
Abstract
Cancer pain management is a challenge for which Chinese herbal medicine might be useful. To study the spinal mechanisms of the Chinese medicated gel Long-Teng-Tong-Luo (LTTL), a 7-herb compound, on bone cancer pain, a bone cancer pain model was made by inoculating the tibias of female rats with Walker 256 cells. LTTL gel or inert gel, 0.5 g/cm2/d, was applied to the skin of tumor-bearing tibias for 21 days beginning a day after the inoculation. Mechanical threshold and paw withdrawal latency to thermal stimulation was measured. Transient receptor potential (TRP) cation channels in lumbar dorsal root ganglia (DRG) were immunostained and counted, and lumbar spinal cord interleukin-17A (IL-17A) was measured with real-time polymerase chain reaction and enzyme-linked immunosorbent assay. TRP antagonists and interleukin (IL)-17A antibodies were intrathecally administered to determine their effects on bone cancer pain. The gel significantly (P < .05) alleviated cancer-induced mechanical allodynia and thermal hyperalgesia and inhibited cancer-enhanced expression of IL-17A in spinal astrocytes and the TRP subfamily members V1, A1, and V4 in lumbar DRG. Intrathecal TRP antagonists at 10 µg significantly (P < .05) attenuated mechanical allodynia, thermal hyperalgesia, and IL-17A expression, indicating that TRP channels facilitate spinal IL-17 expression and cancer pain. IL-17A antibodies inhibited cancer pain, suggesting that IL-17A promotes such pain. The data show that LTTL gel inhibits cancer pain, and this might be accounted for by the decrease in expression of DRG TRP channels and spinal astrocyte IL-17A.
Background
Cancer pain management remains a challenge. Although opioids manage pain, their use is limited by adverse effects.1,2 A reported 41% to 62% of cancer patients use herbs as complementary or alternative therapy to relieve symptoms and improve their quality of life,3,4 and clinical observation suggests that Chinese herbal medicine might be useful for these purposes.5,6
Metastatic bone tumors, considered to be the most common cause of cancer-related pain,7,8 are particularly common in patients with breast cancer. Annually, about 400000 US citizens are affected by skeletal metastases. 9 We recently developed an analgesic herbal gel, Long-Teng-Tong-Luo (LTTL), based on San Qi He Bi Zhen, a classical formula for pain management. Previous studies show that LTTL significantly alleviated bone cancer-induced mechanical allodynia and thermal hyperalgesia when applied to bone cancer sites in rats 10 ; however, the mechanisms were unclear. The aim of this study was to explore spinal mechanisms of LTTL gel on bone cancer pain in a rat model.
It has been reported that the transient receptor potential cation (TRP) channel subfamily V member 1 (TRPV1) was upregulated in dorsal root ganglia (DRG) in a bone cancer pain model 11 and that a subcutaneous TRPV1 antagonist or a disruption of the TRPV1 gene significantly attenuated both ongoing and movement-evoked nocifensive behaviors in this model. 12 In other previous studies, mice lacking TRPA1 or TRPV4 were shown to be less sensitive to painful mechanical stimuli, 13 intrathecal (i.t.) administration of a TRPA1 channel antagonist markedly attenuated mechanical sensitivity in a streptozotocin-induced rat model of diabetes mellitus, 14 and i.t. administration of a TRPV4 antagonist significantly inhibited thermal hyperalgesia in rats with chronic DRG compression. 15 Because these data indicate that these 2 channels have a role in neuropathic pain, we hypothesized that TRPV1, TRPA1, and TRPV4 are also involved in bone cancer pain, that they would be upregulated in DRG neurons in a bone cancer pain model, and that LTTL treatment would decrease this upregulation to inhibit the pain.
It has also been reported that pro-inflammatory spinal cytokines such as interleukin-1β (IL-1β), IL-6, and tumor necrosis factor-α are involved in bone cancer pain. 16 The IL-17 cytokine family has recently been reported to be a critical player in pain modulation. A study shows that spinal astrocytes express IL-17A, peripheral hind paw inflammation induces upregulation of spinal IL-17A, and i.t. antibodies against IL-17A increase hind paw withdrawal latency in rats with hind paw inflammation. 17 Those data indicate that spinal IL-17A facilitates inflammatory pain. We hypothesized that it also plays a role in bone cancer pain and that LTTL would decrease bone cancer–induced upregulation of this cytokine to alleviate bone cancer pain.
Materials and Methods
Herbal Preparation and Extraction
Long-Teng-Tong-Luo gel, originally called Tong-Luo-San-Jie, is composed of 7 herbs (Table 1). The herbs were purchased from Huayu Pharmaceutical Company (Shanghai, China), which retains a complete set of samples of the authenticated herbs, and were prepared traditionally. 18 The active ingredients were extracted and the LTTL gel was prepared as previously described. 10 The chemical profile of the formulated gel was established by ultraperformance liquid chromatography analysis as previously reported. 10
The 7 Herbs in Long-Teng-Tong-Luo (LTTL) Gel.

Animals
Female Sprague-Dawley rats weighing 150 to 170 g (Shanghai Slac Laboratory Animal Co, Ltd, Shanghai, China) were kept under controlled environmental conditions (24°C ± 0.5°C relative humidity 50% to 60%; 7
Experimental Design
Six sets of experiments were carried out: (1) effect of LTTL on cancer-induced pain; (2) cancer and LTTL modulation of TRPV1, TRPA1, and TRPV4 in lumbar DRG; (3) effect of i.t. TRPV1, TRPA1, and TRPV4 antagonists on cancer-induced pain; (4) cancer and LTTL modulation of spinal cord IL-17A; (5) effect of i.t. IL-17A antibodies on cancer-induced pain; and (6) effect of TRP channel antagonists on spinal IL-17A.
In Experiment 1, 30 rats were randomly divided into 3 groups (n = 10/group): (1) sham—vehicle (phosphate buffered saline; PBS) inoculation into the tibia plus topical administration of inert gel; (2) vehicle—Walker 256 cell inoculation plus topical administration of inert gel; and (3) treatment—Walker 256 cell inoculation plus topical administration of LTTL gel at 0.5 g/cm2/d. LTTL gel (treatment group) or inert gel (vehicle and sham groups) was evenly applied to the skin of tumor-bearing tibias twice a day at 8
In Experiment 2, 45 rats were randomly divided into sham, vehicle, and treatment groups as in experiment 1. Each group was subdivided into 3 groups, and TRPV1, TRPA1, and TRPV4 were measured in lumbar 4-5 (L4-5) DRG using immunofluorescence respectively on day 7, 14, or 21 (n = 5/time point).
In Experiment 3, to investigate the effect of TRPV1 antagonist on cancer pain, cancer-bearing rats were divided into 3 groups (n = 8/group): (1) vehicle, dimethyl sulfoxide (DMSO); (2) 2 µg TRPV1 antagonist, 5′-iodo-resiniferatoxin (I-RTX, Sigma Technology Inc, Ltd, St Louis, MO, USA); and (3) 10 µg I-RTX. The antagonist was dissolved in DMSO and injected i.t. 17 to 20 days after Walker 256 carcinoma cell inoculation. Thermal hyperalgesia was assessed by measuring paw withdrawal latency (PWL) before (day 0) and after surgery on days 4, 10, 18, and 20. Mechanical threshold was tested on days 0, 5, 11, 17, and 19. On days 17 to 20, behavioral tests were performed 20 minutes after the I-RTX application. The rats of the other 2 sets were respectively given the TRPA1 antagonist HC-030031 (Sigma Technology Inc, Ltd, St Louis, MO, USA) or gadolinium, a blocker of cation channels including the TRPV4 (Tocris Bioscience, Washington, DC, USA) and similarly tested.
In Experiment 4, 72 rats were divided into sham, vehicle, or treatment groups and euthanized on days 7, 14, or 21 postinoculation (n = 8/group/time point) to measure IL-17A mRNA. The L4-5 spinal cord was removed, frozen on dry ice, and stored at −80°C. The RNA was extracted for IL-17A mRNA analysis using real-time polymerase chain reaction (RT-PCR) at each time point.
In Experiment 5, cancer-bearing rats were divided into three groups (n = 8/group), saline and 2 and 20 µg of IL-17A antibody (Boiss Bio-technology Co, Ltd, Beijing, China) to investigate the effect of IL-17A on cancer pain. The antibody was administered i.t. on 17 to 20 days post–cancer cell inoculation; mechanical allodynia and thermal hyperalgesia were measured at the time points used in experiment 3.
In Experiment 6, the effects of antagonists of the three TRP channels on spinal IL-17A were studied using the rats of experiment 3. On day 21, their spinal cords, treated with 10 µg of a TRPV1, TRPA1, or TRPV4 antagonist, were removed to measure IL-17A using enzyme-linked immunosorbent assay (ELISA). DMSO-treated cancer-bearing rats and sham rats without cancer were used as control (n = 8). The cell type that produces IL-17A was determined by double immunofluorescence of IL-17A and the cell markers glial fibrillary acidic protein (GFAP), OX-42, or NeuN respectively for astrocytes, microglia, and neurons.
Induction of Bone Cancer
Preparation of Cells
Walker 256 cells were prepared as previously described. 10 Briefly, 1 × 107 Walker cells/mL (70-90 g) were injected into the abdominal cavities of the rats. One week later, ascitic fluid was extracted; the cells were harvested, washed with PBS, resuspended in a concentration of 3.5 × 105/6 μL PBS, and placed on ice until implantation.
Implantation of Cancer Cells
The rats (150-170 g) were anesthetized with 10% chloral hydrate (200 mg/kg, intraperitoneal). The bone cancer pain model was prepared according to methods previously described. 10 In brief, after shaving the hair and disinfecting the skin with 75% v/v ethanol, we made superficial lateral incisions in the skin covering the patella. At the intercondylar eminence 7 mm below the knee joint, a 23-gauge needle was inserted into the medullary cavity of the tibia. The needle was replaced by a 10 μL microinjection syringe to inject either carcinoma cells (3.5 × 105) in 6 μL PBS or 6 μL PBS alone slowly into the right tibial cavity. After the syringe was removed, the injection site was sealed with bone wax and the wound was closed with a gelatin sponge. The animals were returned to their home cages after recovering from anesthesia.
Thermal Hyperalgesia (PWL)
Thermal hyperalgesia was assessed with PWL using a previously described method. 19 For 3 days before the test, rats were allowed to acclimatize for 30 minutes each day inside a 25 cm × 15 cm × 15 cm plastic chamber on the glass surface of a Full-Automatic Plantar Analgesia Tester (BME-410C, Chinese Academy of Medical Sciences, Institute of Biomedical Engineering, China). The glass surface was maintained at 30°C during acclimation and testing. PWL was assessed at the time points described in the experimental design. Mean PWL was determined by averaging the latency of 4 tests. There was a 5-minute interval between each test, and a 25-second cutoff was used to prevent tissue damage. The investigator who measure PWL was blind to group assignment.
Mechanical Allodynia (von Frey Filaments)
Hind paw mechanical allodynia was assessed as previously described.10,20 Testing was performed at the time points detailed in the experimental design with a set of von Frey filaments, hair-like instruments of logarithmically incremental stiffness (0.40, 0.60, 1.4, 2.0, 4.0, 6.0, 8.0, and 15.0 g; Stoelting, Wood Dale, IL, USA). First, the 2.0 g filament was applied perpendicularly to the mid-plantar surface of the hind paw and held 1 to 2 seconds. If the response was positive, defined as withdrawal of the paw on stimulus, the next lower von Frey filament was applied after a 10-minute delay; if negative, the next higher filament was applied. Five stimuli were applied after the first response, and the pattern of response was converted to 50% von Frey threshold. The investigator who measure mechanical allodynia was blind to group assignment.
Intrathecal Injection
Lumbar punctures were performed as previously described. 21 An injection catheter was made by stretching a PE10 polyethylene tube (Clay Adams) to 150% of its original length in 70°C water. One end of the catheter was connected to another 10-cm PE10 tube with a 29-gauge needle and then to a 50 μL glass Hamilton syringe with a PE50 tube. Under isoflurane anesthesia, the dorsal pelvic area was shaved and prepared with 70% alcohol. A sterile 21-gauge needle was inserted between L5 and L6. The catheter, prefilled with 10 μL of drug or vehicle separated from 5 μL of saline by a small air bubble, was inserted into the guide needle and rostrally advanced 4 cm from the tip of the needle into the lumber enlargement; its arrival was confirmed by a tail twitch. The drug or vehicle was injected and followed by a saline flush. The catheter and needle were removed from the intervertebral space 3 minutes postinjection. The rats recovered from anesthesia 2 minutes later. The investigator who did the injection was blind to injected drug and group assignment.
Immunofluorescence
Rats were anesthetized with 10% chloral hydrate (200 mg/kg, intraperitoneal) and immediately perfused transcardially with 4% paraformaldehyde (Sigma) in 0.1 M phosphate buffer (PB) at pH 7.4. L4-5 DRG and the spinal cord were removed, immersed in the same fixative for 2 hours at 4°C, and transferred to 30% sucrose (w/v) in PBS overnight for cryoprotection. Thirty-micrometer-thick sections of DRG were cut on a cryostat. Every fifth section was rinsed in PBS, blocked in PBS with 10% normal goat serum for 60 minutes, and incubated overnight at room temperature with rabbit polyclonal antibodies against TRPV1 (1:200, Boiss Bio-technology Co, Ltd, Beijing, China), TRPA1 (1:200, Boiss Bio-technology Co, Ltd, Beijing, China), or TRPV4 (1:200, Boiss Bio-technology Co, Ltd, Beijing, China). After three 10-minute washings in PBS, sections were incubated in a FITC-conjugated goat anti-rabbit (1:500, Boiss Bio-technology Co, Ltd, Beijing, China) for 60 minutes at room temperature. Control sections were similarly processed except that the primary antisera were omitted. The stained sections were mounted on gelatin-coated slides, coverslipped with antifluorescence quenching liquid (Beyotime Institute of Biotechnology, Shanghai, China), and examined under a Lecia DM2500 fluorescence microscope.
TRPV1-, TRPA1-, and TRPV4-immunostained cells were magnified with a 20× objective lens. Five randomly selected sections from each rat were counted. The results were averaged for each rat and then for the group.
For double immunofluorescence of IL-17A/GFAP, IL-17A/OX-42, and IL-17A/NeuN, the spinal cord section was incubated in a mixture of rabbit polyclonal antibodies against IL-17 (1:50, Boiss Bio-technology Co, Ltd, Beijing, China) and mouse monoclonal antibodies against GFAP (1:500, Beyotime Institute of Biotechnology, Shanghai, China), OX-42 (1:500, Abcam Co, Cambridge, UK), or NeuN (1:500, Abcam Co, Cambridge, UK). After three 10-minute washings in PBS, sections were incubated in a mixture of FITC-conjugated goat anti-rabbit (1:500, Boiss Bio-technology Co, Ltd, Beijing, China) and CY3-congugated goat anti-mouse (1:500, Boiss Bio-technology Co, Ltd, Beijing, China) for 1 hour at room temperature. Control sections were similarly processed, except that the primary antisera were omitted. The stained sections were mounted on gelatin-coated slides and observed as above. The investigator who did the analysis of immunestaining, RT-PCR and ELISA data was blind to group assignment.
RT-PCR of IL-17A mRNA
For RT-PCR, the spinal cords were removed 4 hours post–herbal treatment on days 7, 14, and 21. The dorsal horns were separated into contralateral and ipsilateral portions and immediately stored in liquid nitrogen. Total RNA was extracted with Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the standard protocol. Three micrograms of total RNA from each sample was used for reverse transcription to get cDNA. IL-17A PCR primers were as follows: forward, gaagttggaccaccacatga; reverse, tccctcttcaggaccaggat. Hydroxymethylbilane synthase forward primer tccctgaaggatgtgcctac and reverse primer acaagggttttcccgtttg were used for internal control. Quantitative PCR was performed in SYBR Premix Ex TaqTM II (TaKaRa Bio Co, Otsu, Japan). Briefly, each well was loaded with 10 µL of Syb Green mix, 0.4 µL of IL-17A primer, 0.4 µL of hydroxymethylbilane synthase primer, 1 µL cDNA, and 8.2 µL ddH2O. Thermal cycling was performed using an Applied Biosystems 7900HT system (ABI, Applied Biosystems, Inc, Carlsbad, CA, USA) with 2-minute denaturation at 95°C followed by 45 cycles of 95°C for 15 seconds, 58°C for 20 seconds, and 72°C for 20 seconds.
RT-PCR data from L4-5 sections of the ipsilateral spinal cord were analyzed using comparative 2-ΔΔCT CT to find the difference in IL-17A mRNA expression between cancer-bearing rats and control. Each experiment sample was analyzed in triplicate.
Detection of Spinal IL-17A After Intrathecal Administration of TRP Channel Antagonists
TRP channel antagonists were administered i.t. on days 17 to 20. On day 21, the spinal cord was harvested and the ipsilateral and contralateral dorsal horns were separated. Ipsilateral spinal cord IL-17A was measured using commercial ELISA kits (Abcam Co, Cambridge, UK) according to the manufacturer’s instructions.
Statistical Analysis
Behavioral data were expressed as mean ± standard deviation (SD) and analyzed with repeated-measures analysis of variance. RT-PCR, immunofluorescence, and ELISA data were analyzed with 1-way analysis of variance followed by a Newman-Keuls test using SPSS 13.0 statistical software. Statistical significance in all was considered to be P < .05.
Results
LTTL Inhibition of Mechanical Allodynia and Thermal Hyperalgesia
First, mechanical withdrawal threshold of the paw ipsilateral to surgery was unchanged over the 21 days in sham rats without cancer cell inoculation. However, the threshold in vehicle rats with cancer cell–inoculation and vehicle treatment gradually decreased and was significantly (P < .05) lower on days 11 to 21 than that in sham rats (Figure 1A). This indicates that the cancer induced mechanical allodynia in ipsilateral hind paws. Second, the threshold in LTTL-treated rats significantly (P < .01) increased compared with that in the vehicle group. The data demonstrate that LTTL treatment alleviated bone cancer–induced mechanical allodynia. Contralateral hind paws of all 3 groups of rats showed no significant mechanical threshold change.

(A, B) Effect of Long-Teng-Tong-Luo (LTTL) gel on mechanical threshold (A) and hind paw withdrawal latency (B). Mean ± SD; n = 10 per group. The gel was given on days 1 to 21 after cancer cell implantation into the tibia. (A) Mechanical threshold was measured at baseline and on days 5, 11, 17, and 21. Cancer induced a significant decrease in mechanical threshold of the hind paw ipsilateral to the cancer-inoculated tibia; LTTL gel treatment significantly restored it compared to inert gel control. Contralateral mechanical threshold showed no change. **P < .01 versus sham; #P < .05; ##P < .01 versus vehicle. (B) Paw withdrawal latency (PWL) was measured at baseline and on days 4, 10, 16, and 20. Cancer significantly shortened PWL of the hind paw ipsilateral to the cancer cell–inoculated tibia. LTTL significantly restored PWL compared with inert gel control. Contralateral paws showed no change in PWL. **P < .01 versus sham; ##P < .01 versus vehicle.
Figure 1B shows the effect of LTTL gel on PWL. PWL of the ipsilateral hind paw was significantly (P < .01) lower on days 10 to 20 in vehicle rats with cancer cell inoculation and vehicle treatment than that in sham rats without cancer cell inoculation (P < .01), indicating that bone cancer induced thermal hyperalgesia in the hind paw ipsilateral to the cancer-inoculated tibia. LTTL treatment significantly (P < .01) increased PWL on days 16 to 20 compared with vehicle control, showing that the gel alleviated cancer-induced thermal hyperalgesia (Figure 1B). Contralateral hind paws showed no significant PWL change.
LTTL Inhibition of TRP Channel Expression in DRG
As shown in Figure 2A, the number of TRPV1-immunoreactive cells in ipsilateral L4-5 DRG was similar from 7 to 21 days post-PBS inoculation in sham rats. This parallels the behavioral data, which showed no significant mechanical allodynia or thermal hyperalgesia during the same period. However, the number of TRPV1-immunoreactive cells gradually increased from 7 to 21 days in vehicle rats. It was significantly (P < .01) more than that in PBS-inoculated sham rats on days 14 and 21, indicating that the bone cancer induced significant TRPV1 production in ipsilateral DRG. This increased TRPV1 expression was consistent with the significant mechanical allodynia and thermal hyperalgesia on days 14 to 21. Furthermore, comparison between vehicle and LTTL treatment groups showed significantly fewer TRPV1-immunoreactive cells in the LTTL group than in the vehicle group on days 14 (P < .05) and 21 (P < .01), demonstrating that the gel inhibited TRPV1 expression in DRG. The data suggest that LTTL inhibition of TRPV1 expression is related to its inhibition of bone cancer–induced pain.

Effect of Long-Teng-Tong-Luo (LTTL) gel on TRPV1 (A), TRPA1 (B), and TRPV4 (C) immunostaining of ipsilateral lumbar 4-5 dorsal root ganglia (DRG) on days 7, 14, and 21 after Walker 256 cell injection into the tibia. Mean ± SD in A2/B2/C2; n = 5/group. TRPV1, TRPA1, and TRPV4 were detected by immunofluorescence in phosphate buffered saline (PBS)-inoculated sham (a, d, g), vehicle (b, e, h), and treatment (c, f, I) tissues on days 7 (a, b, c), 14 (d, e, f), and 21 (g, h, i). Cancer significantly increased the number of TRPV1-, TRPA1-, and TRPV4-immunopositive cells in DRG on days 14 and 21; LTTL treatment significantly decreased cancer-enhanced immunopositive staining of those 3 TRP channels. **P < .01 versus sham; ##P < .01 versus vehicle; #P < .05 versus vehicle.
As shown in Figure 2, LTTL treatment significantly (P < .01) inhibited bone cancer–enhanced TRPA1 (Figure 2B) and TRPV4 (Figure 2C) expression in ipsilateral L4-5 DRG on day 21 post–cancer cell inoculation compared with vehicle. This indicates that LTTL inhibition of TRPA1 and TRPV4 expression might be involved in its inhibition of bone cancer–induced pain. Control sections without primary antibodies showed no specific staining (data not shown).
TRP Antagonist Inhibition of Mechanical Allodynia and Thermal Hyperalgesia
Mechanical withdrawal threshold of the paw ipsilateral to the cancer cell–inoculated tibia was significantly (P < .01) greater on days 17 and 19 in rats treated with 10 µg of the TRPV1 antagonist I-RTX than in rats treated with DMSO (Figure 3A1). The threshold in rats treated with 2 µg of I-RTX was the same as that in rats treated with DMSO. These data demonstrate that TRPV1 antagonist dosage-dependently inhibits bone cancer–induced mechanical allodynia. I-RTX treatment did not influence mechanical withdrawal threshold of the contralateral hind paw, which remained unchanged from baseline (data not shown).

Effect of a TRPV1 antagonist, I-RTX (A), a TRPA1 antagonist, HC-030031 (B), and a TRPV4 antagonist, gadolinium (C), on mechanical threshold (A1, B1, and C1) and paw withdrawal latency (PWL) (A2, B2, and C2) in cancer cell–inoculated rats. Mean ± SD, n = 8 per group. Antagonists were given (intrathecal) 17 to 20 days post–Walker 256 carcinoma cell inoculation. A1/B1/C1: Mechanical threshold was measured at baseline and on days 5, 11, 17, and 19 after Walker 256 cell injection into the tibia. TRPV1 (A1), TRPA1 (B1), and TRPV4 (C1) antagonists at 10 µg but not 2 µg significantly increased mechanical threshold compared with vehicle on days 17 and 19. A2/B2/C2: PWL was measured at baseline and on days 4, 10, 18, and 20 after Walker 256 cell injection into the tibia. TRPV1 (A2) TRPA1 (B2) and TRPV4 (C2) antagonist at 2 to 10 µg significantly lengthened PWL compared with vehicle on days 18 to 20. **P < .01 versus vehicle; ##P < .01 versus 2 µg antagonist.
Ipsilateral hind paw PWL was significantly (P < .01) greater on days 18 and 20 in rats treated with 10 µg of the TRPV1 antagonist I-RTX compared with that in rats treated with DMSO (Figure 3A2). PWL in rats treated with 2 µg of I-RTX was the same as that in DMSO-treated rats on day 18. These data show that the TRPV1 antagonist significantly inhibits bone cancer–induced thermal hyperalgesia. I-RTX treatment did not influence PWL of the contralateral hind paw, which remained unchanged from baseline (data not shown).
Figure 3B shows the effect of TRPA1 antagonist HC-030031 on mechanical allodynia (Figure 3B1) and thermal hyperalgesia (Figure 3B2). At 10 µg, TRPA1 significantly (P < .01) increased mechanical withdrawal threshold and PWL. This indicates that the antagonist attenuates bone cancer–induced mechanical allodynia and thermal hyperalgesia.
Figure 3C shows the effect of the TRPV4 antagonist gadolinium on mechanical allodynia (Figure 3C1) and thermal hyperalgesia (Figure 3C2). Like the TRPV1 and TRPA1 antagonists, the TRPV4 antagonist significantly increased (P < .01) mechanical withdrawal threshold and PWL at a dosage of 10 µg. These data indicate that the antagonist attenuates bone cancer–induced mechanical allodynia and thermal hyperalgesia.
LTTL Inhibition of IL-17A Expression in the Spinal Cord
RT-PCR analysis showed that IL-17A mRNA levels in the ipsilateral L4-5 spinal cord significantly (P < .01) increased in vehicle- and LTTL gel–treated cancer-bearing rats compared with PBS-inoculated sham rats on days 14 and 21 after cancer cell inoculation (Figure 4). Furthermore, IL-17A mRNA levels were obviously low in LTTL-treated rats compared with that in vehicle-treated rats on days 14 and 21; the difference on day 21 was significant (P < .01). This suggests that LTTL inhibits cancer-enhanced IL-17A expression in the spinal cord.
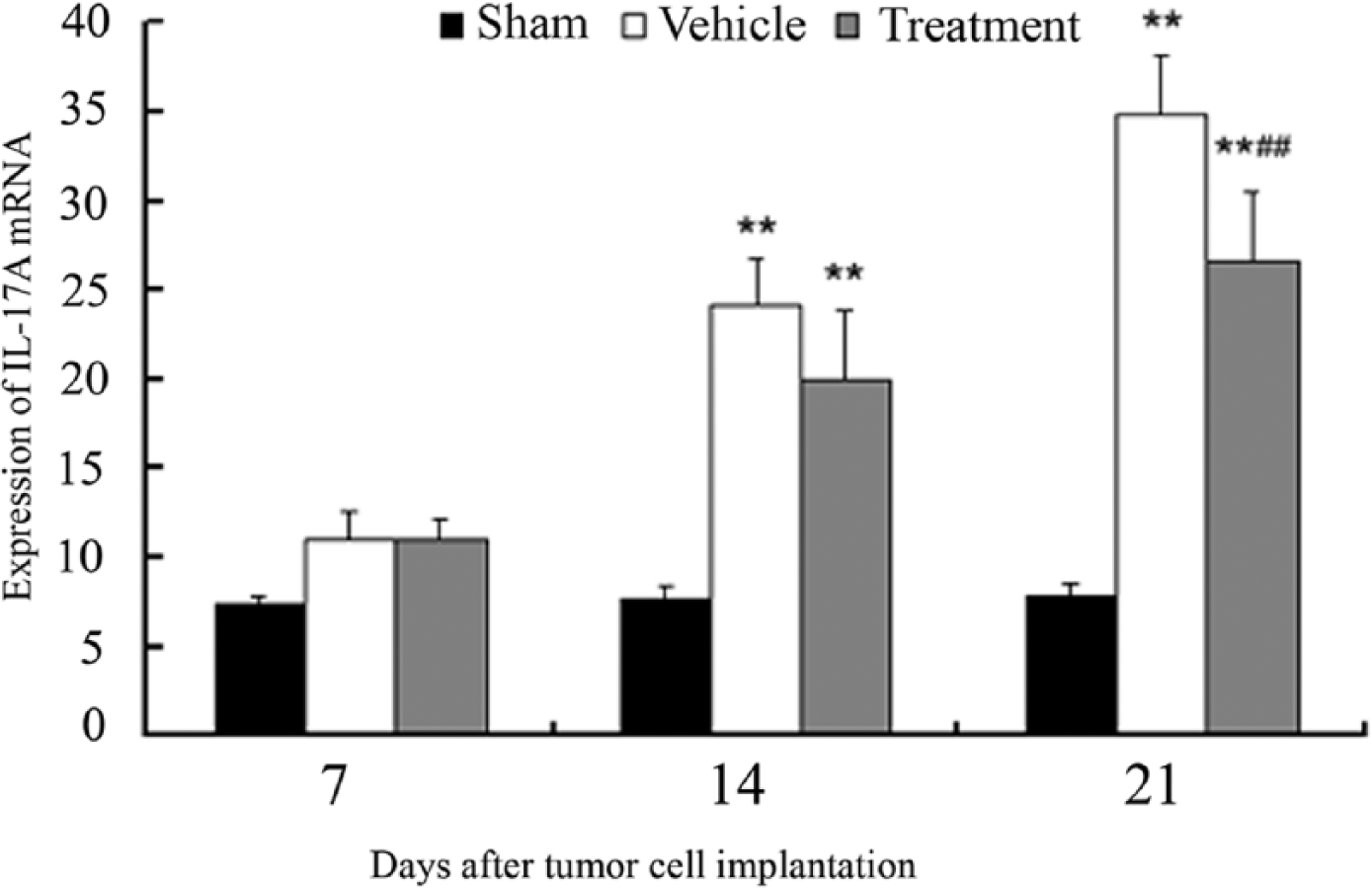
Effect of Long-Teng-Tong-Luo (LTTL) on interleukin (IL)-17A mRNA of the ipsilateral lumbar 4-5 spinal cord on days 7, 14, and 21 after Walker 256 cell injection into the tibia. Mean ± SD, n = 8 per group. IL-17A mRNA was detected by real-time polymerase chain reaction (RT-PCR). Cancer induced a significant increase of IL-17 mRNA in the spinal cord; LTTL treatment significantly decreased it compared with inert gel control on day 21. **P < .01 versus sham; ##P < .01 versus vehicle.
IL-17A Antibody Inhibition of Pain
Figure 5 shows the effect of an i.t. injection of IL-17A antibodies on mechanical allodynia and thermal hyperalgesia. Before the injection, overall mechanical withdrawal threshold means on day 11 and PWL on day 10 were similar in the 3 groups of cancer-bearing rats. Compared with saline injection, 20 µg of i.t. IL-17A antibodies on days 17 to 20 significantly (P < .01) increased mechanical threshold (Figure 5A) on days 17 and 19 and PWL (Figure 5B) on days 18 and 20. At 2 µg, the antibodies had no significant effect on mechanical threshold. These data demonstrate that IL-17A antibodies significantly and dosage dependently inhibit bone cancer–induced mechanical allodynia and thermal hyperalgesia.

Effect of an intrathecal interleukin (IL)-17A antibody on mechanical threshold (A) and paw withdrawl latency (PWL) (B) in cancer cell–inoculated rats 17 to 20 days postinoculation. Mean ± SD, n = 8 per group. (A) Mechanical threshold was measured at baseline and on days 5, 11, 17, and 19 after Walker 256 cell injection into the tibia. IL-17A antibody at 10 µg but not 2 µg significantly increased mechanical threshold compared with vehicle on days 17 and 19. (B) PWL was measured at baseline and on days 4, 10, 18, and 20 after Walker 256 cell injection into the tibia. An IL-17A antibody at 2 to 10 µg significantly lengthened PWL compared with vehicle on days 18 to 20. *P < .05; **P < .01 versus vehicle; ##P < .01 versus 2 µg IL-17 antibody.
TRP Channel Antagonist Inhibition of Spinal IL-17A
As shown in Figure 6, IL-17A levels in the ipsilateral spinal dorsal horn were significantly (P < .01) higher in cancer-bearing rats given i.t. vehicle than in PBS-inoculated sham rats on day 21 after cancer cell inoculation (Figure 6). IL-17A levels were significantly lower (P < .01) on day 21 in rats given 10 µg of the i.t TRPA1 antagonist HC-030031, the TRPV4 antagonist gadolinium, or the TRPV1 antagonist during days 17 to 20 than were levels in rats that received vehicle. These data indicate that TRP channel antagonists significantly hamper bone cancer–enhanced increase of IL-17A proteins.

Effect of TRP channel antagonists on interleukin (IL)-17A protein in the ipsilateral lumbar 4-5 spinal cord on day 21 after Walker 256 cell injection into the tibia. Mean ± SD, n = 5 per group. TRP channel antagonists were given on days 17 to 20. Cancer induced a significant increase of IL-17A protein in the ipsilateral spinal cord. TRP channel antagonists significantly inhibited this increase compared with dimethyl sulfoxide (DMSO) control. *P < .05, **P < .01 versus sham; ##P < .01 versus DMSO.
Double immunofluorescence labeling demonstrated that IL-17A colocalizes with the astrocyte marker GFAP but not the microglia marker OX-42 or the neuronal marker NeuN (Figure 7), showing that IL-17A is produced in astrocytes. Control sections without primary antibodies showed no specific staining (data not shown).

Micrographs show colocalization of interleukin (IL)-17A and glial fibrillary acidic protein (GFAP) in superficial lumbar laminae 21 days after Walker 256 carcinoma cell injection into the tibia. Sections were simultaneously labeled with anti-IL-17A and anti-GFAP, anti-NeuN, or anti-OX-42 antisera. The first column is IL-17 immunostaining (a, b, c, green); the second is immunostaining of three cell markers GFAP, NeuN, and OX-42, respectively, for astrocytes, neurons, and microglia (a*, b*, c*, red); the third is the merged graphs (a**, b**, c**); the fourth is a higher magnification of the white box in column 3. Note that IL-17A is localized in astrocytes (yellow cells), as indicated by arrows (a***). Scale bars = 50 µm.
Discussion
In the present study, topical LTTL application significantly alleviated bone cancer–induced mechanical allodynia and thermal hyperalgesia. This is consistent with previous report 10 and other studies showing that a number of the herbs in LTTL, including Trachelospermum jasminoides, 22 Commiphora myrrha, 23 and Melia toosendan, 24 significantly inhibit visceral pain in animal models and that Boswellia carterii 25 significantly lengthens PWL in an inflammatory pain rat model. Since LTTL has not been applied on a distant site, such as the leg contralateral to cancer cell inoculation, we do not know whether LTTL gel application on non–bone cancer site alleviate bone cancer pain. This warrants further investigation.
LTTL treatment also significantly decreased bone cancer–enhanced TRPV1, TRPA1, and TRPV4 proteins in L4-5 DRG, and TRPV1, TRPA1, and TRPV4 antagonists significantly inhibited bone cancer–induced mechanical allodynia and thermal hyperalgesia. This indicates that LTTL inhibition of TRP channel expression might be related to its inhibitory effect on bone cancer pain.
In previous studies with inflammatory pain models, a TRPV1 antagonist 26 and silencing of TRPV1(+) peripheral axons 27 significantly reduced mechanical and thermal pain elicited by complete Freund’s adjuvant. Those findings demonstrate that TRPV1-expressing neurons in DRG contribute to inflammatory pain. Moreover, disruption of the TRPV1 function significantly attenuated ongoing and movement-evoked nocifensive behaviors in a bone cancer pain model. 12 Our data show that a TRPV1 antagonist inhibits bone cancer-induced mechanical allodynia and thermal hyperalgesia, which suggests that TRPV1-expressing neurons in DRG are involved in cancer-induced mechanical and thermal hypersensitivity. Since genetic downregulation of TRPV1 reduces thermal and mechanical hyperalgesia, 28 LTTL downregulation of TRPV1 might contribute to inhibition of bone cancer pain.
Previous study showed that LTTL inhibits osteoclast activity in cancer-bearing bone. 10 Since osteoclast-mediated bone remodeling produces robust extracellular protons that activate TRPV1 in primary afferent fibers innervating bone, 29 we conclude that this LTTL inhibition consequently blocked TRPV1 activation in primary afferent fibers, leading to downregulation of TRPV1 and pain inhibition. Other bioactive chemicals such as bradykinin, adenosine-5′-triphosphate, nerve growth factor, and prostaglandin E2 can trigger TRPV1.30 -32 It is plausible that LTTL also decreases local concentration of these chemicals to block TRPV1, thus inhibiting pain.
Studies report that TRPA1 channel antagonists markedly attenuate mechanical sensitivity in rodent models of inflammatory, 33 postoperative, 34 and neuropathic pain. 14 An intraplantar injection of the TRPA1 agonist cinnamaldehyde not only significantly decreased mechanical threshold but also significantly shortened PWL from a noxious heat stimulus, 35 indicating that TRPA1 might be involved in both mechanical and thermal sensitivity. In our study, TRPA1 significantly increased in cancer-bearing rats compared to PBS-inoculated rats, and TRPA1 antagonists inhibited both cancer-induced mechanical allodynia and thermal hyperalgesia. This suggests that TRPA1-expressing neurons in DRG are involved in mechanical and thermal sensitivity in cancer pain. Thus, LTTL inhibition of TRPA1 expression might contribute to its inhibition of bone cancer pain.
Studies show that TRPV4 antagonists significantly inhibit thermal hyperalgesia following chronic compression of DRG in rats 15 and abolish chemotherapy (taxol)-induced mechanical hyperalgesia. 36 In our study, TRPV4 significantly increased in cancer-bearing rats compared with PBS-inoculated rats, and TRPV4 antagonists inhibited both cancer-induced mechanical allodynia and thermal hyperalgesia. This indicates that TRPV4-expressing neurons in DRG are involved in mechanical and thermal sensitivity in cancer pain and thus that LTTL inhibition of TRPV4 expression might contribute to its inhibitory effect on bone cancer pain.
In the present study, double-immunostaining showed that spinal IL-17A is expressed in astrocytes but not in microglia and neurons during bone cancer pain. Furthermore, IL-17A expression significantly increased in the spinal cord during cancer pain, and i.t. IL-17A antibodies significantly attenuated bone cancer–induced mechanical allodynia and thermal hyperalgesia, indicating that astrocyte-produced IL-17A might promote cancer pain. Previous studies show that spinal IL-17A facilitates inflammatory 17 and neuropathic pain. 37 Taken together, these studies convincingly show that astrocyte-produced IL-17A facilitates a variety of pain. LTTL treatment significantly decreased cancer-enhanced IL-17A expression, which suggests that LTTL inhibits pain by down-regulating IL-17A expression. Our study also found that TRPV1, TRPA1, and TRPV4 antagonist treatments significantly decrease bone cancer–enhanced IL-17A expression, indicating that TRP channel activation might promote the spinal IL-17A upregulation to facilitate pain. In support, a prior study found that activation of TRPA1 evokes substance P (SP) release from the primary sensory neurons 38 and that SP activates glial cells during pain. 39 Thus, LTTL inhibition of TRP channels may result in decrease of SP release and consequent inhibition of spinal IL-17A to alleviate pain. However, we do not exclude other potential mechanisms of LTTL inhibition of IL-17A.
In conclusion, LTTL treatment significantly alleviates bone cancer-induced mechanical allodynia and thermal hyperalgesia by downregulating TRP channel expression in DRG and IL-17A in spinal astrocytes. Our study is the first to demonstrate that TRPA1 and TRPV4 are involved in cancer pain and that LTTL might downregulate these cancer-enhanced ion channels to alleviate cancer pain. However, mechanisms of LTTL inhibition of TRPA1 and TRPV4 need further investigation.
Footnotes
Acknowledgements
We thank Dr. Lyn for her editorial support.
Authors’ Note
Authors Juyong Wang and Ruixin Zhang contributed equally to this work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Health (No. 1R03TW008375-01A1), the National Natural Science Foundation of China (Nos. 81173225 and 81373622), Shanghai University of Traditional Chinese Medicine 085 Program (No. 085ZY1208), and Longhua Medical Project (No. LYTD-02).