
Abstract
Infiltrating activated monocytes are important mediators of damaging inflammation during influenza A virus (IAV) infection. We show that soluble respiratory proteins [collectins, surfactant proteins D (SP-D) and mannose binding lectin (MBL), H-ficolin and LL-37] inhibit replication of seasonal IAV in human monocytes. The collectins and H-ficolin also increased viral uptake by the cells, while LL-37 did not. H-ficolin was able to inhibit replication of the 2009 pandemic H1N1 strain (Cal09) in monocytes, but SP-D and LL-37 had significantly fewer inhibitory effects on this strain than on seasonal IAV. All of these proteins reduced IAV-induced TNF-α production, even in instances when viral replication was not reduced. We used modified recombinant versions of SP-D, MBL and ficolin to elucidate mechanisms through which these proteins alter monocyte interactions with IAV. We demonstrate the importance of the multimeric structure, and of binding properties of the lectin domain, in mediating antiviral and opsonic activity of the proteins. Hence, soluble inhibitors present in airway lining fluid may aid clearance of IAV by promoting monocyte uptake of the virus, while reducing viral replication and virus-induced TNF-α responses in these cells. However, SP-D and LL-37 have reduced ability to inhibit replication of pandemic IAV in monocytes.
Introduction
The innate immune response is critical in containing influenza A virus (IAV) replication during the first few days of infection (i.e. prior to the generation of an adaptive immune response). The innate immune response to IAV is remarkably complex, as recently reviewed. 1 Our laboratory has focused principally on the role of soluble inhibitors in respiratory lining fluids and resident or recruited phagocytes. In the current study, we evaluate effects of the proteins on monocyte responses to IAV. Alveolar macrophages (AMs) play a key role in host defense against IAV.2–4 Mannose binding lectin (MBL) has been shown to reduce uptake of IAV by AMs and also to reduce virus-induced respiratory burst responses and TNF-α generation by these cells. 5 We chose to study human blood monocytes as there is good evidence that these cells are recruited to the lung during IAV infection where they not only play important roles in defense, but also in mediating lung injury.6–8 The role of activated monocytes and pro-inflammatory cytokines may vary depending on the viral strains, with pandemic and avian strains causing more profound and injurious responses than seasonal strains.9–12 TNF-α and IL-6 responses, in particular, have been linked to symptoms and adverse outcomes of IAV infection.13–16 Despite these findings there has been relatively little study of how innate immune proteins modulate human monocytes responses to IAV. In this study we compare how collectins [surfactant protein D (SP-D) and MBL], H-ficolin and LL-37 modulate responses of human monocytes to seasonal and pandemic strains of IAV.
SP-D and MBL play important roles in host response to IAV. 17 SP-D is constitutively present in respiratory lining fluids and its levels rise during IAV infection. MBL is mainly present in blood but does enter lung fluids during IAV infection.18,19 SP-D and MBL contribute to host defense against seasonal IAV strains.19–21 Deletion of the genes for these collectins in mice results not only in increased viral replication, but also increased inflammatory cell infiltration and pro-inflammatory cytokine production.18,22–24 Low levels of SP-D in human lung lavage have been linked to respiratory illnesses, including cystic fibrosis and chronic obstructive pulmonary disease. 25 SP-D and MBL have been shown to modulate interactions of various pathogens, PAMPs like LPS and DAMPs like apoptotic cell components, with monocytes and macrophages. Hence, it seems likely that SP-D and MBL could modulate monocyte responses to IAV.
Ficolins are a related group of proteins that have a similar structure to MBL, including similar N-terminal and collagen domains but with a different carbohydrate recognition domain (CRD). Like MBL, ficolins can fix complement in the absence of Ab (termed the lectin pathway of complement activation). There are three ficolin variants in humans, called H-, M- and L-ficolin. Mice lacking L-ficolin show increased viral replication, as well as increased mass loss and lung inflammation during IAV infection. 26 We have studied human H-ficolin more extensively as this is present at fairly high concentrations in normal human bronchoalveolar lavage fluids (BALF) as is SP-D. 27 Removal of SP-D or H-ficolin from BALF significantly reduces the intrinsic antiviral activity of the fluid.27,28 For collectin and L-ficolin gene-deleted mice the defects in host defense vs. IAV are corrected by treatment with the missing protein.23,24,26 Deficiency of H-ficolin in humans is associated with increased susceptibility to infection. 29 M-ficolin is of interest as it is produced by human monocytes. 30
LL-37 and defensins are antimicrobial peptides that also inhibit IAV.31–34 Treatment of mice with LL-37 not only reduces viral loads, but also reduces inflammatory responses (including pro-inflammatory cytokine production) during IAV infection. 32 Deficiency of LL-37 in humans is also linked to recurrent infections. 35
We here present comparative effects of SP-D, MBL, H-ficolin, M-ficolin and LL-37 on human monocyte responses to IAV and also evaluate mechanisms of action through use of various modified versions of the proteins.
Materials and methods
Innate immune protein preparations
Innate immune proteins used in this study.

HNP1 was purchased from Bachem (Torrance, CA, USA). H-ficolin and M-ficolin were prepared by Steffen Thiel, as described previously. 39 MBL and chimeric proteins containing N-terminal and collagen domain elements from L-ficolin and carbohydrate recognition domain of MBL were developed as described by Kazue Takahashi and Ian Michelow. 40
Virus preparations
Viral strains used in the paper are outlined in Table 2. Philippines 82/H3N2 (Phil82) strain was kindly provided by E Margot Anders (University of Melbourne, Melbourne, Australia) and the PR-8 strain was kindly provided by Jon Abramson (Wake Forest University, NC, USA). These strains were grown in the chorioallantoic fluid of 10-d-old chicken eggs and purified on a discontinuous sucrose gradient as previously described. 41 The virus was dialyzed against PBS to remove sucrose, aliquoted and stored at −80℃ until needed. Post-thawing, the viral stocks contained ∼5 × 108 infectious focus forming units/ml. The California 2009 H1N1 pandemic strain (Cal09) and the New York 2001 H1N1 (NY01) seasonal strain were prepared by reverse genetics as described. 42
Neutrophil and monocyte preparation
Neutrophils from healthy volunteers were isolated to >95% purity by using dextran precipitation, followed by Ficoll–Paque gradient separation for the separation of mononuclear cells (layering above the Ficoll–Paque) and neutrophils (below the Ficoll–Paque). The neutrophils were purified further by hypotonic lysis to eliminate any contaminating erythrocytes, as previously described. 41 Cell viability was determined to be >98% by trypan blue staining. The isolated neutrophils were re-suspended at the appropriate concentrations in PBS and used within 2 h. Neutrophil collection was done with informed consent, as approved by the Institutional Review Board of Boston University School of Medicine. PBMCs were taken from the layer above Ficoll–Paque and washed several times in PBS. Monocytes were isolated from the PBMC preparations by negative selection using magnetic beads and a Miltenyi monocyte isolation kit (catalogue number 130-091-153; Miltenyi Biotech, Bergisch Gladbach, Germany). RAW cells were purchased from ATCC.
Fluorescent focus assay of IAV replication in monocytes
Monocytes were allowed to adhere to 96-well plates and then infected with diluted IAV preparations for 45 min at 37℃ in PBS. The MOI for these assays was ∼0.1. The virus had been pre-incubated for 30 min at 37℃ with various concentrations of the innate immune proteins or control buffer, followed by addition of these viral samples to the monocytes. The cells were then washed and re-suspended in RPMI with 10% heat-inactivated, autologous serum. No serum was present during incubation of the virus with the monocytes. The monocytes were tested for presence of IAV-infected cells after 20 h of virus infection using a monoclonal Ab directed against the influenza A viral nucleoprotein (provided by Nancy Cox, CDC, Atlanta, GA, USA), as previously described. In prior studies we have shown that results of this assay correlate with results of plaque assays or release of viral RNA into culture supernatants. 31
Measurement of IAV uptake by neutrophils and monocytes
FITC-labeled IAV (Phil82 strain) was prepared, and uptake of virus by neutrophils, monocytes or RAW cells was measured by flow cytometry as described. 43 In brief, IAV was treated with various doses of LL-37 for 30 min at 37℃. Then it was incubated with cells for 45 min at 37℃ in presence of control buffer. Trypan blue (0.2 mg/ml) was added to these samples to quench extracellular fluorescence. Following washing, the cells were fixed with 1% paraformaldehyde and neutrophil associated fluorescence was measured using flow cytometry. The mean cell fluorescence (>1000 cells counted per sample) was measured.
Assay of TNF-α and IL-6 production by human monocytes
Human peripheral blood monocytes were isolated by magnetic bead separation as described above and infected with an MOI of ∼50 of Phil82 IAV for 45 min at 37℃. The virus was either used alone or after 30 min incubation with various concentrations of LL-37. After this the cells were pelleted and then cultured for 24 h at 37℃ in RPMI with 10% heat-inactivated autologous serum in a CO2 incubator. After 24 h, supernatant was collected and assayed for TNF-α using a sandwich ELISA method (catalogue number MTNF-α AI; Endogen, Woburn, MA, USA) or IL-6 using an ELISA kit (Abcam, Cambridge, MA, USA). Results were expressed as percent of control TNF-α or IL-6 production. The mean level of TNF-α production by monocytes treated with IAV alone was 220 ± 10 ng/ml. Levels of TNF-α produced by cells treated with PBS alone (control cells) were between 0 and 20 ng/ml in different sets of experiments. IL-6 levels in control or IAV treated cells were 15.6 ± 3 and 35.5 ± 6 ng/ml.
Statistics
Statistical comparisons were made using Student’s paired, two-tailed t-test or ANOVA with post hoc test (Tukey’s). ANOVA was used for multiple comparisons to a single control.
Results
Effect of SP-D, H-ficolin, M-ficolin or LL-37 on replication of seasonal or pandemic IAV in human monocytes
As shown in Figure 1(a), human SP-D inhibited the replication of the Phil82 H3N2 seasonal IAV strain in human monocytes. SP-D did not inhibit replication of the 2009 pandemic H1N1 strain (Cal09). In contrast, H-ficolin was able to inhibit replication of both Phil82 and Cal09 H1N1 pandemic strain in these cells (Figure 1b). H-ficolin has a different mechanism of binding to, and inhibition of, IAV compared with the collectins SP-D and MBL. Whereas SP-D and MBL bind to carbohydrate epitopes on the viral hemagglutinin, H-ficolin contains a sialylated glycan to which IAV binds. Cal09 has minimal glycan attachments on its hemagglutinin (HA) and this likely accounts for the failure of SP-D to inhibit this viral strain. The distinct mechanism of activity of H-ficolin allows it to inhibit viruses lacking glycosylation on their HA. As a further example of this, H-ficolin inhibited replication of the PR-8 strain. This strain completely lacks glycan on the head region of its HA and is fully resistant to SP-D or MBL. LL-37 caused a lower degree of inhibition of Cal09 than of Phil82 or NY01 (a seasonal H1N1 strain). M-ficolin did not inhibit infection of monocytes with either Phil82 or PR-8 virus in five experiments (data not shown).
Comparison of the ability of SP-D, H-ficolin and LL-37 to inhibit replication of the Phil82 H3N2, Cal09 H1N1, PR-8 H1N1 or NY01 H1N1 strains of IAV in human monocytes. (a) SP-D strongly inhibited replication of the Phil 82 strain in human monocytes but was unable to inhibit that of Cal09. (b) H-ficolin inhibited replication of Phil82, Cal09, and PR-8 strains to similar extents. (c) LL-37 inhibited the Phil82. (d) LL-37 also inhibited NY01 strains to a significantly greater extent than Cal09. *P < 0.05 vs. control. Results are mean ± SEM for four or more experiments.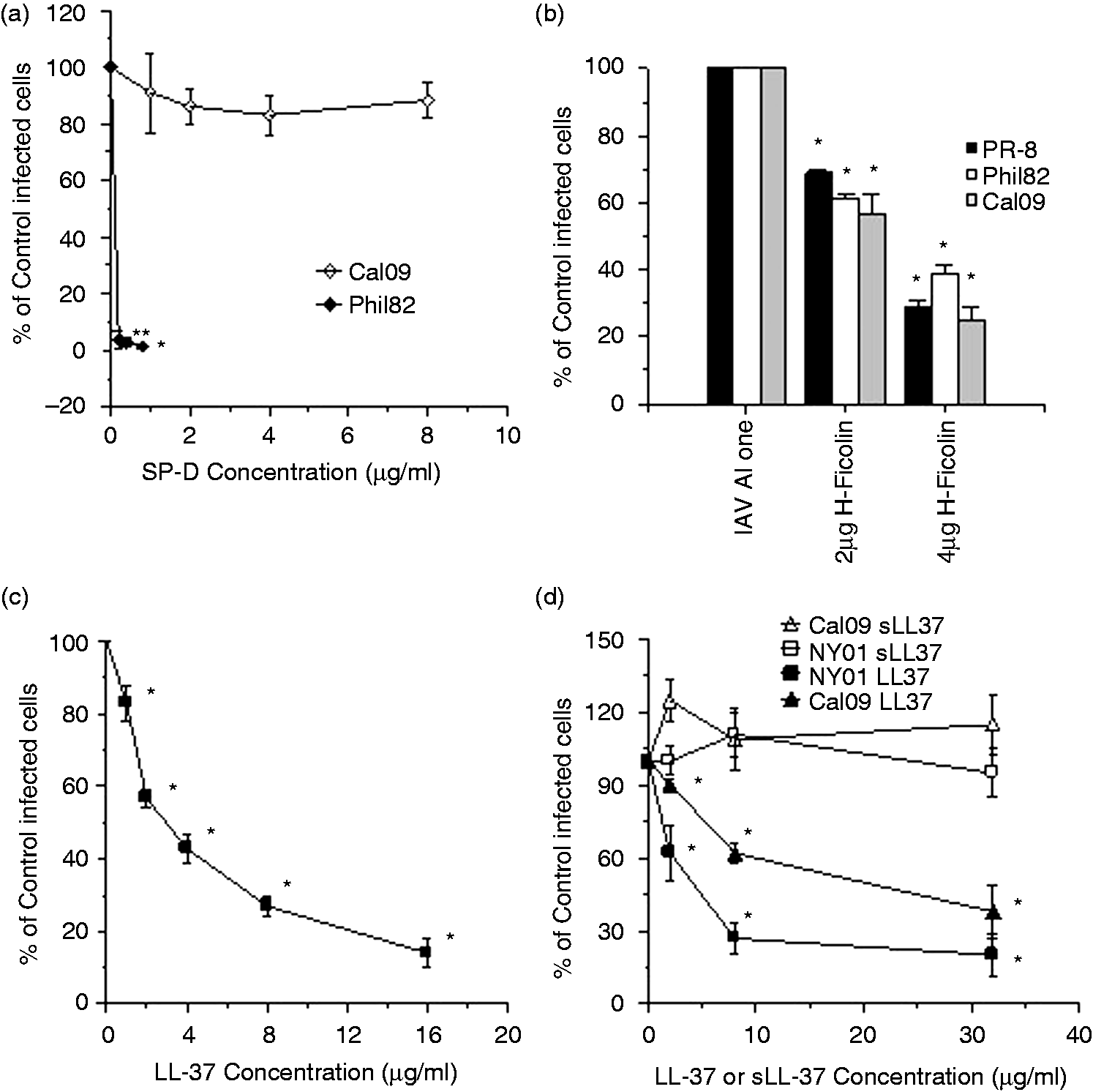
Effects of SP-D, MBL, H-ficolin, M-ficolin and LL-37 on monocyte uptake of IAV
As shown in Figure 2(a), SP-D and MBL increased uptake of Phil82 seasonal IAV by human monocytes. In contrast, H-ficolin and LL-37 did not significantly alter uptake. We included HNP-1 as a control antimicrobial peptide and this was able to increase monocyte uptake of this strain of IAV. H-ficolin and M-ficolin were able to increase monocyte and neutrophil uptake of the PR-8 strain while MBL was not, in keeping with the distinct mechanisms of action of these proteins (Figure 2b, c).
Comparison of the ability of SP-D, MBL, H-ficolin, M-ficolin, LL-37 or HNP-1 to increase uptake of IAV by human monocytes or neutrophils. (a) SP-D caused a marked increase in human monocyte uptake of Phil82 IAV. MBL also significantly increased uptake of this strain but H-ficolin did not. (b) H-ficolin and M-ficolin increased uptake of the PR-8 strain of IAV, while MBL did not. (c) H-ficolin and M-ficolin caused increased uptake of PR-8 by neutrophils, while MBL did not. MBL increased uptake of Phil82 by these cells (data not shown). (d) LL-37 did not increase Phil82 uptake by monocytes, while HNP-1 did increase uptake. *P < 0.05 vs. control. Results are mean ± SEM for four or more experiments.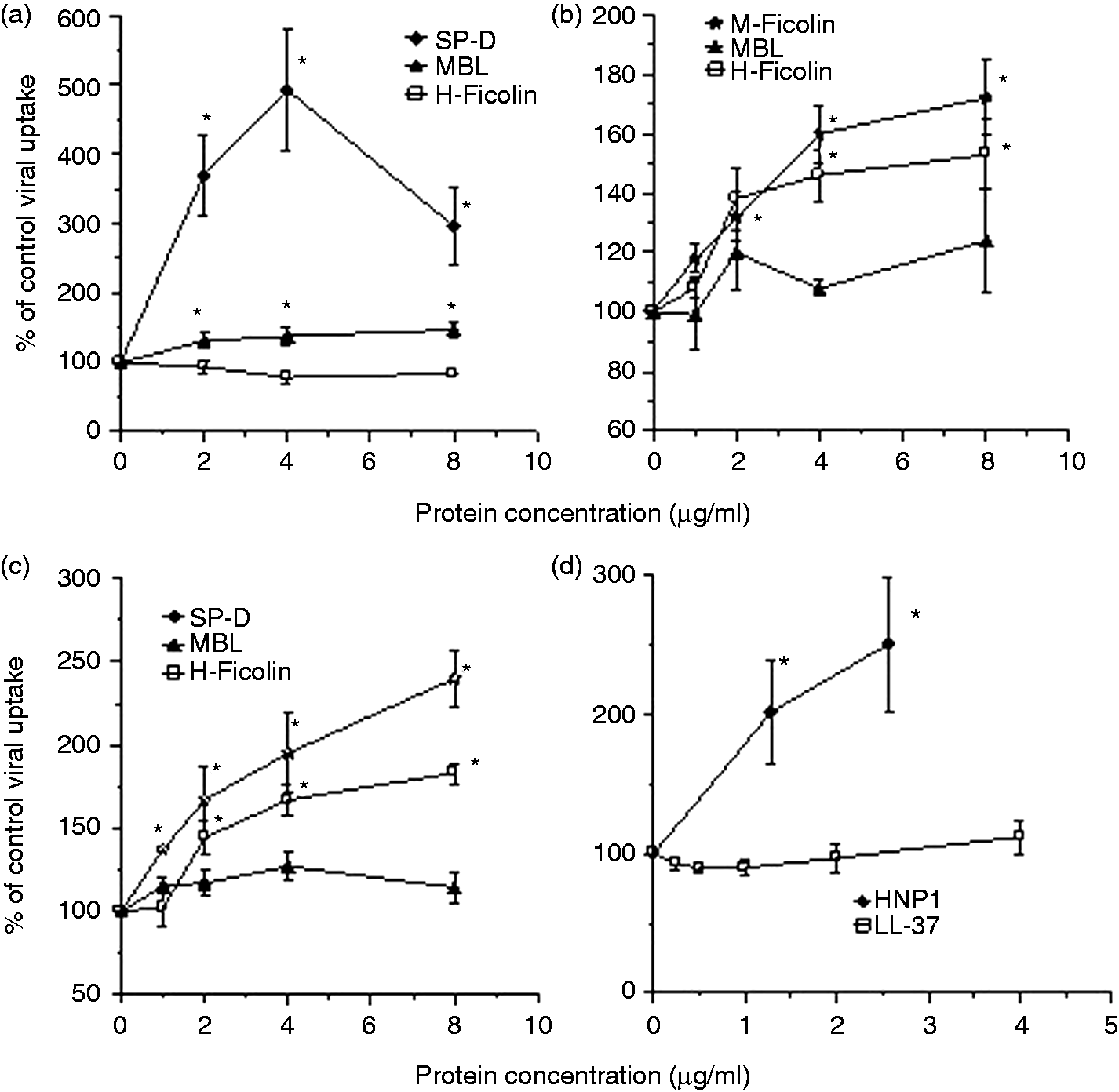
Effects of SP-D, H-ficolin, M-ficolin and LL-37 on monocyte cytokine responses to IAV
As shown in Figure 3(a), SP-D, MBL and H-ficolin inhibited the robust TNF-α production induced by Phil82 IAV in human monocytes. M-Ficolin did not inhibit TNF-α production induced by Phil82 (n = 3; data not shown). SP-D and H-ficolin also inhibited TNF-α production in response to Cal09 (Figure 3b). MBL showed a trend to decrease TNF-α production in response to Cal09, but this was not significant. LL-37 inhibited both TNF-α and IL-6 responses to Phil82 IAV in a dose-related manner. For all of the proteins tested inhibition was seen when the virus was pre-incubated with the protein and not when the protein was added to the cells after viral infection.
SP-D, MBL, H-ficolin and LL-37 inhibit IAV-induced cytokine responses of human monocytes. (a) SP-D, MBL and H-ficolin reduced monocyte TNF-α response to the Phil82 strain of IAV. (b) SP-D, MBL and H-ficolin inhibit the virus-induced TNF-α response to Cal09 as well. (c) LL-37 inhibited both TNF-α and IL-6 responses to Phil82 IAV, whereas the scrambled peptide (sLL-37) did not. LL-37 alone in the absence of IAV did not stimulate any increased TNF-α response compared with control buffer (PBS). *P < 0.05 vs. control. Results are mean ± SEM for four or more experiments.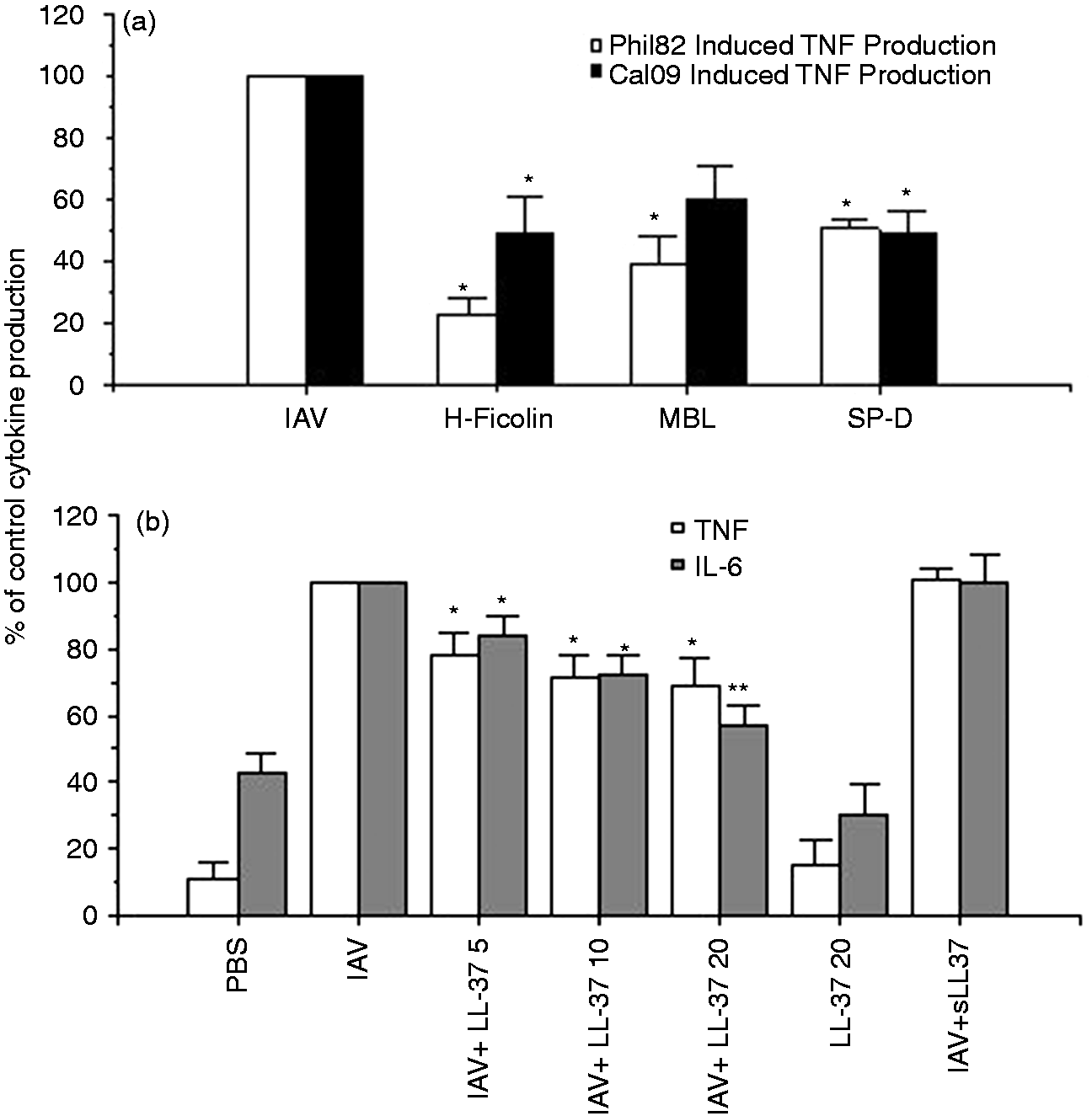
Mechanism through which SP-D modulates interaction of IAV with monocytes
Role of the collagen domain and multimerization
Wild type rat SP-D reduced viral replication of Phil82 virus in murine monocytic RAW cells as assessed by production of viral nucleoprotein at 24 h after infection (Figure 4a). This occurred despite the fact that SP-D increased RAW uptake of the virus (Figure 4b). We used additional recombinant versions of rat SP-D in which the collagen domain was either shortened or deleted, or in which both the collagen domain and N-terminus were deleted, to determine the role of the N-terminus and collagen domain in these responses (Table 2).
Effect of various recombinant SP-D preparations on replication in or uptake of Phil82 IAV by monocytic RAW cells. (a) Percent of control viral nucleoprotein expressing RAW cells 24 h after virus infection. Each of the recombinant rat SP-D preparations caused significant reduction in viral protein expression at the concentrations of 300 ng/ml or higher. The effect of SP-Dmini was significantly greater than that of SP-Dcdm or wild type recombinant rat SP-D (RrSP-D). (b) SP-Dmini, SP-Dcdm and RrSP-D all caused significant increase in fluorescent viral uptake at concentrations of 3 µg/ml or higher. rNCRD did not increase uptake. (c) Recombinant human SP-D (RhSP-D) caused increased viral uptake at all concentrations tested, whereas huNCRD did not. When huNCRD was cross-linked with MAb 240-08 it did increase viral uptake at all concentrations tested. Results are mean ± SEM for four or more experiments and differences were considered significant at P < 0.05 using t-test. Influenza viral strains used in this study.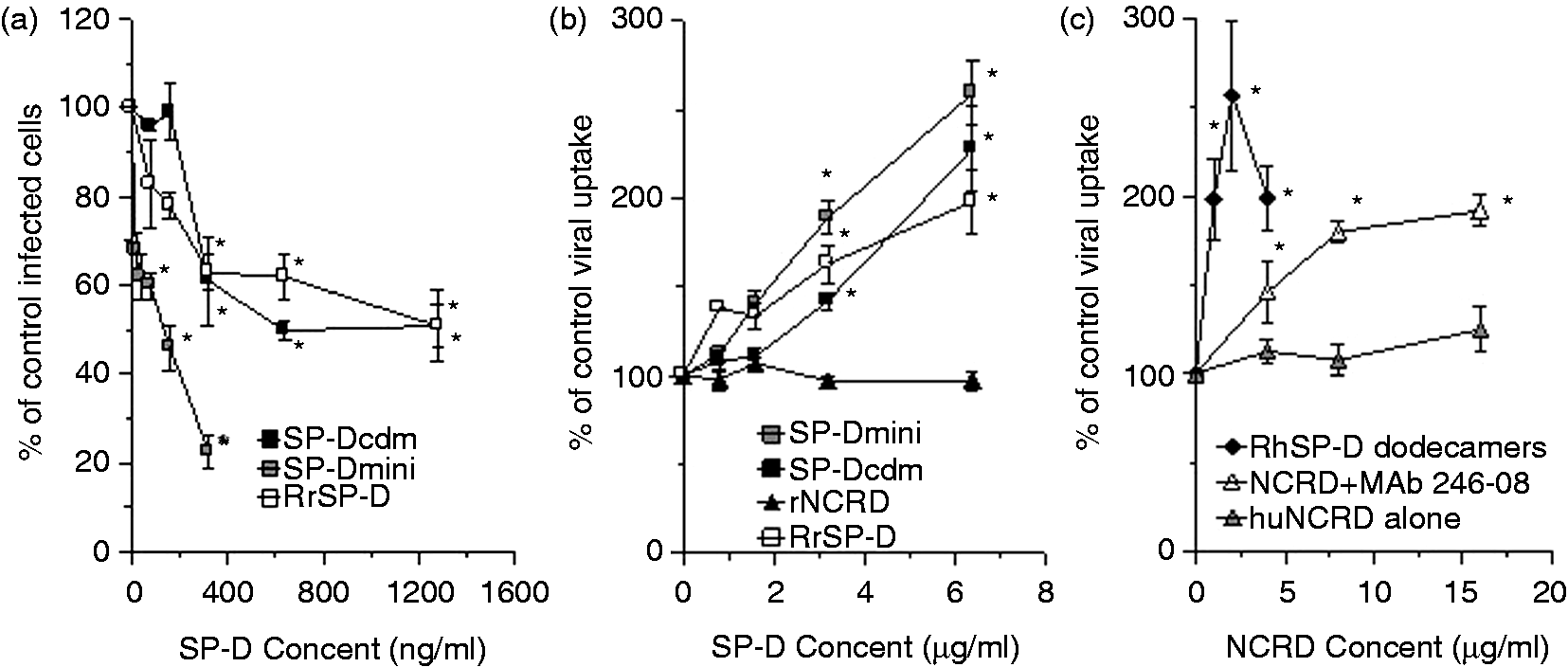

The preparation with the shortened collagen domain (called SP-Dmini) actually had greater ability than the wild type SP-D to suppress viral replication or increase viral uptake by RAW cells (Figure 4a, b). SP-Dcdm completely lacks the collagen domain but retains the N-terminus, allowing it to form multimers with generally four NCRD trimers per molecule.37,38 SP-Dcdm has been shown to restore the ability of SP-D–/– mice to control IAV replication and has significant viral neutralizing activity in vitro. 38 SP-Dcdm reduced viral replication in, and increased viral uptake by, RAW cells. The activity of SP-Dcdm in these assays was similar to that of wild type rat SP-D. These results indicate that the collagen domain per se is not required for these activities of SP-D. The results also indicate that the ability to form multimers and cross-link IAV particles is critical (i.e. as the rNCRD alone did not alter viral interactions with monocytes).
Another way we have addressed the role of cross-linking in activity of SP-D is to use mAbs which are able to cross-link trimeric NCRDs without blocking binding activity. As shown in Figure 4(c), wild type human SP-D NCRD trimers (huNCRD) did not increase Phil82 uptake by RAW cells. However, when the trimers were cross-linked with mAb viral uptake was increased.
Role of the CRD
We have prepared modified versions of the trimeric NCRD domain of SP-D that have increased neutralizing activity for seasonal IAV compared with wild type huNCRD.44–46 The single substitution of arginine 343 with valine (R343V) strongly increased antiviral and aggregating activity. Adding an alanine or serine substitution for asparatic acid 325 caused a small increase in activity, but the combined D325A (orS) and R343V gave the greatest increase in neutralizing and viral aggregating activity. As shown in Figure 5(a), D325A + R343V or R343V significantly reduced replication of human monocyte replication of Phil82 IAV, while huNCRD had minimal effect. D325A + R343V and R343V also inhibited replication of Cal09 in human monocytes while huNCRD did not (Figure 5b). D325A + R343V also strongly increased human monocyte and RAW cell uptake of IAV while huNCRD did not (Figure 5c, d).
Effects of (a, b) wild type huNCRD or mutant versions of huNCRD viral replication in human monocytes, (c) human monocyte uptake of IAV or (d) RAW cell uptake of IAV. (a) Ability of huNCRD to inhibit replication of Phil82 in human monocytes compared with that of the mutant NCRDs, D325A + R343V and R343V. The mutant proteins caused significantly greater inhibition of Phil82 IAV than huNCRD. (b) Similar experiments but using the Cal09 IAV strain. Here, again, the mutant peptides caused significantly greater inhibition of replication than huNCRD (which did not significantly inhibit this strain compared to control). (c) D325A + R343V caused significant increases in uptake of Phil82 IAV by human monocytes, while huNCRD did not. (d) D325A + R343V caused significantly increased viral uptake by RAW cells at all concentrations tested. R343V did so at the highest concentration tested. huNCRD again did not increase uptake. Results are mean ± SEM for four or more experiments. *Significant change compared with control (P < 0.05); #D325A + R343V had a significantly greater effect than the other collectins tested in (b) (ANOVA).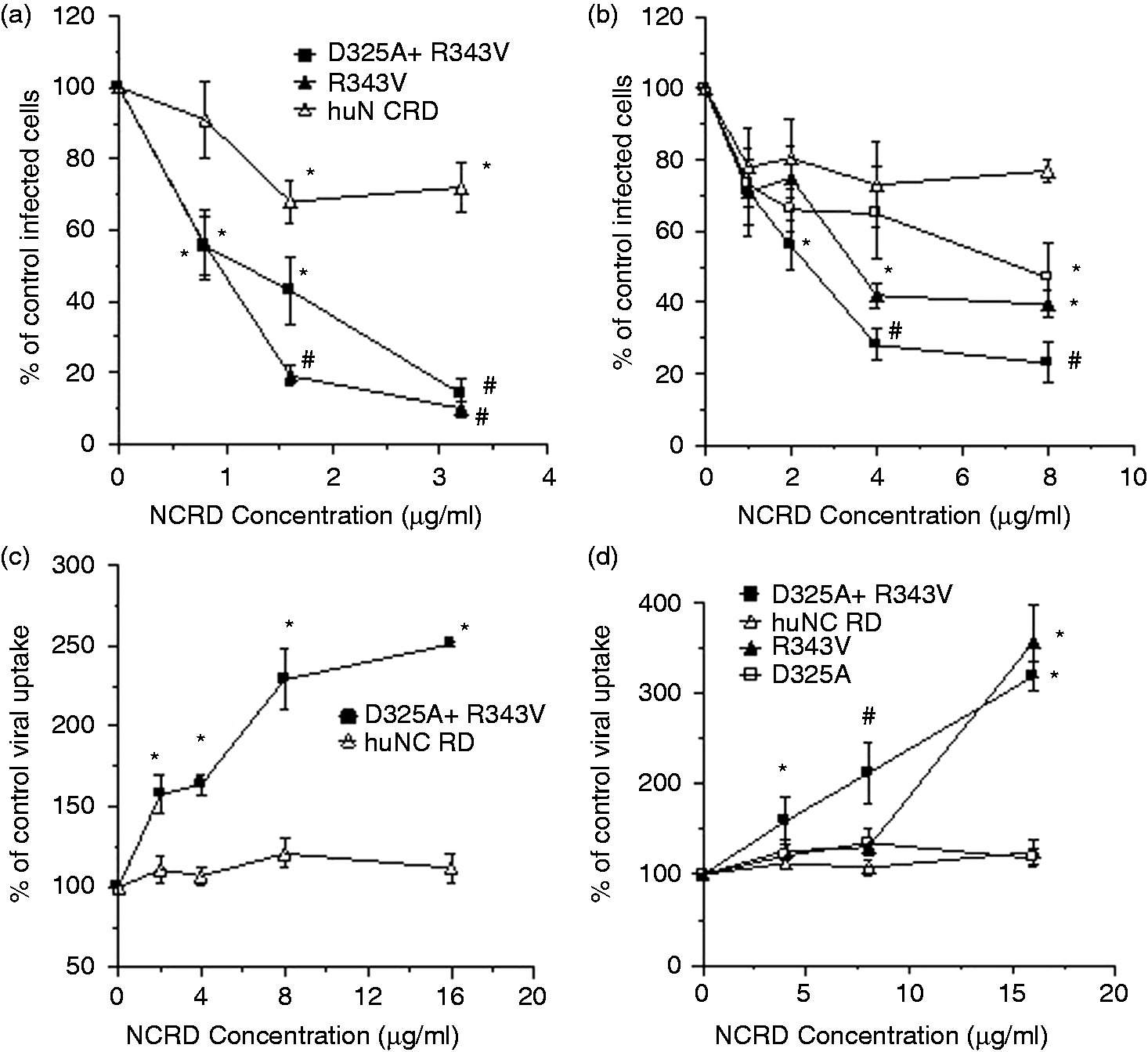
Unexpectedly, huNCRD inhibited human monocyte TNF-α production triggered by both Phil82 and Cal09 viruses, as did D325A + R343V (Figure 6). The ability of SP-D, D325A + R343V or huNCRD to inhibit TNF-α production required direct incubation of the virus with the proteins, as addition of the proteins to the cells prior to infection did not reduce TNF-α production (data not shown). For the Cal09 virus, D325A + R343V caused significantly greater inhibition of TNF-α than either SP-D or huNCRD.
Effects of huNCRD or D325A + R343V on monocyte TNF-α production stimulated by Phil82 or Cal09 IAV strains. huNCRD and D325A + R343V inhibited TNF-α production significantly. Both proteins inhibited TNF-α response to Cal09, but the effects of D325A + R343V were significantly greater than those of huNCRD. *P < 0.05 vs. control. Results are mean ± SEM for four or more experiments.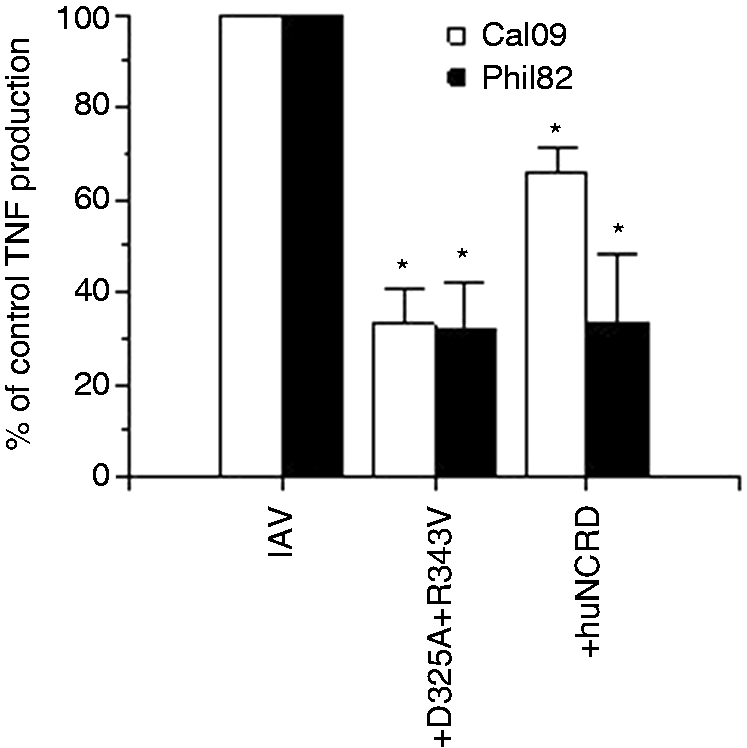
Altering the pattern of viral inhibition by ficolins by replacing their CRD with that of MBL
We have previously reported that chimeric molecules containing various lengths of the N-terminal domains of human L-ficolin and carbohydrate recognition domain of human MBL have increased host protective effects for IAV in vitro and in vivo compared with MBL.47,48 One of these chimeric proteins, called RCL2, had the strongest viral aggregating and HA inhibiting activity with the Phil82 strain. As shown in Figure 6a, RCL2 also caused significantly increased human neutrophil and monocyte uptake of Phil82 IAV. At lower concentrations, the effects of RCL2 were greater than MBL or another chimeric protein RCL3. Despite having reduced antiviral activity compared to RCL2 in vitro and in vivo, RCL3 was protective in a mouse model of IAV infection as evidence by reduced pro-inflammatory cytokine production. 48 As shown in Figure 6(c) both RCL2 and RCL3 strongly reduced human monocyte TNF-α responses to IAV.
Discussion
Recruited monocytes play an important role in host defense and inflammatory responses to IAV. We show that the innate immune proteins SP-D, MBL, H-ficolin and LL-37 fluid can reduce replication of seasonal IAV in monocytes. SP-D and H-ficolin are constitutively present in BALF.27,28 In murine models, the levels of SP-D increase during IAV infection. MBL and LL-37 are mainly detectable in BALF during inflammation. For example, we were not able to show any measurable amount of LL-37 in healthy donor BAL fluid (data not shown). SP-D was not able to inhibit replication of the Cal09 pandemic strain in monocytes, and LL-37 had greatly diminished activity compared with its effect on seasonal H3N2 or H1N1 strains. However, H-ficolin did inhibit replication of Cal09 in monocytes. These results parallel effects of the proteins on viral replication in epithelial cells, with one exception: LL-37 did not inhibit Cal09 in MDCK cells or respiratory epithelial cells but did inhibit it, to some extent, in monocytes. The ability of H-ficolin to inhibit Cal09 presumably relates to its distinctive mechanism of attachment to IAV compared with SP-D or MBL.
SP-D and MBL were able to increase monocyte uptake of Phil82 IAV. H-ficolin and M-ficolin increased uptake of the PR-8 strain. It seems likely that the ability of these proteins to increase viral uptake relates to their ability to cause viral aggregation. LL-37 does not cause viral aggregation, 31 nor does it increase uptake of virus by neutrophils or monocytes (Figure 2). 49 In the case of SP-D multimers with many NCRD heads or cross-linking of isolated NCRDs was needed to increase uptake. The collagen domain of SP-D was not necessary for viral inhibition or increasing viral uptake by monocytes, as evidenced by the fact that SP-Dcdm (which lacks the collagen domain) retains these activities. Of interest, shortening the collagen domain of SP-D (as in SP-Dmini) led to greater activity that the wild type protein. We also found this protein to have increased viral aggregating activity compared with wild type SP-D. 37 Wild type human or rat NCRD trimers did not neutralize IAV or increase uptake by monocytes. However, isolated mutant NCRDs (D325A + R343V or R343V) were able to increase viral uptake. We have previously shown that these preparations can cause viral aggregation, while wild type NCRDs (rat or human) cannot. Overall it seems likely that viral aggregating ability relates to ability to increase uptake by monocytes. The mutant NCRDs, D325A(or S) + R343V or R343V, have higher binding affinity for mannosylated glycans on the Phil82 HA and also bind HA in a different orientation as compared with wild type NCRD. 50 These differences increase the ability of the mutants to block the sialic acid binding site of the HA and to cross-link HA molecules which is likely how they cause viral aggregation.
We have found that D325A(orS) + R343V and R343V can inhibit replication of Cal09 in respiratory epithelial cells, presumably reflecting their greater ability to bind to the single oligosaccharide present on the head region of Cal09. 44 In keeping with this the mutant NCRDs were able to inhibit replication of Cal09 in monocytes, while full-length SP-D could not. 44
H-ficolin was able to increase uptake of PR-8 but not the Phil82 strain of IAV by neutrophils and monocytes. This is in keeping with its different mechanism of viral attachment to IAV. We speculate that lack of glycans on the HA head of PR-8 results in less steric hindrance on binding to sialylated ligands (such as that expressed on the NCRD of H-ficolin).
27
H-ficolin was also able to aggregate particles of the PR-8 strain but not of Phil82 (data not shown). This is again consistent with the hypothesis that viral aggregation by the innate proteins is necessary for increasing monocyte or neutrophil uptake of viral. Replacing the NCRD of ficolin with that of MBL alters the binding properties of the protein and allowed it to cause aggregation of Phil82,
47
and increase its uptake by neutrophils and monocytes (Figure 7). The chimeric protein RCL2 has increased anti-Ebola and anti-IAV compared with wild type MBL.40,47 This appears to result from distinctive properties of the L-ficolin collagen domain and resulting quaternary structure of the protein.
40
RCL2 was also had greater ability to increase viral uptake by neutrophils and monocytes in the current study (Figure 7).
Effects of MBL or recombinant L-ficolin/MBL chimeras on (a, b) neutrophil or monocyte uptake of Phil82 IAV, or (c) IAV-induced TNF-α production. MBL and the chimeras (RCL proteins) caused significant increases in neutrophil and monocyte uptake of Phil82 IAV. (b) MBL, RCL2 and RCL3 increased uptake of Phil82 by monocytes. (c) RCL2 and RCL3 strongly inhibited TNF-α responses of human monocytes to Phil82 IAV. *P < 0.05 vs. control. Results are mean ± SEM for four or more experiments. **Effect of RCL2 was significantly stronger than that of MBL or RCL3.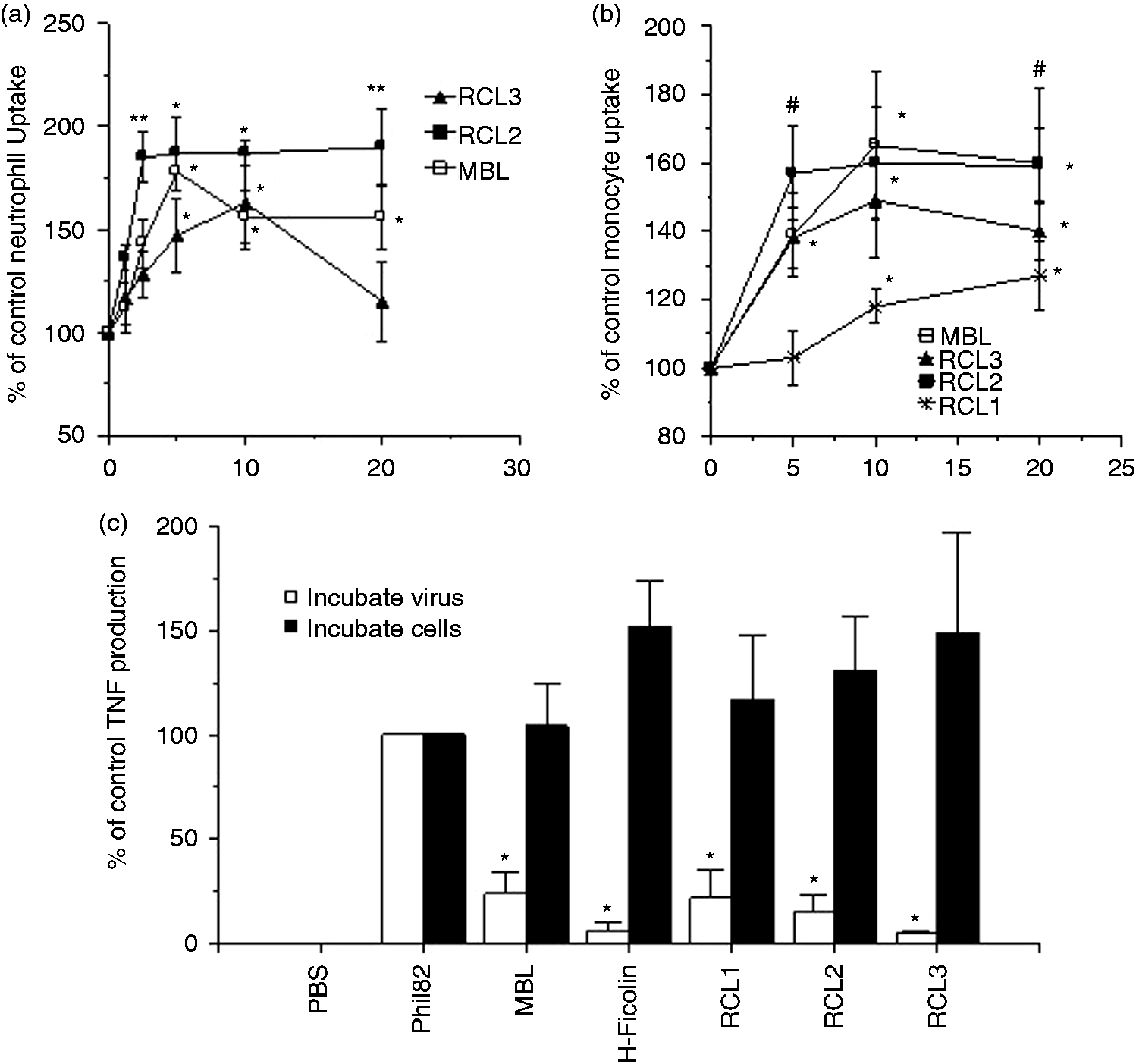
Increased production of TNF-α and or IL-6 during IAV infection is associated with disease severity. 51 SP-D and MBL knockout mice have increased pro-inflammatory cytokine production in response to IAV infection suggesting that collectins down-regulate inflammatory responses in this context. 17 Barlow et al. have shown that LL-37 treatment of IAV infected mice reduces viral replication and pro-inflammatory cytokine production. 32 LL-37 has also been shown to reduce cytokine responses to LPS. 52 We found that SP-D, MBL, H-ficolin and LL-37 all reduced cytokine responses to Phil82. This effect required pre-incubation of the virus with the proteins. The ability to reduce cytokine responses did not always correlate with inhibition of replication as evidenced by the ability of SP-D to inhibit TNF-α response to Cal09, and huNCRD to inhibit this response to Phil82 or Cal09. D325A + R343V caused greater inhibition of TNF-α response to Cal09 than SP-D or huNCRD, but all of these peptides inhibited the response. Of special interest was the finding that huNCRD was able to suppress TNF-α induced by Phil82 or Cal09 viral strains, despite having minimal or no neutralizing activity. With regard to Cal09, D325A + R343V still had greater TNF-α suppressing activity than huNCRD.
Conclusions
Our findings are consistent with a role for SP-D, MBL, H-ficolin, M-ficolin and LL-37 in facilitating clearance of seasonal IAV by monocytes through inhibiting viral replication, and – in the cases of collectins and the ficolins – increasing viral uptake by these cells. All of these proteins, with the exception of M-ficolin also limited pro-inflammatory cytokine production stimulated by IAV in monocytes. In contrast, SP-D had no effect on replication of pandemic H1N1 in monocytes, and the effect of LL-37 was significantly reduced compared with its effects on seasonal IAV strains. These results may have relevance to the increased pathogenicity of pandemic IAV, as it can bypass restriction by some key innate inhibitors in respiratory lining fluid.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIH RO1 HL069031.