
Abstract
IL-17A-dependent immunity is of importance in the protection against extracellular bacterial pathogens. However, IL-17A is also suggested to mediate the pathogenesis of lung diseases, such as acute respiratory distress syndrome. Here, we studied the role of IL-17A in a mouse model of acute pneumonia. IL-17A mediated the expression of keratinocyte-derived chemokine (KC) and the recruitment of inflammatory cells in mice infected with a sub-lethal dose of Pseudomonas aeruginosa. IL-17A deficiency protected mice from lethal P. aeruginosa lung infection. A sub-lethal infection with Streptococcus pneumoniae resulted in increased bacterial burden associated with increased pulmonary inflammation. Thus, the type of infectious bacteria seemed to influence the way in which IL-17A functions during pulmonary infection. Reducing pulmonary inflammation by targeting IL-17A may be a therapeutic option in acute P. aeruginosa pneumonia.
Introduction
Innate and adaptive immunity are necessary to eliminate inhaled pathogens in the lung. 1 In the case of acute pulmonary infections, the inflammatory response of the host needs to be closely regulated to ensure efficient elimination of pathogens and the prevention of immunopathology. However, severe lung infections are also connected with deregulated inflammation leading to lung damage, edema, and loss of pulmonary function.2–5
IL-17A (often named IL-17) is the best-studied member of the IL-17 cytokine family that comprises at least six IL-17 subtypes (IL-17 A to F). 6 IL-17A contributes to the clearance of microbial infections via the induction of the expression of cytokines, chemokines, and antimicrobial peptides.7–10 In the lung, Il-17A is released to the site of infection by diverse cell types, such as activated CD4+ memory T cells, T helper (Th)-17 cells, γδ T cells, innate lymphoid cells and macrophages and mediates the recruitment of inflammatory cells, such as neutrophils.4,9,11–16 Studies showed that IL-17A contributes to the clearance of extracellular bacteria in the respiratory tract. Mice with IL-17 receptor deficiency were more susceptible to Klebsiella pneumoniae lung infection. Over-expression of IL-17A in the pulmonary compartment resulted in improved bacterial clearance and the survival of mice infected with K. pneumonia.10,17 The expression of IL-17A was found to be increased in mouse lungs infected with P. aeruginosa.11,18–21 Il-17A-dependent acquired immunity is also required for efficient clearance of Streptococcus pneumoniae in the upper airways.13,16
Recent studies suggest that IL-17A plays a major role in the pathogenesis of acute respiratory distress syndrome (ARDS).4,22 In experimental LPS-induced ARDS, IL-17A released by innate lymphoid cells mediates neutrophilic inflammation and lung pathology. 4
We therefore investigated the role of IL-17A in a mouse model of severe pneumonia using exemplarily Gram-negative and -positive bacteria. We demonstrated that IL-17A mediates the expression of keratinocyte-derived chemokine (KC) and the recruitment of inflammatory cells in mice infected with a sub-lethal dose of P. aeruginosa. IL-17A-deficient mice were protected from lethal P. aeruginosa lung infection. In contrast, the infection with a sub-lethal dose of S. pneumoniae resulted in increased pulmonary bacterial burden and inflammation in IL-17A deficient mice.
Materials and methods
Mouse experiments
The IL-17A-deficient mice (IL-17A–/–, complete knockout, C57BL/6) were a gift from Dr Yoichiro Iwakura (Institute of Medical Science, University of Tokyo, Japan). 23 The IL-17A–/– mice were transferred into a pathogen-free facility through embryo-transfer. Wild-type (WT, C57BL/6) mice were obtained from Janvier Labs (Le Genest-Saint-Isle, France). All animal experiments were approved by the Landesamt für Soziales, Gesundheit und Verbraucherschutz of the State of Saarland in accordance with the national guidelines for animal treatment. Seven- to 9-wk old female WT C57BL/6 and IL-17A-deficient C57BL/6 mice were slightly anesthetized by means of an intraperitoneal (i.p.) injection of 2.6 mg of ketaminhydrochloride (Ketanest; Pfizer, Germany) and 0.18 mg of xylazinhydrochloride (Rompun; Bayer, Germany) per mouse and infected intranasally with viable P. aeruginosa (1 × 106 colony forming units (CFU) and 4 × 106 CFU) or S. pneumoniae (4 × 107 CFU). 6 Two days after the bacterial infection, the mice were euthanized, the tracheae were cannulated and a bronchoalveolar lavage (BAL) was performed with 1 ml of phosphate-buffered saline (PBS) flushed three times into the lungs. 24 The lungs were removed and homogenized in 1 ml PBS. BAL fluid (BALF) was centrifuged at 4℃ to obtain BAL cells and cell-free supernatants. Inflammatory cells were suspended in 1 ml of PBS. Total cell numbers were determined and cytospins were prepared. Macrophages, neutrophils, lymphocytes, and eosinophils were differentiated by light microscopy. P. aeruginosa and S. pneumoniae CFU values were determined by spreading appropriate dilutions of BAL fluids and lung homogenates on agar plates. 5
Bacterial strains
P. aeruginosa strain PAO1 was grown overnight at 37℃ on LB agar plates. Bacterial cells were taken from the plate, re-suspended in LB medium, and incubated for 2-4 h at 37℃. A type 6A clinical isolate of S. pneumoniae (a gift from Dr Jeffrey Weiser, University of Pennsylvania, USA)25,26 was cultured in Todd Hewitt broth at 37℃. After washing with PBS, bacteria were adjusted to an optical density of 600 nm (OD600) of 1; they were then diluted with PBS and directly used for the experiments. 5
Determination of cytokine concentrations
The concentrations of the inflammatory cytokines KC and IL-1β were measured using an ELISA with a Tecan Ultra 384 ELISA reader and the software Magellan (Tecan, Germany). All ELISA kits were purchased from R&D Systems (UK) and used as instructed by the manufacturer. Protein concentrations were determined using the Pierce BCA-protein assay (Thermo Fisher Scientific Inc, Rockford, IL, USA).
Statistical analysis
Comparisons between two groups were analyzed by the Student’s t-test (two-sided) or Mann-Whitney when data were not normally distributed. Values for mouse experiments are displayed as mean ± SEM. Survival data were compared using a Fisher exact test and a log-rank (Mantel-Cox) test. Results were considered statistically significant for P < 0.05. All statistical tests were performed using the software Prism (GraphPad Software, San Diego, CA, USA).
Results
Il-17A deficiency protects mice from lethal P. aeruginosa infection
It has been shown that excessive inflammation results in lung pathology and reduced bacterial clearance during acute P. aeruginosa pneumonia.2,5 To evaluate the role of IL-17A in the outcome of early pneumonia, WT and IL-17A–/– mice were intranasally infected with a lethal dose (4 × 106 CFU) and sub-lethal dose (1 × 106 CFU) of P. aeruginosa. Mortality was significantly reduced in IL-17A–/– mice infected with a lethal dose compared with WT mice infected with a lethal dose 48 h post-infection (Figure 1a). Figure 1b shows that the bacteria were efficiently cleared from the lungs of mice infected with a sub-lethal dose of P. aeruginosa. However, numbers of bacteria were significantly reduced in the lungs of IL-17A–/– mice.
IL-17A deficiency is protective during P. aeruginosa pneumonia. (a) Survival of WT and IL-17A–/– mice infected with 4 × 106 (lethal dose) CFU or 1 × 106 CFU (sub-lethal dose) of P. aeruginosa for 48 h (n = 16 per group for lethal dose, n = 7 for the sub-lethal dose). *P < 0.05 as compared to WT mice (lethal dose). (n) Numbers of viable bacteria in lung tissue (homogenate) of mice infected with 1 × 106 CFU of P. aeruginosa for 48 h. Data are shown as mean ± SEM. Bars indicate significant differences of *P < 0.05, (n = 7 per group). WT: wild-type, CFU: colony forming units.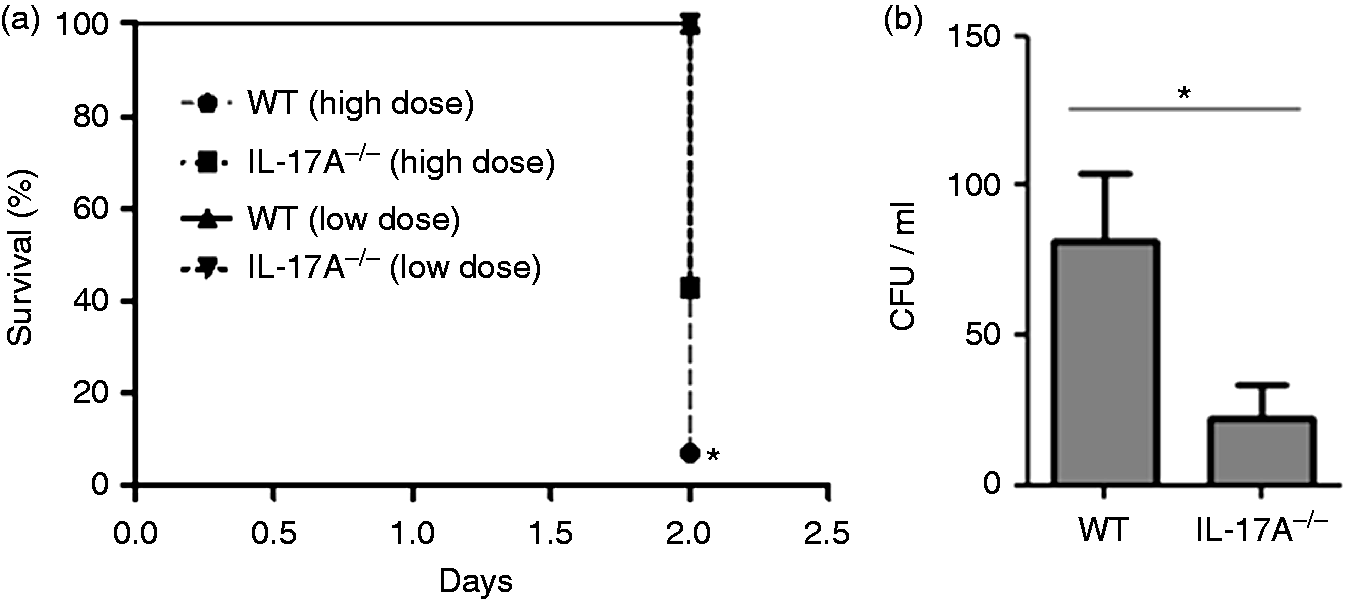
IL-17A mediates P. aeruginosa-induced inflammation
We further analyzed pulmonary inflammation in mice infected with 1 × 106 CFU after 48 h post-infection (sub-lethal dose). IL-17A deficiency associated with a reduced influx of inflammatory cells into the lung. The numbers of total inflammatory cells and the numbers of neutrophils were reduced in BALF of IL-17A–/– mice (Figure 2(a) and (b)). Moreover, concentrations of KC (Figure 2(c)) were significantly reduced in lung tissue obtained from IL-17A–/– mice. However, there was no difference in the concentration of IL-1β in lung tissues of WT and IL-17A–/– mice (Figure 2(d)). KC and IL-1β were below the detection limit (15.6 pg/ml) in BALF of WT and IL-17A–/– mice. Total protein concentrations were significantly reduced in BALF of IL-17A–/– mice (Figure 2(e)).
IL-17A mediates pulmonary inflammation during P. aeruginosa pneumonia. Mice were infected with 1 × 106 CFU of P. aeruginosa for 48 h. (a) Numbers of total inflammatory cells and (b) neutrophils, macrophages and lymphocytes were determined in BALF. The lung homogenate was resolved in 1 ml and concentrations of (c) KC and (d) IL-1β were measured. (e) Total protein concentrations were measured in BALF. Data are shown as mean ± SEM. Bars indicate significant differences of *P < 0.05 (n = 7 per group). CFU: colony forming units; BALF: bronchoalveolar lavage fluid; KC: keratinocyte-derived chemokine.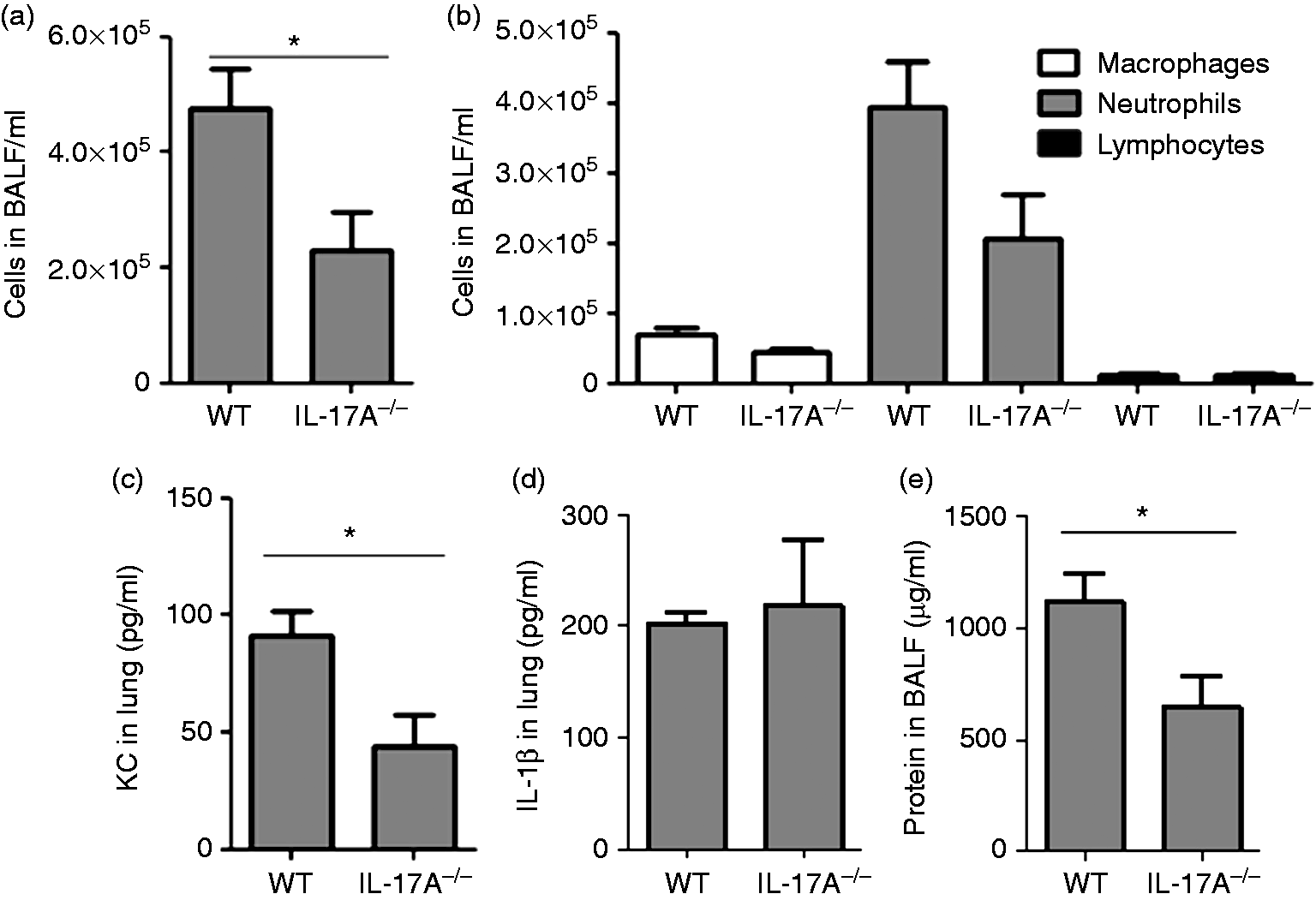
Il-17A is required for clearance of S. pneumoniae
We further evaluated whether IL-17A contributes to the clearance of S. pneumoniae during acute pneumonia. Therefore, WT and IL-17A–/– mice were intranasally infected with a sub-lethal dose of S. pneumoniae (4 × 107 CFU) and analyzed after 48 h. The number of bacteria was increased in BALF (Figure 3(a)) and lungs (Figure 3(b)) obtained from IL-17A–/– mice. The number of total inflammatory cells and neutrophils was increased in BALF of IL-17A–/– mice (Figure 3(c) and (d)). However, contrary to the situation observed in P. aeruginosa-infected mice, there was no difference in the concentrations of KC in lung tissue between WT and IL-17A–/– mice (Figure 3(e)), while concentrations of IL-1β were significantly increased in lung tissue obtained from IL-17A–/– mice (Figure 3(f)). Additionally, total protein concentrations were significantly increased in BALF of IL-17A–/– mice (Figure 3(g)).
IL-17A is required for clearance of S. pneumoniae. Mice were infected with 4 × 107 CFU of S. pneumoniae for 48 h. The number of viable bacteria was determined (a) in lung tissue and (b) BALF. (c) The number of total inflammatory cells and (d) neutrophils, macrophages and lymphocytes was determined in BALF. The lung homogenate was resolved in 1 ml and concentrations of (e) KC and (f) IL-1β were measured. (g) Total protein concentrations were measured in BALF. Data are shown as mean ± SEM. Bars indicate significant differences of *P < 0.05 (n = 7 per group). CFU: colony forming units; BALF: bronchoalveolar lavage fluid; KC: keratinocyte-derived chemokine.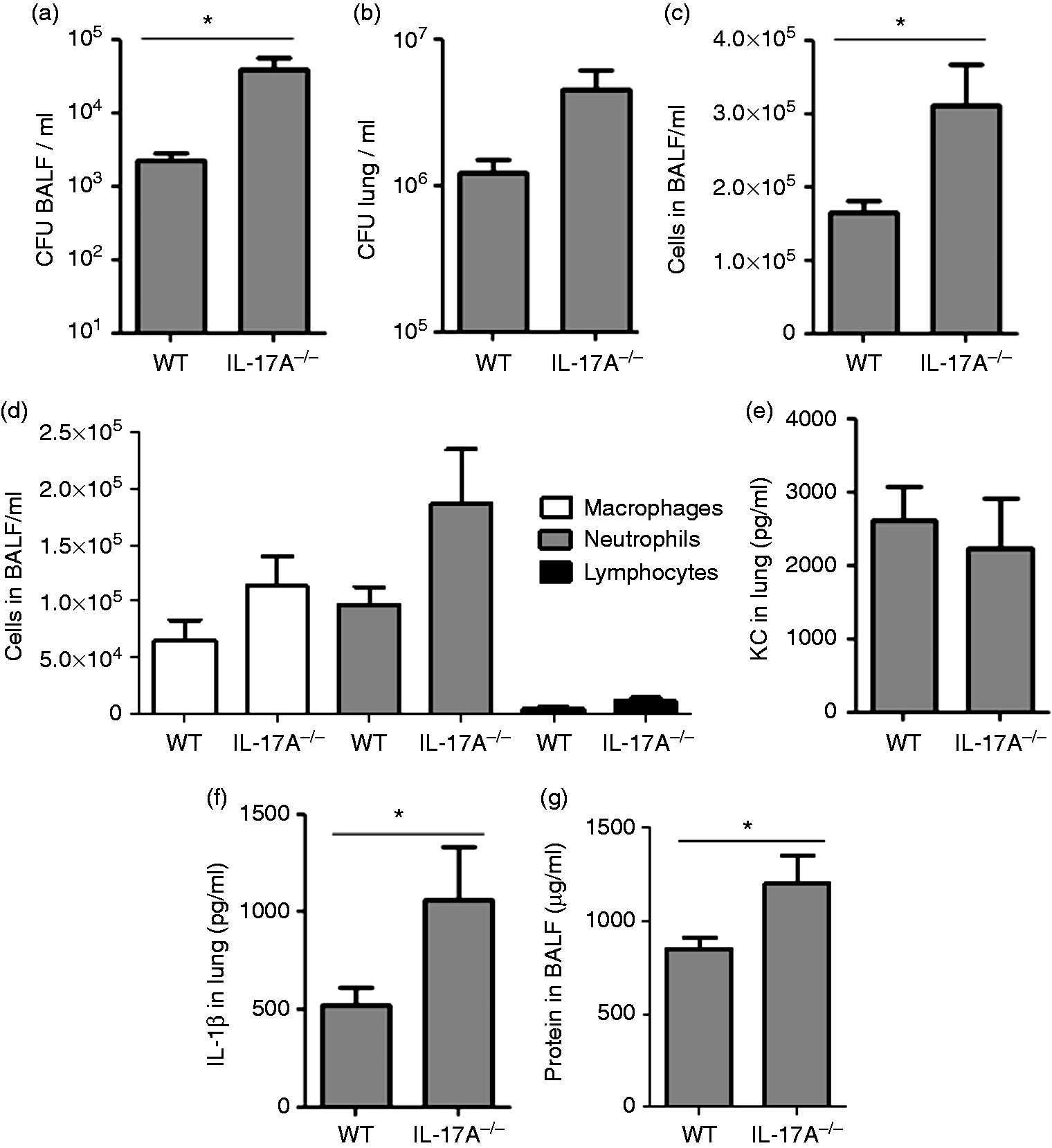
Discussion
Here, we describe a differential role of IL-17A during acute bacterial pneumonia. We demonstrate that IL-17A deficiency protects mice from lethal P. aeruginosa lung infection. We further show that IL-17A is required for the pulmonary expression of the chemokine KC and the recruitment of inflammatory cells into the lung in mice infected with a sub-lethal dose of P. aeruginosa. IL-17A deficiency results, contrary thereto, in increased bacterial burden associated with increased inflammation in the lungs of mice infected with a sub-lethal dose of S. pneumoniae.
Innate and adaptive immunity is required to keep the lung almost completely sterile and free of pathogens. 1 In the case of microbial infection, pro- and anti-inflammatory signals need to be balanced for the clearance of pathogens and avoidance of immunopathology. 27 However, overwhelming inflammation often leads to the pathology of pneumonia. Murine models have shown that P. aeruginosa-induced lung pathology is associated with decreased bacterial clearance and increased mortality.2,5,28,29 Thus, it is suggested that inhibition of inflammatory responses is beneficial during acute and chronic P. aeruginosa lung infections.2,4,5 Indeed, studies have shown that inhibition of the central inflammatory mediator IL-1β or signaling cascades required for the activation of IL-1β results in reduced pathology and enhanced bacterial clearance independent of neutrophilic inflammation in acute experimental pneumonia.2,5,28 Our experiments with viable P. aeruginosa provide evidence that reduced neutrophilic inflammation in IL-17A-deficient mice is beneficial during P. aeruginosa infection. Importantly, decreased pulmonary neutrophilic inflammation in IL-17A-deficient mice was not associated with impaired clearance of P. aeruginosa. Both WT and IL-17A-deficient mice efficiently cleared the bacteria within 48 hours after infection with a sub-lethal bacterial dose. Thus, the decreased inflammatory response in IL-17A-deficient mice resulted in a sufficient recruitment of neutrophils required for the clearance of P. aeruginosa from the lung. 30 Our observation that IL-17A-deficient mice were protected from mortality in lethal P. aeruginosa pneumonia rather suggests that IL-17A is a central mediator of P. aeruginosa-induced overwhelming neutrophilic inflammation that results in fatal pathology. Concentrations of total proteins were significantly reduced in BALF obtained from P. aeruginosa infected IL-17A-deficient mice, indicating that IL-17A-mediated pulmonary inflammation results in lung tissue lesions. Our results are in line with studies showing that neutralization of IL-17A weakens LPS-induced lesions of lung tissue.22,31,32 Furthermore, neutrophilic inflammation was reduced in IL-17A-deficient mice challenged with LPS or P. aeruginosa. 4 Concentrations of KC were significantly reduced in the lungs of IL-17A-deficient mice, whereas concentrations of IL-1β were not affected. Thus, P. aeruginosa IL-17A seems to perpetuate inflammation by inducing the expression of chemokines that direct neutrophils into the lung. As concentrations of IL-1β were not affected in IL-17A-deficient mice, IL-17A seems to function independently or downstream of IL-1β in our infection model. This assumption is supported by findings that IL-1β mediates IL-17A production from neutrophils in inflamed tissue. 33
Importantly, the effect of IL-17A blockade might be specific for P. aeruginosa lung infections. In this study, containment of S. pneumoniae was reduced in the lungs of IL-17A-deficient mice. This was observed in a similar way in a report on Bordetella pertussis infection in which the bacterial load was enhanced in IL-17-defective mice. 34 Moreover, murine studies have shown that IL-17A-mediated inflammation plays a role in the host defense against Gram-negative K. pneumonia.10,17 As seen for IL-17A, studies have shown that the reduction or inhibition of IL-1β signalling results in an increased susceptibility to lung infections with S. pneumonia.5,35–37 Thus, in case of lung infections with Gram-negative or -positive bacteria other than P. aeruginosa, intervention in inflammatory responses by targeting IL-17A- or IL1β-signaling negatively affects the outcome of pneumonia. This observation raises the question as to whether the inverse role of IL-17A in P. aeruginosa—as compared to other bacteria—depends on bacterial behavior and components, on the specific host reaction or on both.
In conclusion, we show that IL-17A has a differential function during severe pneumonia. An IL-17A-dependent host defense might be necessary to kill and clear streptococci and numerous Gram-negative bacteria, whereas IL-17A-mediated overwhelming host reaction or “hyper-inflammation” appears to be detrimental in infections with P. aeruginosa. It has been reported that inhibition of IL-17A is beneficial in patients with psoriatic arthritis. 38 This opens the perspective for IL-17A as a therapeutic target in other inflammatory or infectious diseases. The reduction of pulmonary inflammation through the targeting of IL-17A may be beneficial during acute P. aeruginosa pneumonia in patients.
Footnotes
Acknowledgments
We thank Andreas Kamyschnikow (Saarland University) for excellent technical assistance. We thank Ann Soether and Tina Wiesen-Philipps (Saarland University) for careful reading of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Deutsche Forschungsgemeinschaft (DFG) and the Homburger Forschungsförderungsprogram (HOMFOR) 2014 to Christoph Beisswenger (grant numbers DFG BE 4813/1-1 and DFG Ba 1641/12).