unknown for a long time, a bacterial infection was assumed already in the first descriptions.2,10,11 More recently, Escherichia coli particles have been demonstrated in LP macrophages of HUC patients.6,12 In addition, dogs treated with antimicrobial drugs show eradication of E. coli and lasting clinical remission supporting a crucial involvement of E. coli in the pathogenesis of HUC.7,13
Grossly, the colon reveals focal, dot-shaped reddening in the early stage of HUC, which histopathologically corresponds to vacuolated surface epithelium followed by infiltration with macrophages and neutrophils. 14 Loss of epithelium and extension of the LP by abundant amounts of large macrophages with pink foamy cytoplasm extending into the submucosa represent the hallmark of advanced disease stages of HUC.2,11 Lesions are not consistently distributed as some areas show normal epithelium and LP while large numbers of characteristic macrophages expand the submucosa. 2 Histopathological demonstration of PAS+ material within the cytoplasm of these macrophages has been accepted as a pathognomonic feature of HUC.2,3 Histochemical and electron microscopical studies have been performed to elucidate the structure of the macrophages and the origin of the intracellular PAS+ material.10,11 Histochemically, large cytoplasmic globules are positive for fat stains and exhibit a strong autofluorescence, suggesting that they contain (phospho)lipids. 11 In addition, small granules show a strong affinity for nuclear dyes, such as methyl green–pyronin, and they are interpreted as DNA of organisms. 11 Moreover, the remaining intracytoplasmic PAS+ material is most consistent with mucopolysaccharides and is supposed to originate from degenerated products of organismal cell walls. 11 Ultrastructurally, most macrophages show distention with numerous, round, membrane-enclosed residual bodies interpreted as aged phagosomes, while some smaller macrophages additionally exhibit phagosomes containing bacteria. 10
In addition to studies investigating the intracytoplasmic content of macrophages, immunohistochemistry (IHC) has been used to elucidate the cellular immune response in HUC. 15 Besides the appearance of PAS+ macrophages, lesions are characterised by increased amounts of IgG+ plasma cells, CD3+ T cells, as well as increased expression of MHC class II (MHC II) and L1 Ag, compared with non-lesional areas of HUC patients and healthy controls. 15 Based on serial sections, MHC II but not L1 Ag expression is assumed for PAS+ macrophages, although this has not been substantiated by the use of double-labelling techniques until now. 15
Macrophages play an important role in intestinal immunity by both maintaining intestinal homeostasis and promoting protective immunity during inflammation. 16 In the healthy murine gut, macrophages display an anti-inflammatory phenotype with expression of surface molecules like MHC II and CD163, 17 as well as production of the anti-inflammatory cytokine IL-10.17,18 During inflammation, newly invaded blood monocytes differentiate into pro-inflammatory macrophages that are characterised, for example, by expression of inducible NO synthase (iNOS). 19
To further elucidate the phenotype of the unique macrophage population in HUC, the aim of this study was to perform double labelling with PAS reaction and IHC using different macrophage markers.
Material and methods
Case material
Detailed information about HUC cases and control dogs.
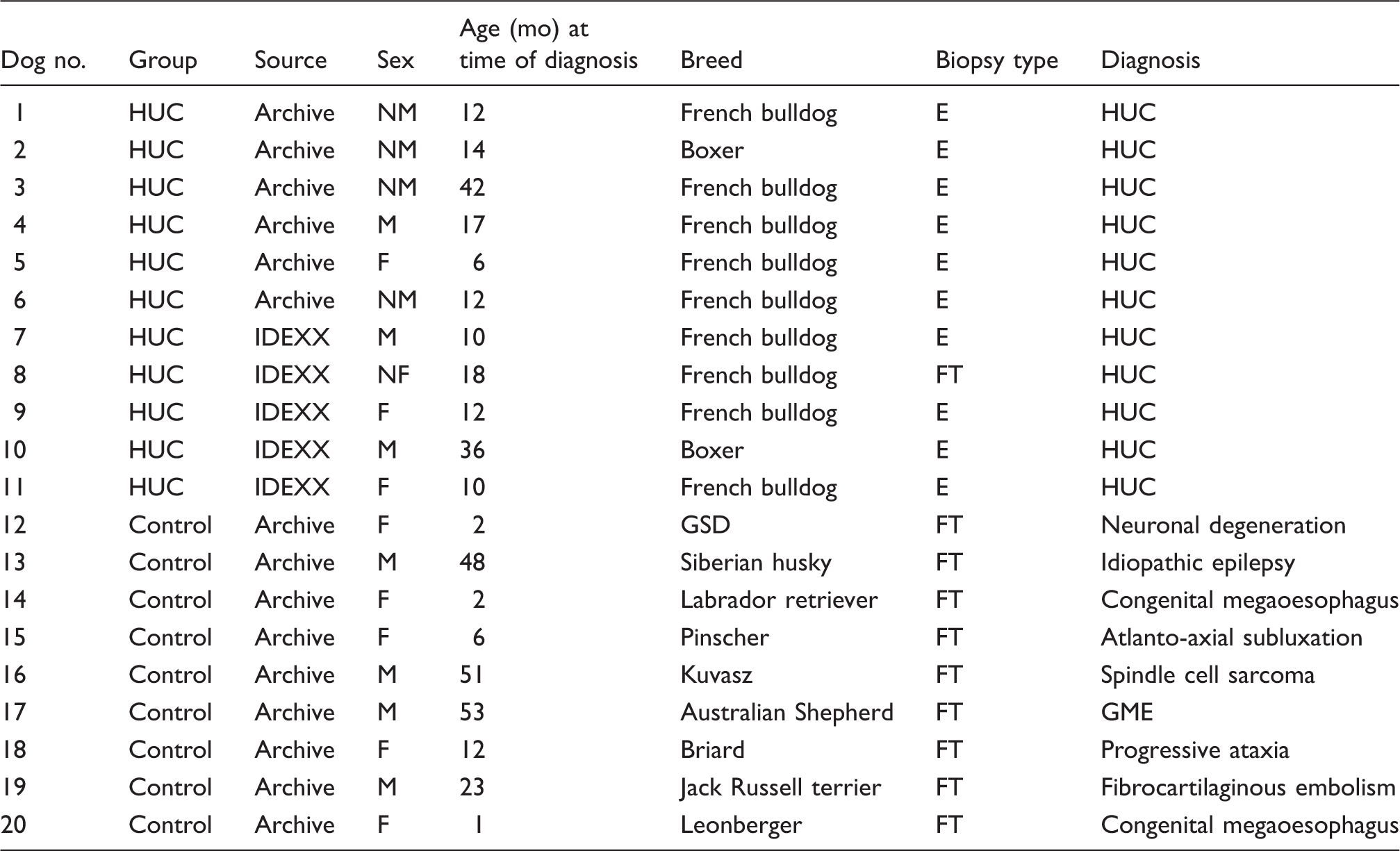
Archive refers to Archive of the Department of Pathology, University of Veterinary Medicine Hannover; IDEXX refers to the Archive of the Vet Med Labor GmbH, Division of IDEXX Laboratories Ludwigsburg.
NM: neutered male; E: endoscopic; M: male; NF: neutered female; FT: full thickness; F: female; GSD: German shepherd dog; GME: granulomatous mengingoencephalomyelitis
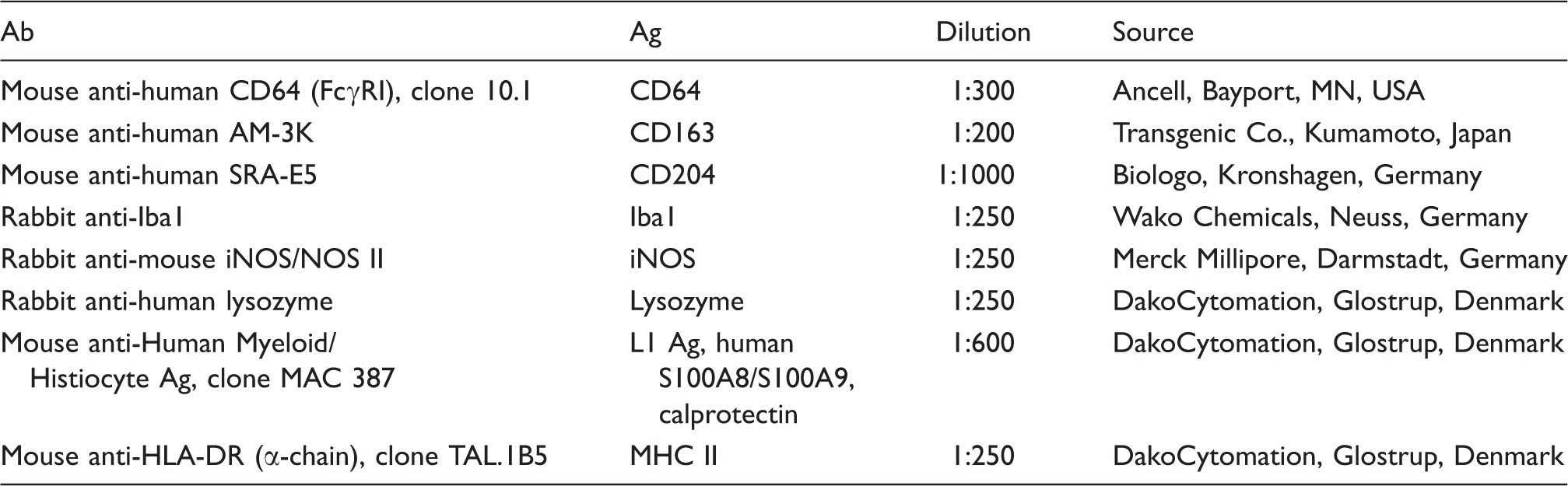
Histochemical and immunohistochemical procedures
Primary Abs used in the present study.
Evaluation of sections
Haematoxylin and eosin- and PAS-stained sections were evaluated in accordance with established guidelines. 23 To evaluate IHC, 10 images per section were taken at 400 × magnification from randomly selected areas using an Olympus BX-51 digital camera microscope [Olympus Optical Co. (Europe) GmbH, Hamburg, Germany] and cell-D imaging software (Olympus Soft imaging Solutions, Münster, Germany). This included representative amounts of all anatomical localisations present on the slides such as pericryptal, subepithelial and submucosal areas if available (n = 4). Afterwards, photographs were manually evaluated using the ImageJ-based imaging processing package Fiji (http://fiji.sc/Fiji). Only cells with a macrophage-like morphology characterised by a moderate-to-large, pleomorphic cell shape with plenty of cytoplasm and euchromatin-rich, central-to-paracentral, partly bean-shaped nucleus (designated as macrophages in the following) in the LP were counted. Thereby, cells with the following staining patterns were distinguished: cells solely displaying Ag expression (brown); exclusively PAS+ cells (pink); cells exhibiting both Ag and PAS staining (light brown to pink). The counting areas were measured with Fiji measurement tools and cell counts were expressed as cells per 150,000 µm2.
Statistical analysis
Statistical analyses were made using R version 3.1.2 (The R Foundation for Statistical Computing, Vienna, Austria). Initially, assessment for normal distribution of data was performed by means of the Shapiro–Wilk test. Subsequently, the Wilcoxon rank sum test was used to evaluate differences between cell counts in HUC cases and controls. Moreover, the macrophage subtypes in HUC were compared using the Wilcoxon signed rank test. Spearman's rank correlation coefficient (Spearman’s rho) was calculated to detect possible associations between the expression of CD64, CD163, CD204, iNOS, L1 Ag and lysozyme, as well as between ionised calcium-binding adaptor molecule (Iba)1 and MHC II in the PAS+ and PAS– macrophage subsets in HUC samples. For significant P-values, the Spearman’s rho was interpreted as follows: if the rho was between 0.00 and 0.19 it was defined as a ‘very weak correlation’; if between 0.20 and 0.39 as a ‘weak correlation’; if between 0.40 and 0.59 as a ‘moderate correlation’; if between 0.60 and 0.79 as a ‘strong correlation’; if between 0.80 and 1.00 as a ‘very strong correlation’. P-Values ≤ 0.05 were defined as statistically significant.
Results
Histopathology
In control animals, only mild changes, such as a slight increase in LP lymphocytes and plasma cells, minimal crypt distension and mild mucosal fibrosis were detected, all of which were interpreted as a background finding, as described previously.20,21 PAS+ cells similar to macrophages characteristic for HUC were absent in the colon of dogs without gastrointestinal disorders.
Evaluation of HUC cases displayed marked pathological changes that were predominated by massive infiltration of LP with an abundance of large pink macrophages with strongly PAS+ granular-to-foamy cytoplasm. In the majority of sections lesions were most prominent beneath basal crypts adjacent to the lamina muscularis mucosae showing ribbon-like formations of PAS+ macrophages. Macrophage infiltrations habitually separated crypts from each other and occasionally caused disruption of normal architecture with crypt loss. In biopsies of 4 of 11 cases (dogs 1, 2, 4 and 8) the submucosa was also assessable and contained large amounts of PAS+ macrophages often arranged in large clusters. Multifocal disruption and loss of surface epithelium with underlying infiltrates mainly of neutrophils and to a lesser extent of macrophages represented a consistent finding. In addition, the surface and crypt epithelium were characterised by a marked loss of goblet cells. Frequently, characteristic macrophage infiltrations were accompanied by smaller amounts of lymphocytes, plasma cells and neutrophils.
Colonic steady-state macrophages were characterised by expression of Iba1, MHC II, CD163 and CD204
Under homeostatic conditions (henceforth termed ‘steady state’), high numbers of colonic macrophages were characterised by expression of Iba1 and MHC II (Figure 1). Lower numbers of macrophages were also positive for CD163 and CD204. In contrast, only few macrophages were labelled for L1 Ag, while expression of CD64 and lysozyme was detected in single cells.
Expression of colonic macrophage markers in canine HUC and controls. Note significant differences regarding expression of lysozyme, L1 Ag and CD163. Wilcoxon rank sum test P-values: *P ≤ 0.05; ***P ≤ 0.001.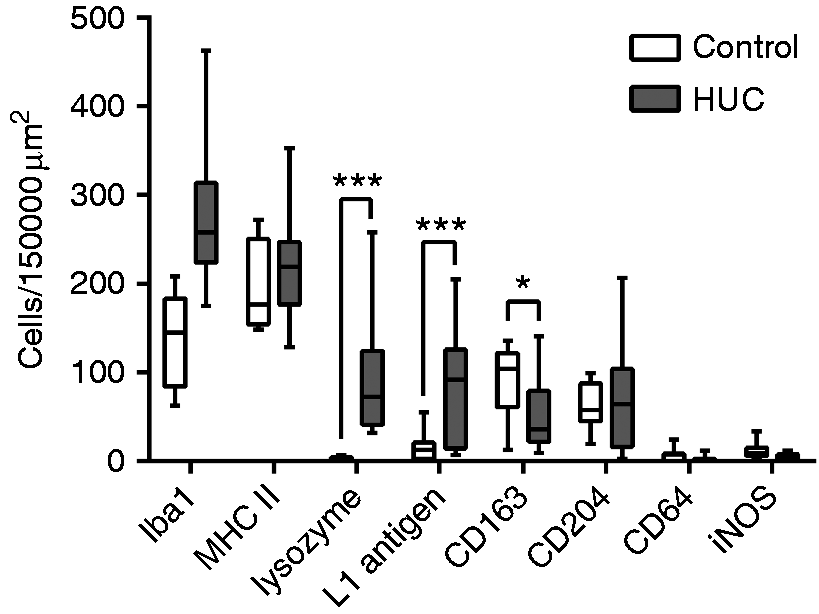
HUC is hallmarked by up-regulation of lysozyme and L1 Ag but decreased amounts of CD163-expressing macrophages
In general, the expression of macrophage markers was strongly associated with the presence of granulomatous foci within the inflamed tissue. However, there was no specific distribution pattern according to anatomical regions (e.g. apical or basal crypt area). Canine HUC was characterised by significantly increased amounts of lysozyme- (P < 0.0002) and L1 Ag-expressing macrophages (P < 0.0075) compared with controls (Figure 1). In contrast, disregarding macrophage subpopulations based on PAS staining, the colonic mucosa exhibited significantly lower numbers of CD163+ macrophages in total in HUC samples compared with the control dogs (P < 0.0465). However, no significant differences were detected in numbers of Iba1-, MHC II-, CD204-, CD64- and iNOS-expressing macrophages in diseased compared with healthy animals.
Phenotypes of macrophage subpopulations in HUC
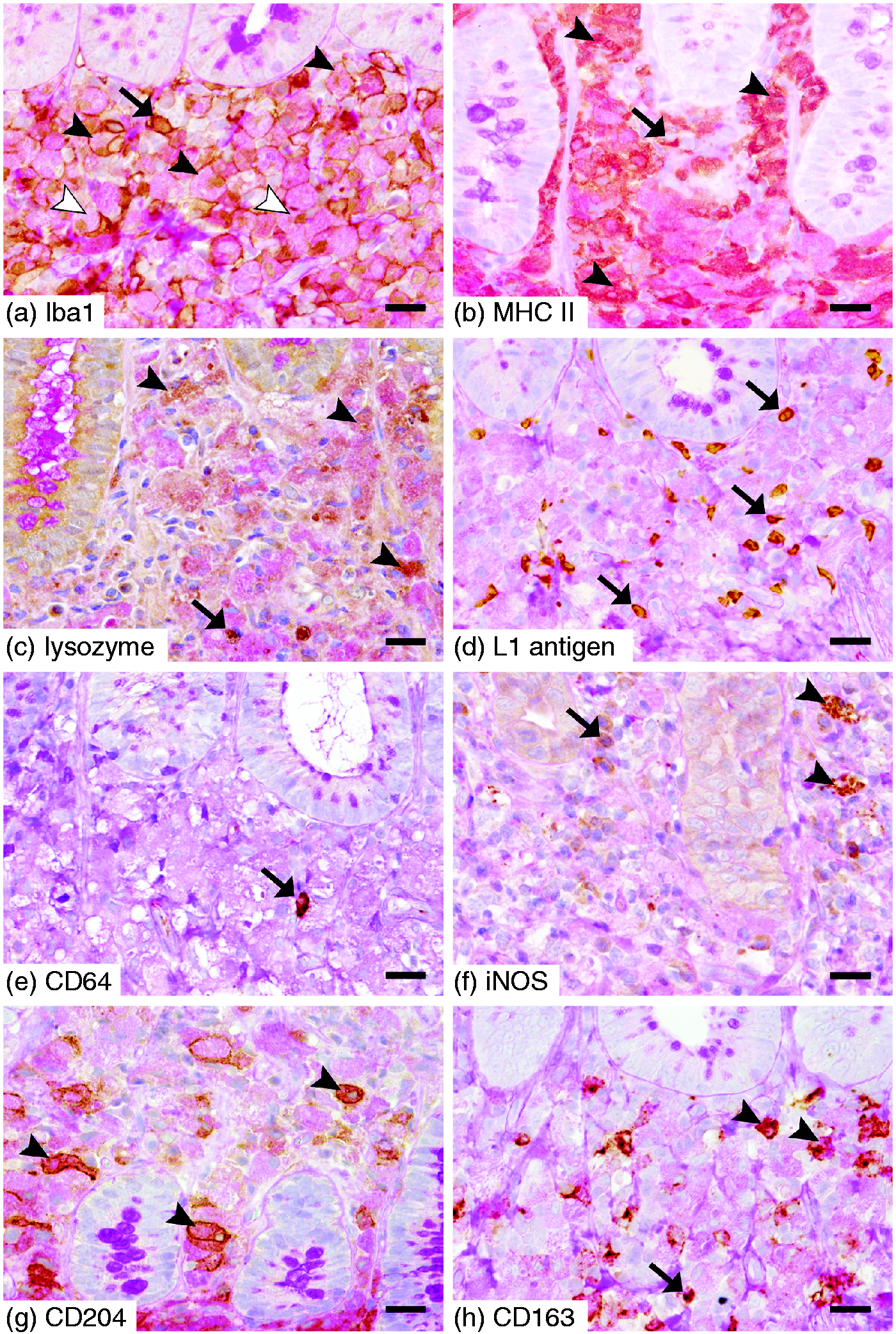
Double-labelling using PAS reaction and IHC for macrophage markers revealed significant differences between PAS+ and PAS– macrophage populations (Figures 2 and 3). The majority of PAS+ and PAS–macrophages were characterised by expression of Iba1 and MHC II. For Iba1, both nuclear and/or cytoplasmic-membranous staining patterns were observed in HUC and the number of Iba1+ cells was significantly increased in PAS+ compared with PAS- macrophages (P < 0.003). In addition, a significant number of PAS+ macrophages showed immunoreactivity for lysozyme, while this was almost absent in PAS– cells (P < 0.002). This was in contrast to L1 Ag (Figure 3d), which predominated in PAS– but was absent in PAS+ macrophages (P < 0.001). Both CD64 and iNOS were expressed by very low numbers of macrophages in canine HUC. However, significant more PAS- macrophages (P < 0.0092) expressed CD64, while the number of iNOS-expressing cells was significantly increased in PAS+ compared with PAS– macrophages (P < 0.018). Significantly elevated numbers of CD204+ cells were detected in the PAS+ subset (P < 0.002). This was in contrast to CD163-expressing cells, which showed no differences between the macrophage subsets.
Expression of macrophage markers in PAS+ and PAS– subsets in canine HUC. (A) Absolute cell numbers per area; (B) percentages. Apart from the difference in numbers of Iba1+ macrophages, most likely reflecting the histopathological pattern of the disease, PAS+ macrophages were characterised by significantly increased expression of lysozyme, iNOS and CD204 but lower expression of L1 Ag and CD64. Note the logarithmic presentation of the y-axis due to pronounced variants in cell numbers between different markers. Wilcoxon signed rank test P-values: *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001. Expression of macrophage markers in the LP of canine HUC. (A) The majority of PAS+ macrophages show expression of Iba1 with a nuclear (white arrowheads) and/or membrane–cytoplasmic staining pattern (black arrowheads). There are few admixed PAS– Iba1+ macrophages (arrow). (B) Note the brown-to-pink cytoplasmic staining indicating expression of MHC II in most of PAS+ macrophages (arrowheads). There are also low numbers of PAS– MHC II+ macrophages present (arrow). (C) Large pink PAS+ macrophages are admixed with PAS+ lysozyme+ cells (arrowheads), as well as a few PAS– lysozyme+ cells (arrow). (D) Note the numerous PAS– L1 Ag+ macrophages (arrows). LP is crowded with PAS+ macrophages that lack expression of L1 Ag. (E) There is only one single PAS– CD64+ cell (arrow) surrounded by large numbers of PAS+ CD64– macrophages. (F) Note the low numbers of PAS+ iNOS+ macrophages (arrowheads) and PAS– iNOS+ cells (arrow). (G) Multiple PAS+ macrophages display a membranous–cytoplasmic expression of CD204 (arrowheads). (H) PAS+ CD163+ macrophages (arrowheads) are admixed with PAS+ CD163– cells, as well as a few PAS– CD163+ macrophages (arrow). ABC immunohistochemistry and PAS reaction, bar = 20 µm.
Correlation analysis revealed strong correlation between Iba1 and MHC II in canine HUC (rho = 0.75, P < 0.001). In addition, there was a moderate association between lysozyme and iNOS (rho = 0.47, P < 0.029), as well as a moderate correlation between L1 Ag and CD64 expression (rho = 0.48, P < 0.023) among HUC macrophage subsets. In contrast, L1 Ag showed a moderate-to-strong negative correlation with lysozyme (rho = –0.84, P < 0.001) and iNOS (rho = –0.52) in canine HUC. CD204 was positively correlated with lysozyme expression (rho = 0.55, P < 0.009) but negatively associated with L1 Ag (rho = –0.47, P < 0.028) among macrophage subsets.
Discussion
Macrophages are heterogeneous members of the mononuclear phagocyte system that are involved in both innate and adaptive immune responses. 24 Despite some morphological and functional similarities between macrophages, monocytes and dendritic cells, these lineages can be distinguished based on their ontogeny. 25 While both monocytes and dendritic cells continually arise from different progenitor cells during adult haematopoiesis, two different origins are currently suggested for macrophages.25,26 First, the majority of classical macrophages present in peripheral tissues arises during the embryonic development and is maintained by self-renewal independent from further haematopoiesis (tissue-resident macrophages).27,28 Second, macrophages can develop from tissue-entering monocytes in adults independent from embryonic precursor cells (monocyte-derived or ‘adult’ macrophages) and this can occur under both steady-state and inflammatory conditions. 17
In the mammalian intestine, macrophages are present in large amounts,29,30 whereby the largest numbers are found in the colon, which is most likely related to its high bacterial load. 31 In contrast to resident counterparts in other tissues, intestinal macrophages do not respond to TLR ligands by producing inflammatory mediators and thereby do not contribute to a pro-inflammatory environment. 17 This selective inertia of intestinal macrophages is needed to maintain epithelial integrity and homeostasis, and disturbances of this condition are supposed to be involved in the pathogenesis of chronic enteric disorders such as Morbus Crohn (MC) and ulcerative colitis (UC) in humans.17,32,33 For several years, it is has been established that classically (M1) and alternatively (M2) activated macrophages are distinguished by associated pro- and anti-inflammatory properties, respectively.34–36 Recently, however, this popular concept of macrophage polarity has been critically discussed, 35 and for intestinal macrophages this bipolar classification also seems to be inappropriate. 37 Therefore, we will continue to use the terms ‘pro-inflammatory’ and ‘anti-inflammatory’ rather than M1 or M2 macrophages.
Canine HUC is characterised by high numbers of PAS+ macrophages crowding the colonic mucosa. Although PAS-stained granules are partly attributed to phagocytosis of bacterial organisms, little is known about the origin, phenotype and function of these cells. Therefore, the aim of this study was to characterise canine colonic macrophages in the inflamed mucosa of HUC patients and healthy controls. This investigation revealed that colonic macrophages highly express Iba1 in both HUC samples and control colons. Iba1 expression is specific for all cells of the monocyte/macrophage lineage except alveolar macrophages in mice and is therefore considered as a ‘pan-macrophage’ marker. 38 In the murine gut, Iba1 expression is high in the LP and most of those cells are double positive for the murine macrophage-specific marker F4/80. 38 Thus, Iba1+ cells are supposed to represent mainly connective tissue macrophages in the murine gastrointestinal tract. In contrast to rodents, Iba1 expression has not been described in the intestine of dogs until now, although it has been proven as a useful macrophage marker in the canine skin. 39 Interestingly, PAS+ macrophages in HUC revealed both nuclear, as well as cytoplasmic-membranous, staining patterns. Iba1 is a key molecule in membrane ruffling and phagocytosis and therefore functions in activated macrophages and microglia. 40 In wild type macrophages Iba1 is localised in the nucleus, while macrophage activation results in Iba1 activity in the cytoplasm, including the interaction with cytoplasmic signal cascades and actin filaments, that contributes to the formation of phagocytic cups. 40 Thus, the strong membranous staining pattern of many PAS+ macrophages in HUC may indicate them to be active in phagocytosis. In addition, the significant difference between Iba1+ PAS+ and Iba1+ PAS– macrophages most likely illustrates the histopathological pattern of HUC, as the colonic mucosa is predominated by huge amounts of PAS+ but only few PAS– macrophages, rather than implicating an up-regulation of Iba-1 in the PAS+ subset.
Colonic macrophages were further featured by high MHC II expression. MHC II is an integral transmembranous molecule, which is required for Ag presentation to CD4+ T cells. 41 High expression of MHC II is also reported in human and murine resident colonic macrophages. 17 A previous study on canine HUC demonstrated an increased MHC II expression in the inflamed mucosa. 15 In contrast, the results of the present study showed no significant differences in numbers of MHC II-expressing macrophages between the inflamed mucosa and control samples. This is an interesting finding as the contribution of bacterial organisms to the development and persistence of HUC as well as the fulminant mucosal inflammation would generally suggest increased local Ag processing and presentation that may be associated with up-regulation of MHC II. 42 The strong correlation between MHC II and the ‘pan-macrophage’ marker Iba1 among macrophage subsets in canine HUC implicates MHC II to label the majority of macrophages rather than discriminating an increase in pro-inflammatory cells by IHC, as this method does not allow differentiation between cytoplasmic and surface MHC II expression. This may elucidate the importance of the use of additional markers other than MHC II when investigating canine macrophages. However, additional studies, including the examination of membranous MHC II expression (e.g. detected by flow cytometry), are needed to further explain the impact of MHC II in the canine colonic mucosa and in canine HUC.
Remarkably, in addition to the presence of Iba1 and MHC II, colonic macrophages expressed scavenger receptors CD163 and CD204 in the steady state. CD163 physiologically mediates the endocytosis of haptoglobulin–haemoglobin complexes by tissue-resident macrophages. 43 This mechanism aims to protect the tissue against haemoglobin-mediated oxidative damage and therefore suggests an anti-inflammatory potential. 44 In contrast, CD204 [also designated as scavenger receptor class A (SR-A)] acts as an innate immune receptor with a broad ligand-binding capacity, including exogenous Ags such as bacterial LPS.45,46 Indeed, there is evidence that CD204 mediates inhibitory cascades limiting the pro-inflammatory immune response stimulated by PAMPs. 47 For example, ligation of SR-A by mAbs induces less IL-6 and IL-12 but increased IL-10 release by LPS-stimulated murine macrophages.48,49 Furthermore, CD204 inhibits the stimulatory capacity of mouse APCs in vitro and in vivo.50,51 Recent observations demonstrate CD163 expression to be a feature of resident colonic macrophages in humans and mice. 17 Although little is known about the role of CD163 and CD204 in the canine gut, the results of the present study implicate canine colonic steady-state macrophages, similar to those in humans and mice, to support an anti-inflammatory environment that is beneficial for intestinal homeostasis and oral tolerance. Interestingly, our results revealed a significant reduction in CD163+ macrophages, and this further implicates canine HUC to be associated with a loss of anti-inflammatory macrophages in the colonic mucosa, which has also been demonstrated in MC. 17 Furthermore, canine HUC showed significantly decreased CD204 expression in PAS– compared with PAS+ macrophages and correlation analysis revealed CD204 expression to be positively correlated with lysozyme but negatively associated with L1 Ag expression. The low CD204 expression in PAS– cells may indicate their pro-inflammatory phenotype. However, the higher presence of CD204 in PAS+ macrophages is an interesting finding that may suggest an increase in pro-inflammatory markers, such as lysozyme and iNOS, rather than a decreased expression of CD204 to be responsible for the phenotypic shift of macrophage in canine HUC. However, this could also implicate an additional role of CD204 under inflammatory conditions (e.g. mediated by its role as an innate immune receptor). Therefore, further studies are needed to elucidate a possible role of CD204 in the canine intestine.
Besides the use of macrophage-specific Abs, we further performed double-labelling with PAS reaction to elucidate the main subsets of canine macrophages in HUC. Thereby, PAS+ macrophages were characterised by marked expression of lysozyme, while it was almost absent in the PAS– subset. Lysozyme is known to enhance the breakdown of bacterial cell walls by catalysing the hydrolysis of specific glycosidic linkages. Furthermore, it has been proved to indicate histiocytic differentiation in human and canine proliferative and granulomatous diseases.52–55 Additionally, lysozyme has been shown to be expressed by canine macrophages but not dendritic cells. 56 The results of the present study suggest that PAS+ rather than PAS– macrophages are mainly involved in bacterial degradation. This is in accordance with the ultrastructural demonstration of bacterial components within the cytoplasm of PAS+ cells in boxer dogs with HUC. 10 Furthermore, lysozyme immunoreactivity is demonstrated in the majority of macrophages in normal canine tissues. 56 In the present study, however, only few lysozyme+ macrophages were present in controls, indicating that significant degradation of microorganisms (such as commensal bacteria) by lysozyme activity is not a feature of the canine colonic mucosa in the steady state.
In contrast to lysozyme, L1 Ag was expressed by many PAS– macrophages, while it was absent from the PAS+ subpopulation. L1 Ag, also designated as calprotectin, is a complex of the calcium-binding proteins S100A8 and S100A9, which mediates antimicrobial and chemotactic properties.57–59 It is present in almost all human circulating monocytes and neutrophilic granulocytes. 60 L1 Ag expression, which can be achieved by IL-1β, TNF-α and LPS, induces the production of pro-inflammatory cytokines, such as IL-1β, IL-6 and IL-8, in human peripheral blood mononuclear cells.61,62 Furthermore, L1 Ag has been demonstrated in human pro-inflammatory macrophages early after their recruitment from the peripheral blood, whereas it is down-regulated later in the macrophage maturation process.63,64 Therefore, PAS− L1 Ag+ macrophages most likely represent immature pro-inflammatory macrophages, illustrating a continuous influx of monocytes to the inflamed mucosa in HUC, whereas PAS+ cells represent mature macrophages that have lost L1 Ag expression. This supports the hypothesis of a previous study where absence of L1 Ag was suggested for PAS+ macrophages, based on serial sections. 15 However, it still needs to be clarified whether PAS+ macrophages are only derived from circulating monocytes or also from tissue-resident cells. In addition, future studies should also focus on changes in circulating monocytes along with other changes in the peripheral blood in HUC.
Investigation of the colonic mucosa in canine HUC demonstrated increased iNOS expression in the PAS+ subpopulation, while significantly smaller amounts of iNOS+ macrophages were PAS–. This was similar to lysozyme, as correlation analysis revealed an association between iNOS and lysozyme. iNOS is a cytoplasmic enzyme catalysing the reaction of amino acid l-arginin to l-citrullin and NO, and plays an important role in the generation of reactive oxygen species (ROS) and thereby in the elimination of pathogens. 65 iNOS is typically expressed by macrophages activated by different pro-inflammatory cytokines, such as TNF-α and IFN-γ, or by microbial products, including LPS.66,67 In contrast, anti-inflammatory macrophages are featured by absence of iNOS but expression of arginase-1, which competes with iNOS for the limited substrate l-arginine. 68 iNOS is expressed by human intestinal epithelial cells and up-regulation can be achieved by infection with enteroinvasive bacteria in vivo.69,70 In UC, high quantities of iNOS have been demonstrated in the inflamed mucosa, while it was absent in inactive disease stages. 71 In addition, iNOS-deficient mice are more susceptible to intestinal damage induced by acetic acid and show a longer course of colitis. 72 Activated canine macrophages have been demonstrated to produce large amounts of NO for the effective destruction of several pathogens, such as canine leishmaniasis73–75 or Trypanosoma cruzi infection. 76 However, studies investigating iNOS in the canine gut and in canine HUC are not available so far. Recently, a genome-wide association scan of granulomatous colitis in boxer dogs and other breeds was performed and revealed single nucleotide polymorphisms associated with granulomatous colitis in the gene encoding neutrophil cytosolic factor (NCF) 2. 3 NCF2 is also known as p67phox and constitutes a cytosolic subunit of the multiprotein complex NADP oxidase, which is also responsible for generation of ROS. 77 This suggests that a defective respiratory burst in canine HUC could be responsible for an impaired interaction of the canine mucosal immune system with intestinal bacteria possibly contributing to the development of clinical signs and pathological lesions. 3 In the present study, iNOS expression was significantly increased in PAS+ compared with PAS– macrophages, and this suggests that PAS+ rather than PAS– cells represent mature pro-inflammatory macrophages, which contribute to the inflammatory process. Interestingly, iNOS was only expressed by a small subset of both PAS+ and PAS– colonic macrophages when considering the total numbers of cells labelled for the ‘pan-macrophage’ marker Iba1. However, it remains unclear whether this result represents a defect in respiratory burst or maybe a pathogen-induced ROS reduction, as described for canine leishmaniasis. 73
CD64 acts as a transmembrane ‘high-affinity’ receptor binding IgG (Fcγ receptor I) and it has been established as a reliable marker to distinguish intestinal macrophages (CD64+) from dendritic cells (CD64–) in mice, 19 and (with some limitations) in humans. 78 CD64+ monocytes have been demonstrated in ischemic canine myocardium, 79 but data on the role of CD64 in the gastrointestinal system of dogs were missing until now. Scant numbers of CD64+ macrophages were present in the colonic mucosa in the steady state. This result is similar to that of the human intestinal macrophage phenotype characterised by a virtual absence of CD64 expression, which discriminates them from blood monocytes. 80 CD64 is suggested to mediate pro-inflammatory functions as its expression can be induced by IFN-γ in human macrophages and neutrophils in vitro, and contributes to Th1 polarisation of T cells during murine colitis.19,81 In addition, significant up-regulation of CD64 by neutrophils has been demonstrated in the inflamed mucosa of patients suffering from MC and UC. 82 In the present study, the colonic mucosa in canine HUC was not characterised by significantly increased amounts of CD64+ macrophages compared with the controls. However, although the numbers of CD64+ cell were very low, its expression predominated in PAS– macrophages but was almost absent in the PAS+ subpopulation. Furthermore, this polarity was correlated with the distribution of L1 Ag among HUC macrophage subsets. This result is in accordance with findings in paediatric MC revealing a strong correlation between expression of CD64 and calprotectin in the ileal mucosa. 83 The authors further demonstrated an association between the CD64 index and both mucosal inflammation and an increased risk to develop an ongoing relapsing clinical course. Therefore, CD64 is discussed as a potential biomarker for intestinal inflammation.82,83 The correlation of CD64 and L1 Ag expression in canine HUC may implicate CD64+ PAS– macrophages to represent pro-inflammatory cells recently recruited from circulating monocytes during colonic inflammation.
Conclusions
In summary, the results of the present study revealed new information concerning phenotypes and possible dynamics of canine colonic macrophages in health and HUC. In control animals, the predominance of Iba1, MHC II, CD163 and CD204 implicated an anti-inflammatory phenotype of canine colonic macrophages in the steady state, possibly contributing to intestinal homeostasis. In addition, the low expression of L1 Ag in control samples suggests that an infiltration of monocytes does not substantially contribute to the pool of canine colonic macrophages under non-inflammatory conditions. Therefore, we suggest that the majority of canine steady-state colonic macrophages most likely represent anti-inflammatory tissue-resident cells. In contrast, decreased expression of CD163 but up-regulation of L1 Ag and lysozyme in canine HUC implicate a switch from an anti-inflammatory to a pro-inflammatory macrophage phenotype contributing to the inflammatory process. Furthermore, main macrophage subsets can be distinguished based on the presence of PAS+ granules in canine HUC. The predominance of L1 and (to a lower extent) CD64 but very low expression of lysozyme and iNOS illustrated PAS– macrophages to represent immature pro-inflammatory macrophages recently recruited from blood monocytes. This implicates that an increased cellular influx significantly contributes to the macrophage populations in canine HUC. In contrast, PAS+ macrophages were characterised by a L1– CD64– lysozyme+ (partly) iNOS+ phenotype, suggesting them to be matured pro-inflammatory cells contributing to phagocytosis and degradation of bacteria. Although we cannot finally rule out partial recruitment and reprogramming of steady-state tissue-resident macrophages under the local influence of pro-inflammatory stimuli such as LPS, data from humans have demonstrated the inability of resident intestinal macrophages to produce pro-inflammatory cytokines in response to several inflammatory factors and thereby they are suggested to mediate ‘inflammatory anergy’. 80 Therefore, PAS+ macrophages in canine HUC may have most likely arisen from infiltrating monocyte-derived PAS– cells, which undergo phenotypic and functional maturation under pro-inflammatory circumstances.
Footnotes
Acknowledgments
The authors wish to thank Jaco van der Lugt, Sophie Maes and Fabio Aloisio for providing case material. Our thanks also go to Franziska Heinrich for testing some of the Abs on canine formalin-fixed tissues, as well as to Bettina Buck, Kerstin Rohn, Caroline Schütz and Petra Grünig for their excellent technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.