
Abstract
Heat shock proteins (HSPs) are highly chaperonic molecules that give immediate response during any stress, tissue damage or bacterial infections. In the present study, the role of HSPs upon bacterial encounter is studied by applying external heat induction to live Caenorhabditis elegans. Heat shock was observed to increase the life span of wild type C. elegans upon pathogenic encounter, indicating a role of HSPs in bacterial infection and immunity. Similar increase in resistance towards pathogenesis observed in long-lived C. elegans daf-2 mutants and the increase in the lifespan indicated a role for the insulin/IGF-1 signaling (IIS) pathway in HSP-mediated pathogenic resistance. The microscopic observation of C. elegans after external heat induction and sequential exposure of pathogens indicated reduction of egg viability. Results of Real-time PCR and immunoblotting analysis of candidate genes revealed that heat shock and IIS pathways collaborate in the observed pathogenic resistance and further suggested SGK-1 to be the possible factor linking both these pathways. In addition, survival assays carried out using mutants equips us with supporting evidence that HSP and HSF-1 are necessary for the accelerated lifespan of C. elegans. Our findings thus confirm that crosstalk between HSPs and SGK-1 influences C. elegans longevity.
Introduction
Physiological stressors like temperature, UV light and certain environmental cues influence the longevity of every individual. Heat stress plays a vital role in maintaining and activating important regulatory players of higher invertebrates. Heat shock proteins (HSPs), which are activated upon heat stress, have gained much prominence in aging research and are thought to be of importance for longevity and overall maintenance of proteome integrity with advancing age. The most compelling evidence so far has been collected from invertebrate models such as Drosophila and Zebrafish, in which the joint effects of longevity and thermo tolerance were found to be a plausible reason for the increased lifespan of these models.1,2 However, Caenorhabditis elegans, a simple soil nematode, considered a versatile model for aging studies, has not been exploited much for heat stress-related lifespan extension against bacterial pathogens. HSPs are considered quintessential molecules produced under heat stress and certain external stresses in this nematode and are therefore thought of as danger-associated molecular patterns.3–5 One of the remarkable characteristics of HSPs is their immunodominance. However, the fallouts of these molecular chaperones are under the control of heat shock factor-1 (HSF-1). HSF-1 is a transcription factor that communicates with the insulin/IGF-1 signaling (IIS) pathway and thus plays a crucial role in longevity. It works parallel to daf-16 FOXO, a transcriptional factor acting downstream of the IIS pathway.6–8
HSF-1 is activated during heat stress and undergoes a stepwise process that includes oligomerization, post-translational modification and nuclear translocation. These successive events presumably result in the combined effect of HSP expression, producing a heat shock response. 9 A number of reports confirm that alleviation of the IIS pathway activates HSF-1, resulting in the enhanced lifespan of the nematode.6,8 A similar phenomenon has been proven to be the reason for the increased longevity observed in daf-2 mutants. 10 Besides its role in longevity, IIS regulation has also been reported to kindle the response of nematode towards many pathogenic bacteria.11,12
Under heat stress, the chaperoning activity of the HSPs modulates and supports folding of polypeptides during synthesis, prevents protein denaturation or aggregation, and directs proteins for degradation when refolding fails.13,14 It has already been proven that the expression of chaperones varies within different cell types during aging.15,16 However, their total capacity and regulation upon external heat shock and consecutive infection has not been well studied. The loss of HSF-1 activity may also lead to immuno-senescence. However, organisms exposed to mild heat stress become long-lived, suggesting that activation of certain stress responses could be sufficient to extend the lifespan.17,18 Although complete deficit of HSF-1 confers lack of immunity, analysis of nematodes with moderately reduced levels of HSF-1 or other HSPs has revealed a novel function in immunity, as well as aging. The applicability of these observations validates the basic aging research in context with the immune response by associating with the IIS pathway. With this milieu, the present study was taken to investigate the heat shock response and its role in prolonging the lifespan during bacterial encounter. Employing the nematodes, culture-specific differences in chaperoning and regulation of the HSF-1 controlled heat shock response was analyzed with an external induction of heat. The association between the IIS pathway and expression of heat shock genes/proteins during a pathogenic encounter was explored, and our observations provide evidence that HSF-1 confers longevity to C. elegans by boosting the immunity to certain extent.
Materials and methods
Bacterial strains and C. elegans
The bacterial strains Staphylococcus aureus (ATCC 11632) and Proteus mirabilis (ATCC 7002) were obtained from ATCC. Escherichia coli OP50 (laboratory food source of C. elegans), C. elegans WT Bristol N2, daf-2 mutant (e1370), hsf-1 mutant (sy441) and daf-21 mutants [JT6130 (p673)V and PR673(p673)V] were obtained from the Caenorhabditis Genetics Centre (Minnesota, Saint Paul, USA). S. aureus and E. coli were maintained in Luria Bertani medium, while P. mirabilis was maintained in nutrient medium. Both the C. elegans N2 wild type and daf-2 mutants were consistently maintained at 20℃ on nematode growth medium (NGM) seeded with E. coli OP50, as per standard methods. 19 All assays were performed with a constant bacterial inoculum (0.3 OD at 600 nm), i.e 12 × 106 cells/ml for P. mirabilis and 18 × 106 cells/ml for S. aureus.
Nematode lifespan assay
Lifespan assays were performed using synchronized population of nematodes. Synchronization was attained by bleaching the nematodes. Subsequently, the eggs were allowed to develop to the young adult stage. The synchronized population was divided into two sets: one set was given a heat shock for 2 h at 35℃ and a recovery period for 4 h at 20℃; the other set was maintained continuously at 20℃. Twenty numbers of age-synchronized young adult C. elegans daf-2 mutant, hsf-1 mutant and daf-21 mutants from both heat shock-induced and non-induced plates were exposed individually to the pathogens P. mirabilis and S. aureus contained in 24-well microtiter plates. The nematodes (heat shock-induced and non-induced) exposed to laboratory food source E. coli OP50 served as control. The microtiter plates were incubated at 20℃ and the lifespan changes were monitored for every 6 h throughout the assay period. Animals were scored as dead when they ceased pharyngeal pumping and did not respond to prodding with a platinum wire.
Chemotaxis assay
A chemotaxis assay was performed to assess the difference in the food preference of external heat shock-induced and non-induced wild type nematode. Briefly, cultures of pathogen [either P. mirabilis or S. aureus (zone A)] and E. coli OP50 (zone B) were spotted at a distance of 3 cm from the center of NGM plates (90 mm) and the preference of the nematode was monitored. Subsequently, 25 wild type animals from each set of heat shock-induced and non-induced populations were thoroughly washed from E. coli OP50 lawn and placed at the center of the plate. The total number of nematodes that have moved towards each zone was counted every 4 h and the results were plotted with 12 h intervals up to 48 h. The assay included controls in which both zone A and zone B were spotted with OP50 or P. mirabilis or S. aureus.
Microscopic observations
For microscopic visualization, external heat-induced and non-induced nematodes were subsequently infected with P. mirabilis and S. aureus separately for time periods of 12, 24 and 36 h. The infected nematodes were washed three times with M9 buffer and placed in a 1-mM sodium azide solution on a 2% agarose pad for microscopic observations under inverted fluorescent microscope (Nikon, Tokyo, Japan). The phenotypic and lifespan changes were monitored and recorded using an attached digital camera.
Total RNA isolation and real-time PCR analysis
List of primer sequences used in the present study.

Western blotting
To isolate total protein for Western blot analysis, wild type and daf-2 mutant nematodes were induced with heat shock and subsequently exposed to pathogen, as detailed in the nematode lifespan assay. Total protein isolated from nematodes exposed to E. coli OP50 served as the control. For protein extraction, the worm aliquots were subjected to sonication and quantified using Bradford’s method. Protease inhibitor cocktail (Sigma, St. Louis, MO, USA) was added to each sample prior to sonication to prevent protein degradation. The samples were separated by 12% SDS-PAGE. The gel was equilibrated and the protein was transferred to a polyvinylidenedifluoride membrane (Whatman, GE Healthcare, Chicago, IL, USA) using semi-dry blotting apparatus (Amersham, Little Chalfont, UK). 21 Consequently, the membrane carrying the transferred proteins was incubated in blocking solution of 5% defatted skimmed milk powder (HiMedia Laboratories, Mumbai, India) dissolved in Tris-buffered saline containing 0.1% Tween-20 for 3 h at 4℃. The membrane was probed individually with anti-HSF-1, anti-HSP-90 and anti-SGK-1 Abs, and also with anti β-actin as a loading control for 6 h at 4℃, and then in the secondary Ab conjugated with alkaline phosphatase (AP) for 4 h at 4℃. The membrane was subsequently taken for immuno-detection by developing it in 1 × AP buffer solution containing the substrates nitro-blue tetrazolium and 5-bromo-4-chloro-3’-indolyphosphate and left until the band appeared. The quantification and fold change were determined using ImageLab software (Bio-Rad, Hercules, CA, USA).
Statistical analysis
All the experiments were performed in triplicates and one-way ANOVA (SPSS 17; IBM, Armonk, NY, USA) was used to compare the mean values of each treatment. The average of data from three independent experiments was taken and the significance in differences between the means of parameters were determined by using the Duncan test followed by the Dunnet test (P < 0.05) comparing between groups (control vs. treated).
Results
Heat shock elicits C. elegans immunity towards pathogenic exposure
To study the effect of external heat induction on the longevity of nematode, the lifespan of wild type C. elegans was checked by exposing the nematodes to a Gram-positive pathogen, S. aureus, and a Gram-negative opportunistic pathogen, P. mirabilis, with and without heat induction prior to exposure. The heat non-induced nematodes showed susceptibility to S. aureus with complete killing at 90 ± 2 h, whereas the nematodes exposed to external heat stress before S. aureus infection showed an extended lifespan up to 102 ± 2 h (Figure 1a). Induction of heat before pathogenic exposure improved the survival rate and lengthened the lifespan of nematodes. The observed result is in agreement with previous reports wherein S. aureus has been reported to kill wild type C. elegans in liquid cultures completely at 90 h.22,23 We also tested whether heat stress has an impact on the behavior of the opportunistic pathogen P. mirabilis and the laboratory food source E. coli OP50. The survival of wild type animals exposed to both strains was not significantly different between the heat shock-induced and non-induced groups. This result could be explained by the fact that P. mirabilis, being an opportunistic pathogen, is not fatal to the nematode and hence no difference in survival was observed with and without heat induction. To further expand this observation, chemotaxis assay was performed, which assesses the preference of C. elegans towards food over pathogen. Complementing the survival assay, the nematodes did not exhibit any aversion towards P. mirabilis. In fact, the nematodes were found to be more attracted to P. mirabilis than the food source (Figure 1b). However, both the heat-induced and non-induced nematodes moved towards the food source, avoiding S. aureus in the S. aureus-spotted plates (Figure 1b). However, no difference in the preference was observed between heat induced and non-induced worm. These observations clearly indicate that although the heat shock induction provided resistance to the nematode against virulent bacterial pathogen, S. aureus, it had no effect on the food preference or chemotactic behavior.
(a) Survival of wild type nematode pre-exposed to heat shock (HS) and subsequently infected with S. aureus and P. mirabilis. Nematodes fed on E. coli OP50 were considered as the control. (b) Food preference of the nematode after external heat induction. (c) Microscopic images of the nematodes with and without heat shock treatment before the pathogenic exposure.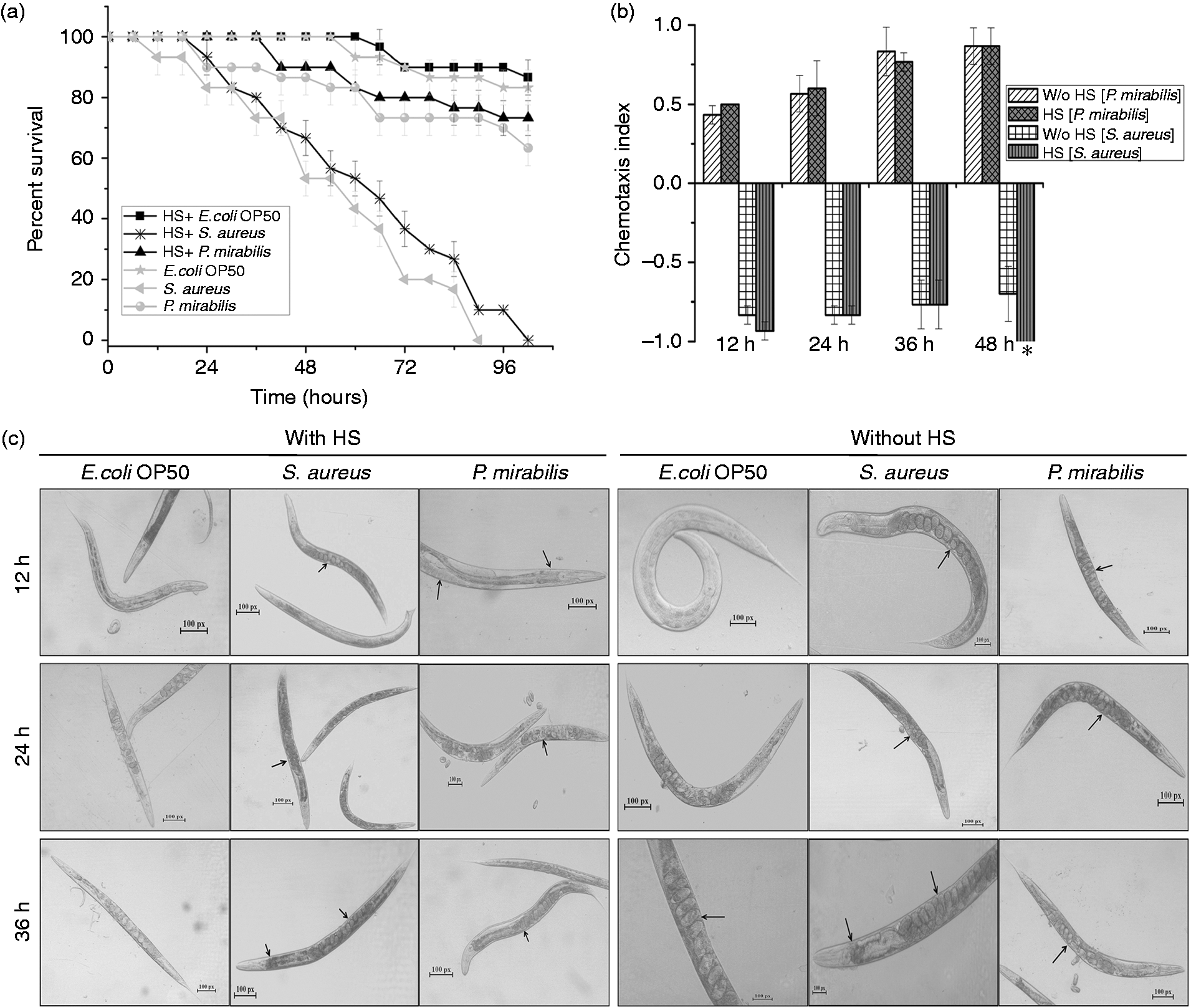
In order to get deeper insight into the physiological changes in nematodes during heat induction and consequent pathogenic exposure, microscopic visualization was performed, including controls. As expected, there was no significant change in the physiology of nematodes exposed to E. coli OP50 and P. mirabilis. However, prominent changes were observed in S. aureus-exposed nematodes. While the heat shock-induced nematodes showed a lesser amount of colonization, non-induced group had an elevated amount of colonization. Moreover, the nematodes that were not affected by heat stress showed pharyngeal disruption and immature eggs in the later hours (Figure 1c).
Higher transcript levels of daf-16 and sgk-1 increases the nematode lifespan
The expression level of genes (tol-1, daf-2, age-1, hsp-1, hsp-6, hsp-90, daf-16, sgk-1 bec-1) representing different immune and aging pathways present in C. elegans were monitored upon heat stress followed by pathogen exposure. tol-1 is Toll interleukin receptor-coding gene, which aids in activating the immune cascade via MAPK pathway. It plays a crucial role in mounting an appropriate immune response against broad range of pathogens and thereby confers resistance. tol-1 has been previously reported to help in innate immune regulation towards S. enterica and E. faecalis.24,25 The study by Jebamercy et al. also emphasized the need of tol-1 in the defense mechanism of the worm against Proteus species, and the involvement of MAPK pathway for immunity.
26
In the present study, the expression level of tol-1 during both pathogenic exposures was found to be consistently up-regulated in all the heat shock non-induced groups, whereas in heat shock-induced groups it was found to be down-regulated (Figure 2a, b).
Relative expression level of genes involved in the (a, b, e, f) MAPK pathway and the IIS pathway, (c, d) heat shock (HS) response and (g) autophagy of nematodes infected individually with P. mirabilis and S. aureus in the presence and absence of heat shock induction at various time periods (12, 24 and 36 h).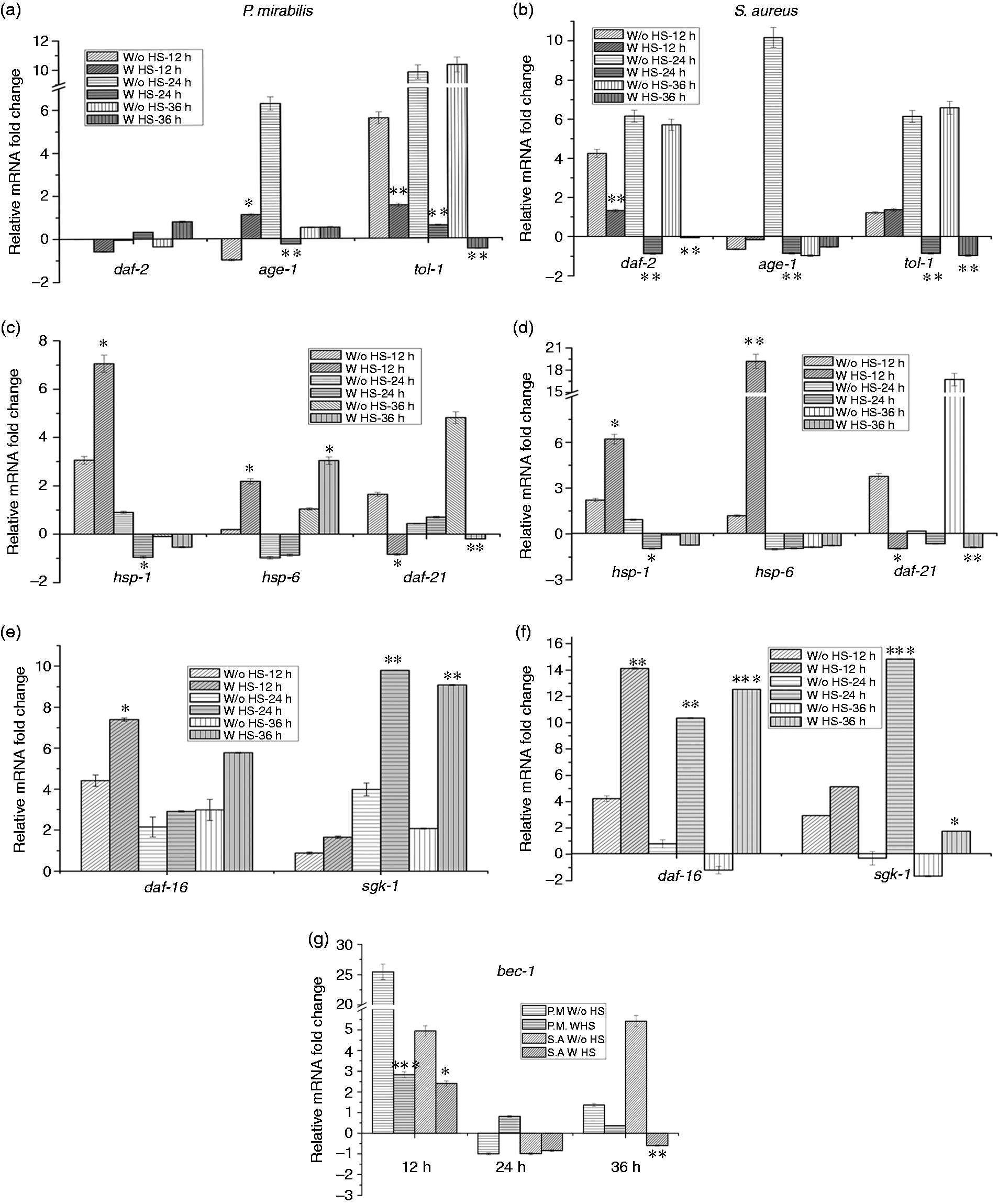
However, daf-2 and age-1 are intermediate players in the IIS pathway, which modulates longevity. The results of Kenyon et al. indicate that down-regulation of daf-2 prevents the activation of age-1, allowing daf-16 to translocate into the nucleus, which, in turn, becomes the reason for the increased lifespan of nematodes. 7 In the current study, no significant regulation of daf-2 was observed in both induced and non-induced nematodes during P. mirabilis exposure. However, a considerable down-regulation in the expression of daf-2 was observed in heat-induced samples during S. aureus exposure. In addition, except for 24-h exposed samples, no significant modulation in expression was observed for age-1 in both heat-induced and non-induced groups with P. mirabilis and S. aureus exposure (Figure 2a, b).
Induction of heat shock regulates a certain set of heat shock-responsive genes, such as hsp-1, hsp-6 and daf-21. 10 Notably, daf-21 is thought to play a crucial role in gonad function. Reduction of daf-21 levels in wild type nematodes leads to reduced motility and induction of the muscular stress response. 27 Hence, heat shock genes (hsp-1, hsp-6 and daf-21), were quantified for their expression levels. As expected, hsp-1 and hsp-6 were observed to be up-regulated in both S. aureus- and P. mirabilis- exposed nematodes in the initial hours of heat induction (12 h). However, no significant regulation was observed at later hours, probably owing to the diminished effect of heat induction. At 24 and 36 h, all the heat shock-responsive genes were down-regulated, except daf-21 in the heat shock non-induced samples (Figure 2c, d). The regulation of daf-16 and sgk-1 levels were also checked to provide more insight in their regulatory roles during the heat induction. daf-16 is a positive regulator of the IIS pathway and a crucial transcription factor. In the present study, daf-16 was observed to be up-regulated at 12 h when compared with its expression in the 24 and 36 h heat-induced samples and in the non-induced groups during P. mirabilis exposure (Figure 2e). In the S. aureus-exposed nematodes, the heat-induced samples had substantial up-regulated of daf-16 but was down-regulated in the non-induced samples as exposure time increased (Figure 2f). However, sgk-1, a serum glucocorticoid-inducible kinase involved in the IIS pathway showed a consistent up-regulation at all the time points of heat-induced samples subsequently exposed to P. mirabilis (Figure 2e). In the S. aureus-exposed samples there was a slightly diminished regulation at 36 h (Figure 2f) in the heat-induced groups.
BEC-1 is majorly involved in autophagy, a conserved lysosomal degradation pathway that provides primary defense against pathogenic exposure. It functions in programmed cell death by forming a complex with CED-9 and BCL-2. 28 As anticipated, in the present study, bec-1 was found to be highly modulated in the earlier stages of infection (12 h) in both P. mirabilis and S. aureus, when compared with later stages (24 h and 36 h). Of note, bec-1 was prominently up-regulated in heat non-induced P. mirabilis-exposed samples (Figure 2g).
IIS pathway is involved in extending the lifespan of daf-2 mutant nematodes
The general notion that IIS pathway activation decreases the longevity of nematodes was investigated. The mutation in daf-2 is universally known to increase the lifespan of C. elegans. Generally, daf-2 mutant nematodes have a resistance towards infection and other kinds of physiological stresses.
29
Strikingly, heat induction boosted the immune system of these nematodes during exposure to S. aureus, with a survival rate of 146 ± 2 h; unexposed nematodes had a survival rate of 136 ± 2 h (Figure 3). Heat shock increased the lifespan of daf-2 mutant nematodes during S. aureus infection by 18 h, which is twice that of wild type nematodes wherein only a 10 h increase in lifespan was observed. This observation prompted us to investigate the repercussions of heat shock on the IIS pathway.
Survival of daf-2 mutant induced with heat shock (HS) and subsequently infected individually with S. aureus (P < 0.05) and P. mirabilis (P < 0.05). E. coli OP50 food source served as control.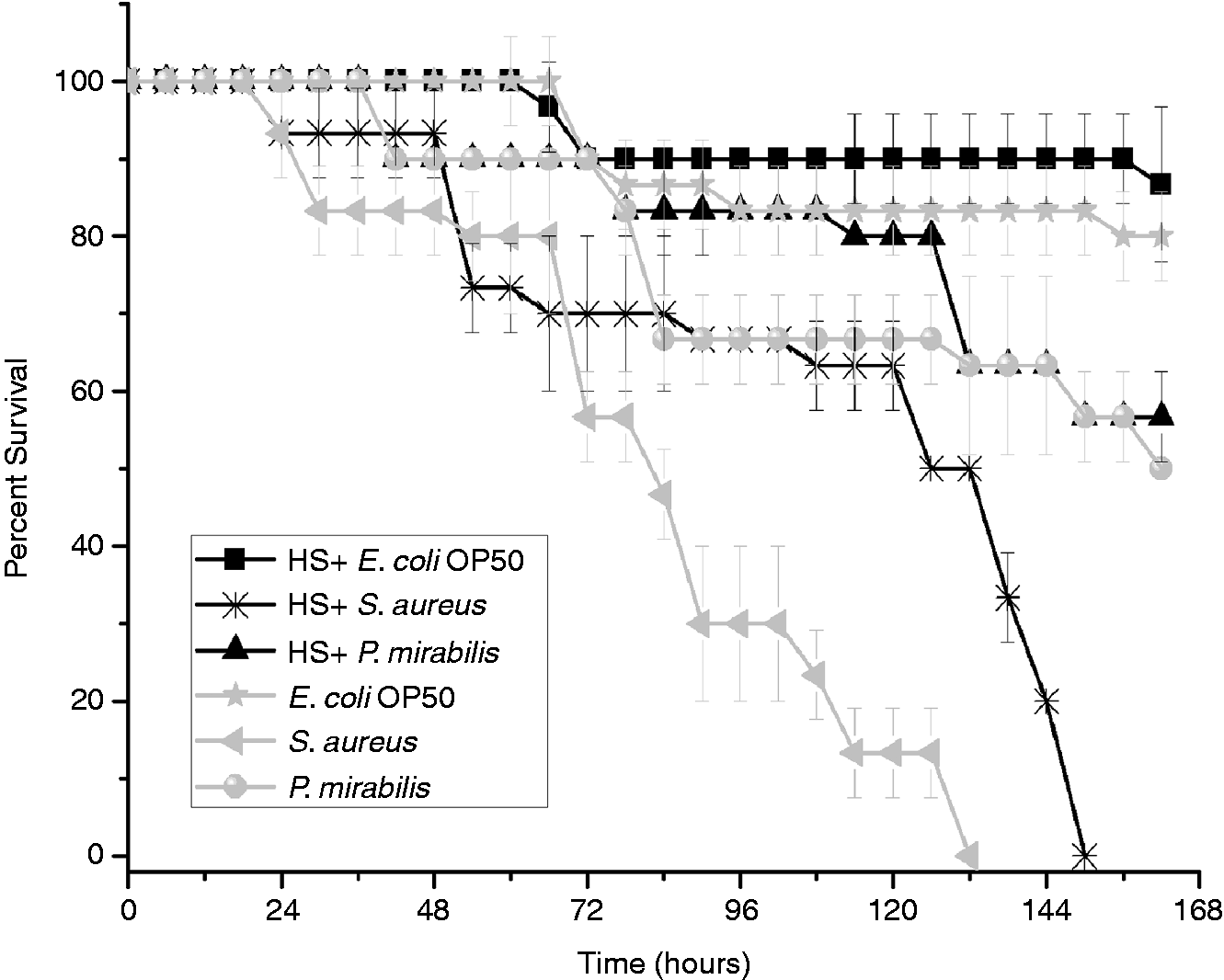
SGK-1 and HSF-1 aids in longevity of the nematode through parallel events
The protein expression levels of HSF-1, HSP-90 and SGK-1 were evaluated and quantified in the wild type and daf-2 mutant nematodes. HSPs are considered as the primary known target of the transcriptional factor HSF-1. Hence, both HSF-1 and HSP-90 were taken for protein-level quantification. It was also assumed that SGK-1 signals HSF-1 by acting parallel to it and supports the extension of lifespan. This hypothesis prompted us to check the presence of SGK-1 during heat induction before pathogenic exposure. The blot clearly depicts that heat shock treatment up-regulates the expression of SGK-1 in both N2 and daf-2 mutant during pathogenic exposure (Figure 4a), and the quantification graphs show more up-regulation of all three isoforms of SGK-1 in wild type than in daf-2 mutant nematodes during S. aureus infection, indicating that SGK-1 mediated activation of transcription factor DAF-16 (Figure 4b). The HSF-1 level is more up-regulated in wild type than in the daf-2 mutant, and identical regulation was detected in HSP-90 (Figure 4c, d). The elevated level of HSF-1 in the N2 also provides evidence on the concept that the protein chaperone HSP-90 is activated by the exertion of HSF-1 nuclear translocation. Moreover, the result shows that during heat shock, HSF-1 acts parallel to DAF-16 to provide pathogen resistance, thereby extending the lifespan.
(a) Western blot depicting the regulation of SGK-1, HSF-1 and HSP-90 proteins in wild type and daf-2 mutants infected with S. aureus in the presence and absence of heat shock pre-treatment. β-Actin was used as reference control. Relative intensity profile of (b) SGK-1(including its three isoforms), (c) HSF-1 and (d) HSP-90 with heat shock induction compared with control (without heat shock induction).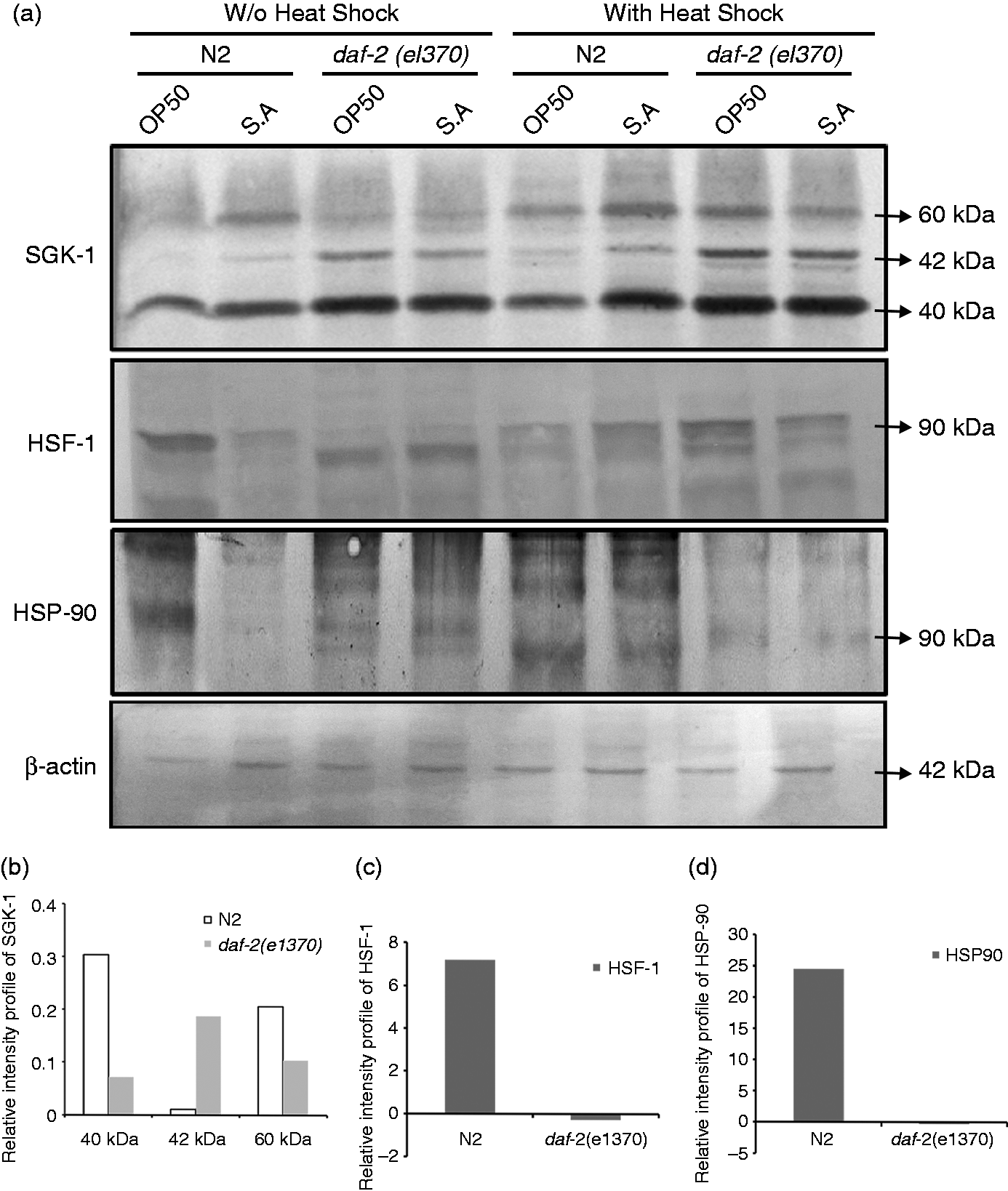
hsf-1 and hsp-90 mutations drastically reduce the lifespan of the nematode
The survival rate of heat-induced nematodes with mutation in hsf-1and hsp-90 upon pathogenic exposure was observed to be less than that of heat non-induced nematodes. Interestingly, they were found to have died earlier than heat non-induced wild type nematodes. The hsf-1 mutant nematodes induced by external heat stress and with sequential exposure to S. aureus had an early mortality with complete killing at 65 ± 2 h; the S. aureus-exposed group had complete killing at 78 ± 2 h (Figure 5a). The mortality rate was even higher in the hsp-90 mutants, PR673 (Figure 5b) and JT6130 (Figure 5c). Both had an LT50 at 30 ± 2 h. However, the complete killing of the nematodes induced by heat and subsequently by S. aureus was at 60 ± 2 h for the former and 65 ± 2 h for the latter. On the contrary, the nematodes exposed to P. mirabilis and E. coli OP50 had similar survival rate in both wild type and the mutant nematodes.
Survival of (a) hsf-1, (b) hsp-90, PR673, and (c) hsp-90, JT6130, mutants induced with heat shock and subsequently infected individually with S. aureus and P. mirabilis. Nematodes fed on E. coli OP50 were considered as the control. Significant difference was observed in (b) and (c) that were induced with heat shock and subsequently infected with S. aureus (P < 0.05).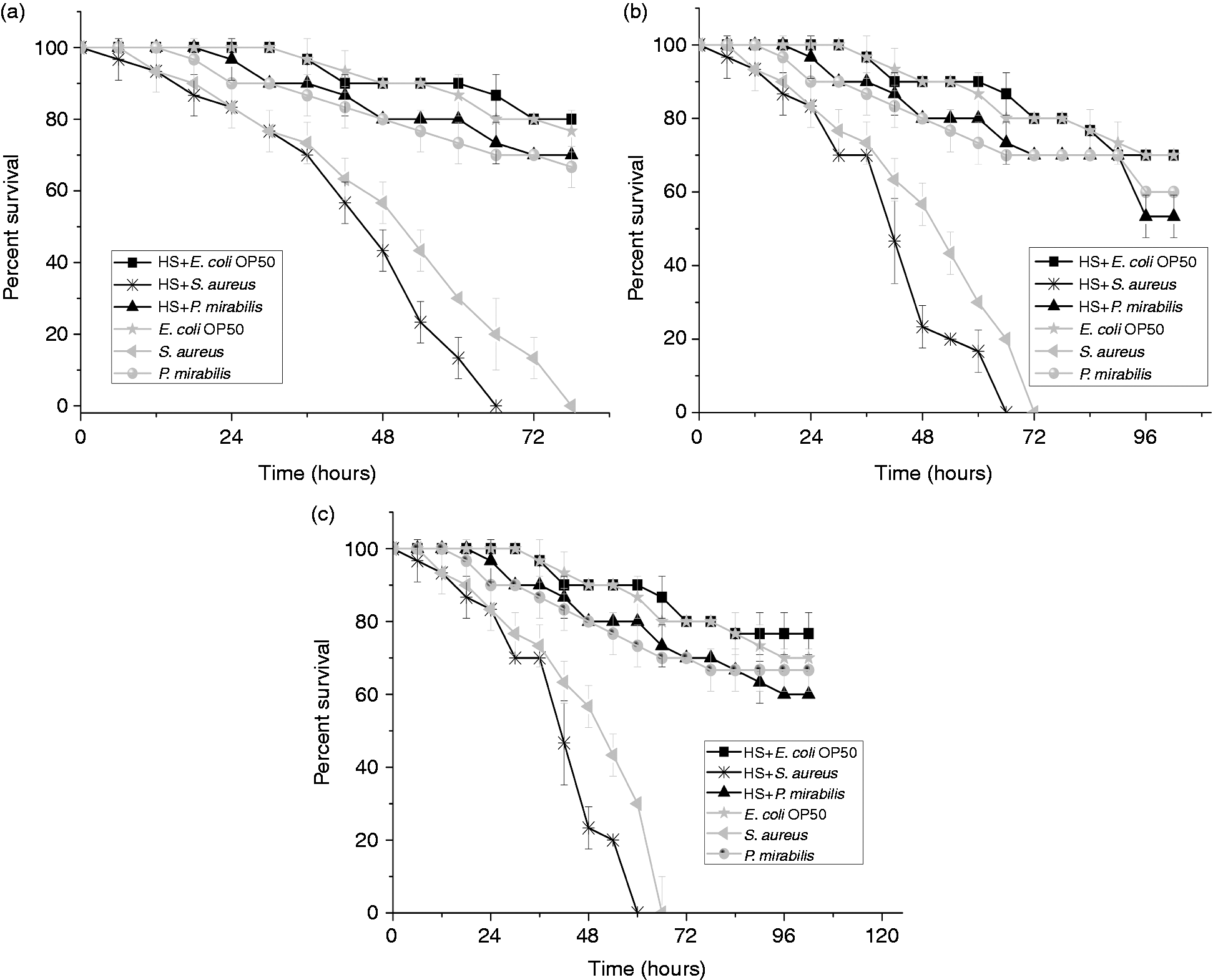
Discussion
C. elegans are typically grown at 15–25℃, and the temperature range of 30–35℃ is established as a stressor. 18 The nematodes respond to heat stress by up-regulating the expression of HSPs, a group of proteins involved in damage repair and also in the stabilization of immune homeostasis. HSF-1 is the prime receiver of heat shock, which, when accelerated, induces specific chaperones (HSPs) that facilitate maintenance or activation of a subset of client proteins important for stress resistance. 10 Reports from various independent studies prove a connection between heat shock response and longevity. For instance, a single heat shock applied at the starting stage of the C. elegans life cycle gave rise to a 20% longer lifespan, while multiple mild heat shocks throughout the lifetime increased its duration of survival by about 50%. 30 Heat stress was also reported to provide positive effects to C. elegans induced at 35℃ for 2 h. 18 However, prolonged exposure to this temperature killed the nematodes. 31 Moreover, studies have proved that the enhanced lifespan upon short-term induction of high temperature on L4-stage nematodes was not due to reverting back to the diapause state but to certain molecular modulations that aided in reduced death of the nematode. 18 However, the IIS pathway is widely known to be involved in aging, 32 and few studies have reported its role in pathogenic resistance. 29 HSPs, being the biomarkers of damage, are thought to work downstream of IIS pathway, but the link between activation of HSPs and the IIS pathway is still ambiguous. In the present study, the missing link between the IIS pathway and heat shock response was examined. In addition, the hypothesis that external heat shock might mediate pathogenic resistance was investigated.
The longevity of the host upon pathogenic exposure is determined by two factors, the first being how well the host is able to resist the pathogen by providing immunity, and the second being how well the normal molecular mechanism that provides the host with a long lifespan works. Hence, the nematodes were given a heat shock and subsequently exposed individually to S. aureus (a pathogen proven to be lethal to C. elegans) and P. mirabilis (an opportunistic pathogen proven to be non-lethal to C. elegans) and it was evaluated if heat shock could elicit resistance and increase longevity. Wild type nematodes induced with a heat shock were found to have a significant degree of resistance towards the pathogen, S. aureus. However, no such increase in lifespan was observed for P. mirabilis and the laboratory food source E. coli OP50 (Figure 1a). Similar results were also observed in a survival assay carried out using daf-2 mutant nematodes (Figure 3). daf-2 mutants generally enter the dauer stage and presumably lives for > 2 mo. Dauer larvae usually have a lower metabolic rate, are resistant to oxidative stress, and express high levels of antioxidant enzymes and HSPs. The increase in the survival rate of both the wild type and daf-2 mutant nematodes was directly related to the effect of heat shock applied. S. aureus, being a pathogen to nematodes, will elicit the IIS pathway, while exposure to the non-pathogens P. mirabilis and E. coli OP50 will not significantly elicit the same pathway. On the contrary, as heat shock was given during all the exposures, the heat shock pathway would have been activated all of them. Hence, the enhanced lifespan observed in S. aureus-exposed nematodes was possibly because of the combined effect of the activated IIS pathway and heat shock pathway. This preliminary result, which speculates on a positive link between the IIS pathway and the heat shock pathway, prompted us to explore the molecular mechanism involved.
In order to further validate the above results, real-time quantification of representative genes involved in both the pathways was performed. The MAPK pathway is the first innate immune response that is activated when the nematode is exposed to pathogens. In order to determine if the heat shock-mediated pathogenic resistance is via the MAPK pathway, expression of tol-1, the prime receptor of the pathway, was analyzed upon heat shock induction and subsequent pathogenic exposure. The up-regulation of tol-1 in the heat non-induced samples suggests activation of the MAPK pathway when there is a pathogenic encounter. In case of the P. mirabilis-exposed sample, the up-regulation was significant at the initial stage of infection, indicating that the nematode raised a response earlier, consequently suppressing the pathogenesis. However, in the S. aureus-exposed sample, the up-regulation was prominent only at the later stage, indicating that the host was not able to elicit a response against it, proving its pathogenicity towards the nematode. Conversely, tol-1 was found to be down-regulated in all the heat-induced samples (Figure 2a, b), providing evidence that heat shock-mediated pathogenic resistance is not associated with the MAPK pathway.
Furthermore, the expression of daf-2 and age-1, the receptor and the adapter proteins, respectively, of the IIS pathway was analyzed to examine its role in pathogenic resistance and longevity. The transcript level of daf-2 was down-regulated at all three time points in S. aureus exposure alone in heat-induced samples (Figure 2a, b). Earlier studies have provided evidence that daf-2 down-regulation increases longevity of nematodes, as its suppressive effect on daf-16 was relieved, allowing the translocation of daf-16 into the nucleus. 33 The present study provides similar evidence for the observed longevity, as daf-2 was down-regulated in S. aureus-exposed heat-induced samples alone in which, enhanced life span was observed. Complementing this result, expression of age-1, the downstream adapter of daf-2 was found to be down-regulated in heat-induced, S. aureus-exposed samples (Figure 2b). The expression levels of tol-1, daf-2 and age-1 together validate our earlier hypothesis that heat shock mediates pathogenic resistance by associating with the IIS pathway.
To validate that external heat shock does, indeed, activate the heat shock response pathway, irrespective of the pathogenic exposure, candidate heat shock genes were selected and their expression analyzed. The initial hours (12 h) of heat shock-induced samples in the case of both S. aureus and P. mirabilis showed elevated levels of hsp-1 and hsp-6, implying the necessity of the chaperonic molecule in ameliorating the damage caused by heat shock (Figure 2c, d). The report by Gaiser et al. states that the expression level of daf-21 was scarcely affcted by heat stress compared with any other heat shock protein. 27 The results of present study complement those of Gaiser et al. (Figure 2c, d), 27 indicating that the declined expression level of daf-21 does not affect the longevity of nematodes; rather, the collective effect of HSPs provided a proper immune response against the pathogen. Analogous to the regulation pattern of daf-16, sgk-1 gene expression in heat-induced samples and consecutive pathogenic exposures have suggested that sgk-1 was required for the regulation of the daf-16 transcription factor (Figure 2e, f). bec-1 in C. elegans aids in autophagy. 33 Autophagy was recently linked directly to aging in C. elegans, because autophagy was shown to play a critical role in several nutrient-sensing longevity processes, including the TOR and IIS signaling pathways, 34 as well as in the dietary restriction paradigm. 35 In the absence of heat induction, bec-1 was found to be considerably up-regulated in the P. mirabilis-exposed sample than the S. aureus-exposed sample (Figure 2g). A conceivable speculation could be that the significant up-regulation of bec-1 efficiently induced autophagy and protected the nematodes from P. mirabilis infection. However, the induction level observed in S. aureus was not enough to confer such resistance. Comparing the expression level between heat-induced and non-induced samples, bec-1 was found to be majorly down-regulated in heat shock-induced samples, manifesting our earlier hypothesis that heat shock induces pathogenic resistance by means other than the native innate immune pathways. Determination of the transcriptomic status of bec-1 in the present study revealed that decreased activity of bec-1 in the late hours stimulated the cell death machinery. Therefore, the loss of bec-1 leads to decline in the lifespan.
To further scrutinize the possible relationship between heat stress and the innate immune response of the nematode, the study was extended further to validate, via immuno-blotting, the involvement of the IIS and heat shock pathways. Representative players of both the pathways (SGK-1, DAF-21 and HSF-1) were selected for analysis. SGK-1 was selected as a representative of the IIS pathway, as it has been reported to activate directly DAF-16. 36 However, HSF-1 was selected with the rationale that it is the core regulator that controls all other heat shock proteins. 31 DAF-21 was chosen for analysis to validate the results of real-time PCR, which suggested its insignificant role in a heat shock-mediated response.
Complementing the real-time results, HSF-1 and SGK-1 were found to be up-regulated in heat shock-induced samples of both the wild type and daf-2 mutants (Figure 4a–c). Contradicting the real-time quantification results, HSP-90 was also observed to be up-regulated at the protein level (Figure 4a, d). A plausible reason could be that HSP-90 is under the control of some post-transcriptional regulation. This hypothesis needs to be validated further. A recent study by Mizunuma et al. reported that SGK-1, a component of DAF-2 signaling pathway, modulates longevity, depending on environmental cues.
37
Yet another study, by Belova et al., reported that HSP-90 is required for the phosphorylation and activation of SGK-1.
38
McColl et al. reported that the overall mRNA levels of several HSPs are significantly higher in IIS mutants.
39
It is also known that the expression of certain HSF-1 and HSP targets are at least partially accountable for the lifespan extension observed in IIS pathway mutants.
6
In addition, a recent report by Durai et al. has also supported the view that the high transcript and protein levels of HSP are needed for both the immune and stress responses.
40
The observation that SGK-1 is enhanced upon heat shock induction and subsequent pathogenic exposure, even in daf-2 mutants, in the present study, suggest that SGK-1 up-regulation was not mediated by daf-2. It is possible that HSP-90, which is up-regulated upon heat shock, would have activated SGK-1, which, in turn, would have further activated its downstream regulators, including DAF-16. This evidence further strengthens our earlier hypothesis that the IIS and heat shock pathways together play a role in the enhanced lifespan observed upon pathogen encounter. The proposed pathway for the enhanced lifespan observed upon pathogenic exposure in heat shock-induced nematodes mediated by the heat shock and IIS pathways is depicted in Figure 6.
Schematic representation of the plausible role of HSF-1 in inducing longevity upon heat induction and consequent pathogenic exposure.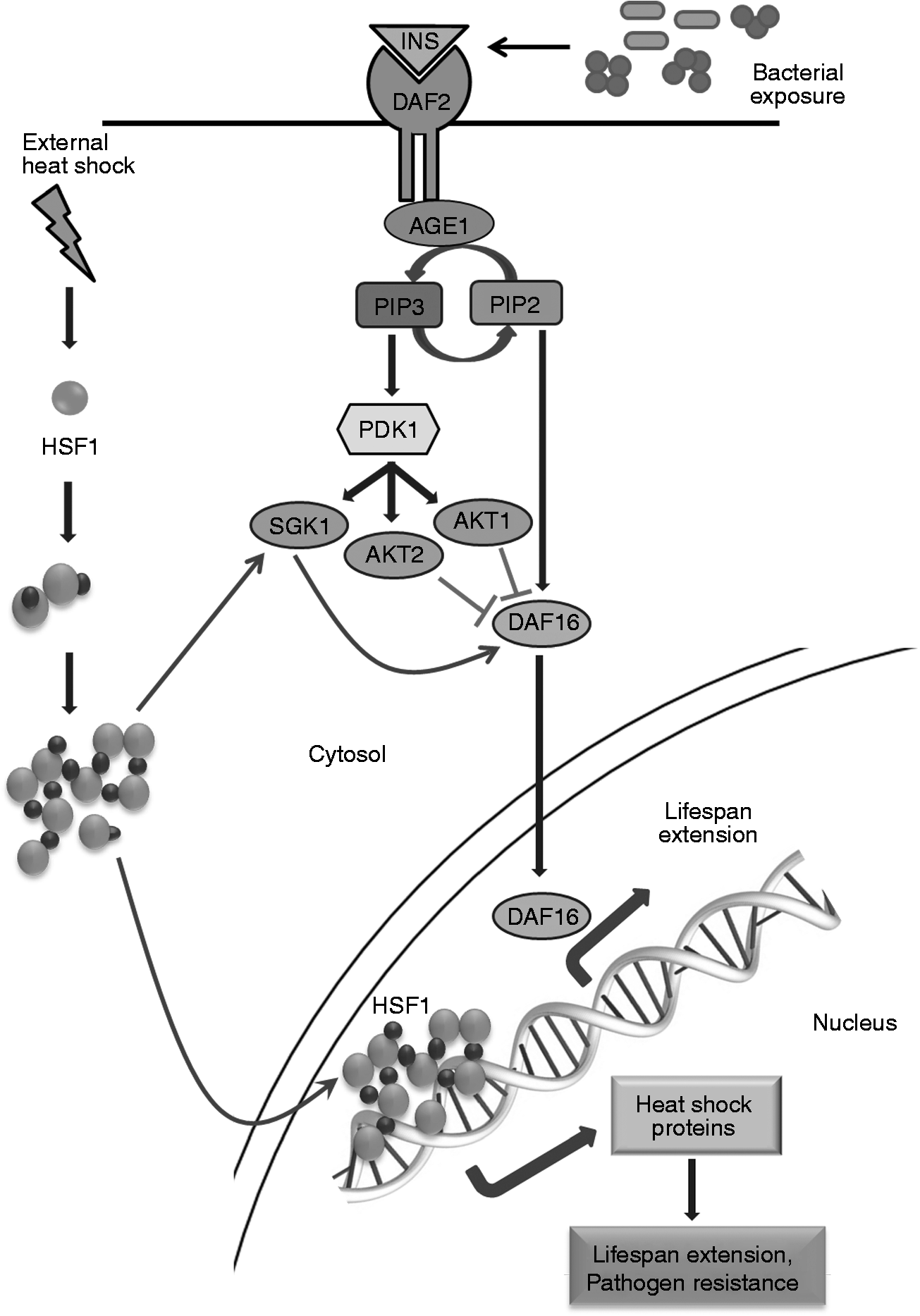
Finally, in order to further confirm the obtained results, which speculate a key role for the heat shock pathway in pathogenic resistance and longevity, a survival assay was performed using hsf-1 and hsp-90 mutants. The results convincingly propose that mutation in both hsf-1and hsp-90 shorten the lifespan of nematodes. This observation is consistent with a report by Morley and Morimoto, wherein hsf-1 mutants have a decreased lifespan. 8 Mutants exposed to both heat shock and pathogens had a decreased lifespan compared with the groups that were exposed individually to either heat shock or pathogens. This result confirms that, in addition to its role in longevity, the heat shock pathway is also involved in pathogenic resistance and immunity.
Conclusion
Analysis of key factors that link the IIS pathway with the heat shock prone longevity mechanism (expression levels of HSF-1, HSP-90 and SGK-1, and survival assay using mutants) has revealed a significant relationship between the heat shock response and pathogenic resistance. The results of the present study suggest that increasing the ambient growth temperature of the nematode enhances resistance to S. aureus pathogenesis. Notably, the results for the first time, SGK-1 is put forth as the key determinant linking the IIS pathway and the heat shock response pathway, which ultimately contributes to pathogenic resistance and increased life span of the nematodes. Further research is needed to identify other novel proteins and players involved in heat shock-mediated pathogenic resistance.
Footnotes
Acknowledgements
We are grateful to Caenorhabditis Genetics Center, which is funded by the National Institute of Health, National Centre for Research Resources for providing C. elegans N2 WT, mutant strains and E. coli OP50. We thank S. Marudhu Pandiyan and Prasanth Mani Iyer for their technical assistance, and D. Viszwapriya for critical reading of the manuscript. We acknowledge the Computational and Bioinformatics facility provided by Alagappa University Bioinformatics Infrastructure Facility (funded by DBT, GOI; Grant No. BT/BI/25/001/2006).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: KB gratefully acknowledges the Department of Biotechnology (DBT), University Grants Commission (UGC), Indian Council of Medical Research (ICMR), Council of Scientific and Industrial Research (CSIR) and Department of Science and Technology (DST), Government of India, New Delhi, India, for the financial assistance. UP is indebted to the Department of Science and Technology (DST No: SR/SO/AS-80/2010) for the financial assistance (DST SERB- JRF). Appreciation is also due to PURSE [Grant No.SR/S9Z-23/2010/42(G)] & FIST (Grant No.SR-FST/LSI-087/2008) and SAP-DRS-I [Grant No.F.3-28/2011 (SAP-II)].