
Abstract
Enterococcus faecalis is considered a major bacterial pathogen implicated in endodontic infections and contributes considerably to periapical periodontitis. This study aimed to investigate the potential mechanisms by which E. faecalis accounts for the bone destruction in periapical periodontitis in vitro. Osteoclast precursor RAW264.7 cells were treated with E. faecalis ATCC 29212 and a wild strain of E. faecalis derived clinically from an infected root canal. The results showed that, to some extent, E. faecalis induced the RAW264.7 cells to form tartrate-resistant acid phosphatase (TRAP)-positive multinucleated osteoclast-like cells. This pathogen markedly stimulated RAW264.7 cells to express semaphorin 4D (Sema4D), which inhibits bone formation. Once RAW264.7 cells were primed by low-dose receptor activator of nuclear factor-kappa B ligand (RANKL), E. faecalis could significantly increase the production of TRAP-positive multinucleated cells and up-regulate the expression of osteoclast-specific markers, including NFATc1, TRAP and cathepsin K. Both p38 and ERK1/2 MAPK signaling pathways were activated by E. faecalis in RANKL-primed RAW264.7 cells, and meanwhile the expression of Sema4D was highly increased. In conclusion, E. faecalis may greatly contribute to the bone resorption in periapical periodontitis by promoting RANKL-dependent osteoclastogenesis and expression of Sema4D through activation of p38 and ERK1/2 MAPK signaling pathways.
Keywords
Introduction
Enterococcus faecalis is a facultative anaerobic Gram-positive bacterial pathogen strongly associated with primary endodontic infections, 1 and is mainly found in root-filled teeth with refractory periapical periodontitis. 2 Persistent bacterial infection of the root canal system leads to periapical inflammation and subsequent periapical bone destruction. 3 Therefore, further understanding of the mechanism underlying E. faecalis-induced bone resorption may accelerate the development of novel therapeutic strategies to modulate effectively the destructive process of these periapical inflammatory bone lesions.
Bone remodeling is dependent on osteoclast–osteoblast crosstalk and interactions. The dynamic process of bone remodeling can be divided into initial stage of bone resorption, transition of differentiation of osteoblasts and new bone formation until quiescence. In the initial stage, the migration of osteoclast precursors and the differentiation of osteoclasts play a pivotal role in bone resorption. 4 Another critical determinant to the outcome of this process is the balance of both osteoblast and osteoclast functional activities. Disruption of this intricate balance of bone remodeling, for example higher bone resorption by osteoclasts or weaker bone formation by osteoblasts, can be seen in various bony lesions or disorders such as osteopenia and osteoporosis.5,6 Osteoclasts that degrade bone matrix are multinucleated cells originating from the fusion of monocyte/macrophage precursor cells. 7 Receptor activator of nuclear factor-kappa B ligand (RANKL) is essential for osteoclast differentiation and can be expressed in osteoblasts, stromal cells and lymphocytes. 8 RANKL and M-CSF are essential for osteoclast differentiation, 9 whereas it has been reported that fusion and activation of osteoclasts can rely only on RANKL independent of M-CSF. 10 Inflammatory cytokines play important roles in the regulation of osteoclastogenesis and bone resorption. It has also been shown that the osteoclast-like cell differentiation can be induced by TNF-α in synergy with or independent of RANKL.11,12 Although it is controversial whether LPS alone can induce RAW264.7 cells to form osteoclast-like cells, LPS stimulation promotes the late phase of osteoclast differentiation induced by RANKL.13,14
There are key markers for the different stages of bone remodeling. Tartrate-resistant acid phosphatase (TRAP) is a marker highly expressed by osteoclasts during bone resorption. 15 Serine protease cathepsin K is another marker involved in bone resorption, which is produced while osteoclasts dissolve bone matrix elements. 16 The transcription factor, nuclear factor of activated T cells c1 (NFATc1), modulates osteoclast-specific genes, including those of TRAP and cathepsin K.17,18 As a local factor, semaphorin 4D (Sema4D), expressed by osteoclasts, mediates osteoclast–osteoblast communication.19,20 There are two types of Sema4D: transmembrane and soluble proteins. 21 Sema4D may inhibit bone formation in the initial stage of bone remodeling and increase the osteoblastic motility to maintain a certain distance between osteoblasts and osteoclasts. 20 Sema4D also modulates the bone resorption indirectly, dependent on the ovarian function by an unknown pathway. 22
To date, little is known about the effects of E. faecalis on osteoclasts with regard to the destruction of bone homeostasis often seen in refractory periapical periodontitis. In the present study, we hypothesized that E. faecalis could induce bone resorption through promotion of osteoclastogenesis. Here, we used heat-killed E. facaelis ATCC 29212 and clinical strain P25RC to treat murine RAW264.7 cells and the RANKL-primed RAW264.7 cells, respectively, to investigate the effects of E. faecalis and the possible underlying mechanism of bone resorption related to persistent periapical periodontitis.
Materials and methods
Preparation of heat-killed bacteria
Enterococcus facaelis ATCC29212 was purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA). Another clinical strain, E. faecalis P25RC, was isolated from an infected root canal at the Hospital of Stomatology of Peking University. 23 The two strains of E. faecalis were cultured in brain–heart infusion (BHI) broth anaerobically at 37℃. Standard bacterial inoculum at an OD of 0.5 at 600 nm, which corresponded to 2 × 108 CFU/ml, was prepared from mid-logarithmic phase cultures. Contaminations were ruled out by Gram staining. The bacteria were collected by centrifugation, washed twice with PBS (pH 7.4) and re-suspended in a small volume of PBS. The bacteria were killed by heating to 85℃ for 1 h. The inactivated bacteria were then inoculated on BHI agar plates and cultured overnight anaerobically and aerobically at 37℃, which showed that there were no colonies, confirming the heat-killing process.
Cell culture and osteoclastogenesis
The murine macrophage cell line, RAW 264.7 cells (TIB 71; ATCC) were cultured in DMEM (Gibco, Life Technologies, Carlsbad, CA, USA) supplemented with 10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin under a humidified 5% CO2 atmosphere at 37℃. RAW264.7 cells were seeded in six-well plates at a density of 2 × 105 cells/well and treated with the two heat-killed strains of E. faecalis for 6 d, or RAW264.7 cells were primed with 20 ng/ml recombinant murine RANKL (R&D Systems, Minneapolis, MN, USA) for 4 d and then treated with E. faecalis for 2 d. The culture medium was replaced every 2 d.
Cell proliferation assay
RAW264.7 cells were seeded in a 96-well plate at a density of 2 × 103 cells/well. After 24 h of culture, the cells were treated with different multiplicities of infection (MOIs) of the two heat-killed strains of E. faecalis for an additional 2, 4 and 6 d. Cell proliferation was assessed using a Cell Counting Kit-8 (Sigma-Aldrich, St. Louis, MO, USA), following the manufacturer’s instructions. Each treatment was performed six times. Absorbance was measured at 450 nm with spectrophotometry.
Gene expression analysis
Total RNA was extracted using a RNeasy Plus Mini Kit (Qiagen, Hilden, Germany), and the concentration of RNA was measured using a Nanodrop 2000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA). Reverse transcription was performed using SuperScript VILO MasterMix (Invitrogen, Carlsbad, CA, USA) and cDNA was used as a template. Primer sequences were as follows: cyclooxygenase 2 (COX-2), 5’(TGAGTACCGCAAACGCTTCTC)3’ and 5’(TGGACGAGGTTTTTCCACCAG)3’; 24 c-fos, 5’(ACGTGGAGCTGAAGGCAGAAC)3’ and 5’(AGCCACTGGGCCTAGATGATG)3’; 25 NFATc1, 5’(CAAGTCTCACCACAGGGCTCACTA) 3’ and 5’(TCAGCCGTCCCAATGAACAG)3’; 25 TRAP, 5’(ACACAGTGATGCTGTGTGGCAAC TC)3’ and 5’(CCAGAGGCTTCCACATATATGATGG)3’; 26 cathepsin K, 5’(CTGAAGATGC TTTCCCATATGTGGG)3’ and 5’(GCAGGCGTTGTTCTTATTCCGAGC)3’; 26 GAPDH 5’(ATGTGTCCGTCGTGGATCTGA)3’ and 5’(ATGCCTGCTTCACCACCTTCT)3’. 27 Real-time PCR was carried out using a Real-Time PCR StepOne system (Applied Biosystems, Foster City, CA, USA) and the PCR products were detected with SYBR Green (Life Technologies, Austin, TX, USA). The reaction conditions were 95℃ for 10 min, 95℃ for 15 s for 40 cycles and 60℃ for 1 min. The experiment was performed in triplicate. The relative expressions for each gene were normalized to the expression of GAPDH using the 2−ΔΔCт method. 28
ELISA assay
The culture supernatants were collected. The expressions of TNF-α, IL-1β and IL-6 were detected using corresponding ELISA kits (R&D Systems). Soluble Sema4D was detected using a mouse Sema4D kit (Sino Biological Inc., Beijing, China). The detection was repeated three times and carried out in triplicate.
Blocking assay
A blocking assay was performed to explore the role of TLR2 in the E. faecalis-mediated effects on RAW264.7 cells. RAW264.7 cells were seeded in a 96-well plate at a density of 2 × 104 cells/well. Cells were pretreated with 10 µg/ml anti-TLR2 mAb (InvivoGen, San Diego, CA, USA) for 1 h prior to exposure to the two heat-killed E. faecalis strains at an MOI of 10 for 6 h and 24 h, respectively. The level of TNF-α in culture supernatant was detected using ELISA.
TRAP assay
Cells were washed with PBS and fixed with 4% paraformaldehyde for 15 min. TRAP staining was carried out using the Acid Phosphatase Leukocyte kit (Sigma-Aldrich). Each treatment was performed in triplicate. The TRAP-positive multinucleated cells were observed and counted under a microscope.
Western blotting
The whole-cell lysate was obtained using M-PER Mammalian Protein Extraction Reagent (Thermo Scientific, Rockford, IL, USA) and quantified with a BCA protein assay kit (Thermo Scientific). Briefly, 30 µg of protein was subject to 12% SDS-PAGE (Bio-Rad Laboratories) and blotted onto polyvinylidene fluoride membranes (GE Healthcare, Uppsala, Sweden). After blocking for 1 h in TBS-T (20 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20) containing 5% skim milk, the membranes were washed three times in TBS-T and incubated with specific anti-phospho- and anti-MAP kinase Abs for ERK1/2 and p38 (Cell Signaling, Beverly, MA, USA) and anti-Sema4D Ab (Abcam, Hong Kong) overnight at 4℃. Following three washes in TBS-T, the membranes were probed with second HRP-conjugated anti-rabbit Ab for 2 h. The membranes were developed using enhanced chemiluminescence reagents (Thermo Scientific). The proteins were normalized to β-actin (Santa Cruz, Santa Cruz, CA, USA). The density of bands was analyzed by Quantity One 4.6.9 software (Bio-Rad Laboratories).
Statistical analysis
Data are presented as means ± SD and analyzed by ANOVA using SPSS version 20 (IBM, Armonk, NY, USA). The Bonferroni method was used to compare the difference between two groups. A P-value < 0.05 was considered significantly different.
Results
Effects of E. faecalis on RAW264.7 cells
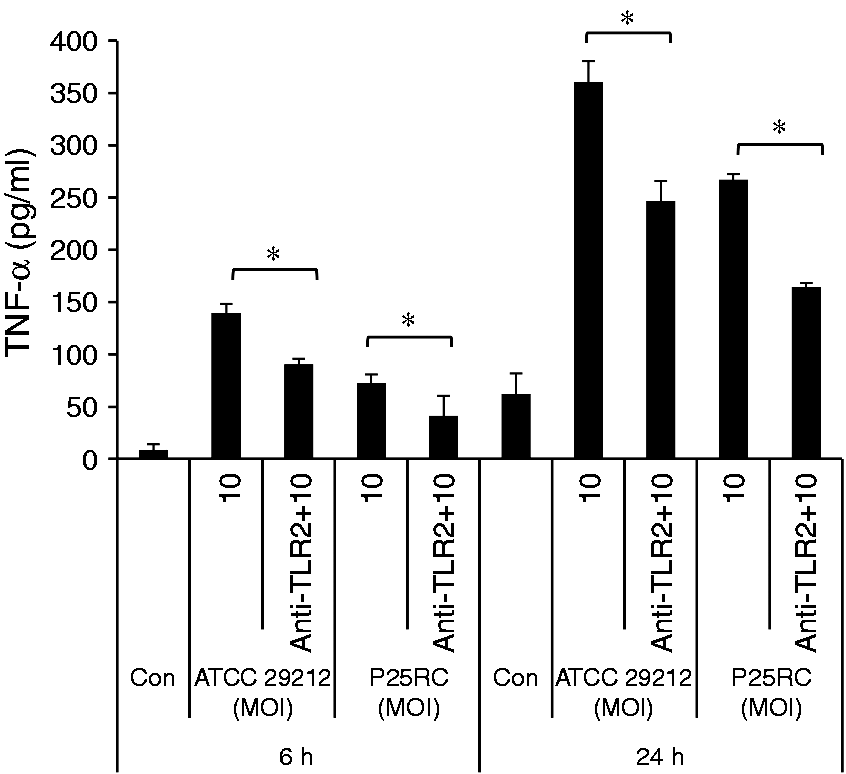
Enterococcus faecalis inhibited the proliferation of RAW264.7 cells when incubated at different MOIs for 2, 4 and 6 d (Figure 1A). The expression of TNF-α was significantly induced by both standard and clinical E. faecalis strains at an MOI of 1000 in RAW264.7 cells (Figure 1B). However, the expressions of IL-1β and IL-6 were not promoted (Figure 1C, D). The gene expression of COX-2 was highly induced (Figure 1E). Western blot analysis revealed the basal Sema4D expression in the osteoclast precursor RAW264.7 cells. Sema4D expression was significantly increased in the presence of E. faecalis at an MOI of 1000 compared with the control without E. faecalis stimulation (Figure 1F). This result was further confirmed by ELISA, which showed increased soluble Sema4D in RAW264.7 cells when treated with E. faecalis (P < 0.05) (Figure 1G). While TLR2 on RAW264.7 cells was blocked by anti-TLR2 mAb, the expression of TNF-α was significantly decreased (Figure 2).
The effects of E. faecalis on RAW264.7 cells. (a) The proliferation of RAW264.7 cells with treatment of the two strains of E. faecalis at different MOIs for 2, 4 and 6 d, respectively. RAW264.7 cells were treated with the two strains of E. faecalis at an MOI of 1000 for 6 d and the last 2-d culture supernatants were collected. The expressions of (b) TNF-α, (c) IL-1β and (d) IL-6 were assayed using ELISA kits. RAW264.7 cells were treated with the two strains of E. faecalis at an MOI of 1000 for 2 d. (e) The gene expression of COX-2 was assayed using real-time PCR. (f) The expression of Sema4D was assayed using Western blotting. (g) The soluble Sema4D was detected using the ELISA assay. The mean and SD are shown. *P < 0.05 was considered statistically significant. Untreated RAW264.7 cells were set to be the control. Anti-TLR2 mAb significantly inhibited the effect of E. faecalis on TNF-α expression. RAW264.7 cells were pretreated with 10 µg/ml anti-TLR2 mAb for 1 h prior to exposure to E. faecalis at MOI of 10 for 6 h and 24 h. The mean and SD are shown. *P < 0.05 was considered statistically significant. Con: untreated RAW264.7 cells (control).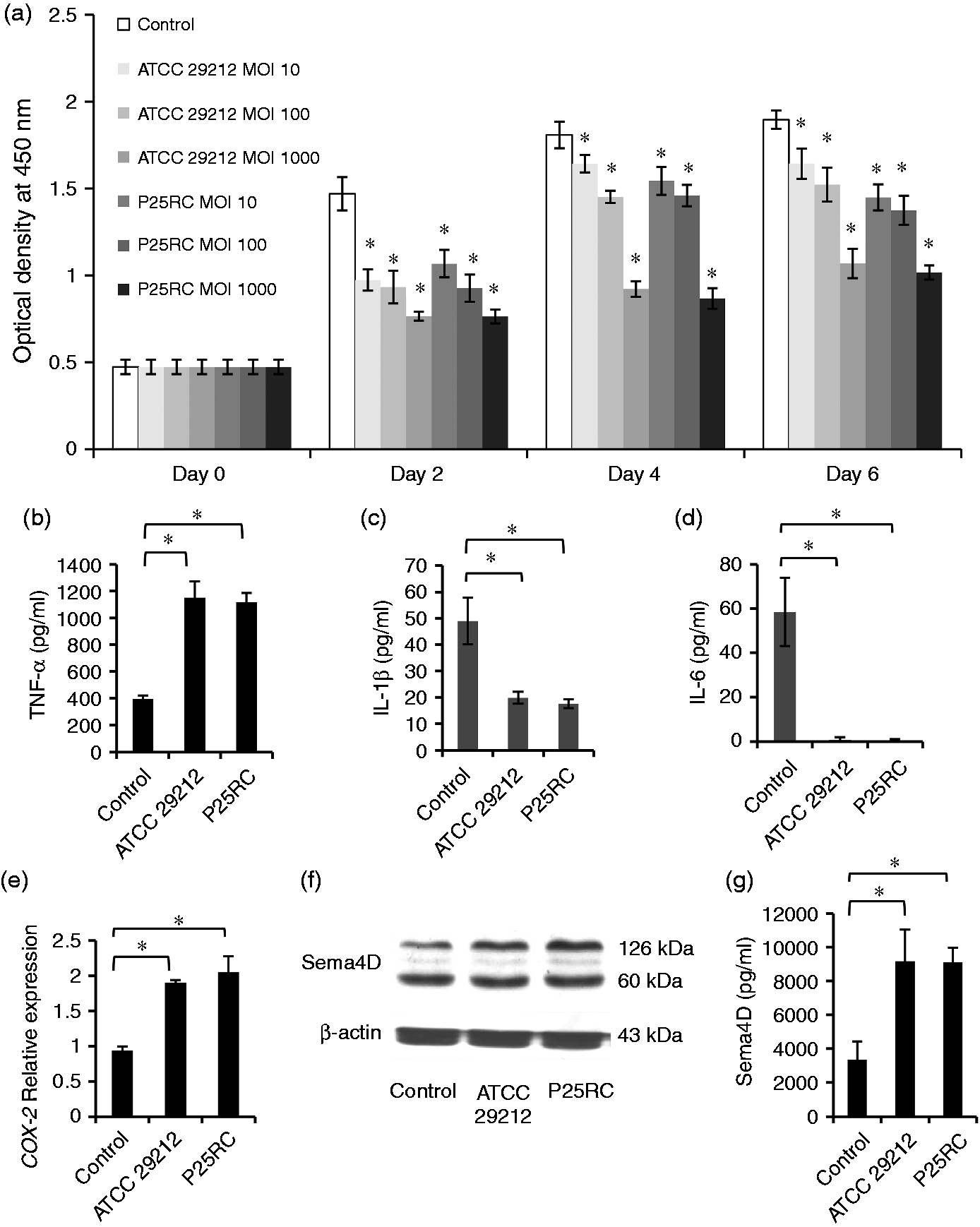
Effects of E. faecalis on osteoclastogenesis
RAW264.7 cells were treated with the two E. faecalis strains at an MOI of 1000 for 6 d and untreated cells served as the control. The TRAP staining assay demonstrated that E. faecalis alone could slightly induce the formation of TRAP-positive multinucleated cells (Figure 3A). Then, RAW264.7 cells were pretreated with low-dose RANKL for 4 d followed by the challenge of two E. faecalis strains at different MOIs for 2 d. RAW264.7 cells in the absence of E. faecalis were cultured for 6 d as a control. Interestingly, a significantly higher amount of TRAP-positive multinucleated cells were generated in RANKL-primed cells with the treatment of E. faecalis compared with control cells (P < 0.05) (Figure 3B).
Osteoclast differentiation induced by E. faecalis in RAW264.7 cells and RANKL-primed cells was detected using a TRAP staining assay. (a) RAW264.7 cells were treated with the two strains of E. faecalis at an MOI of 1000 for 6 d. (b) RAW264.7 cells were primed with low-dose RANKL (20 ng/ml) for 4 d and then treated with the two strains of E. faecalis at different MOIs for 2 d. A TRAP staining assay was carried out and visualized at 200× magnification. Scale bar: 100 µm. The numbers of TRAP-positive multinucleated cells (MNCs) with >3 nuclei were counted from nine random fields of view under an inverted bright field microscope. The mean and SD are shown. *P < 0.05 was considered statistically significant. Untreated RAW264.7 cells were set to be control.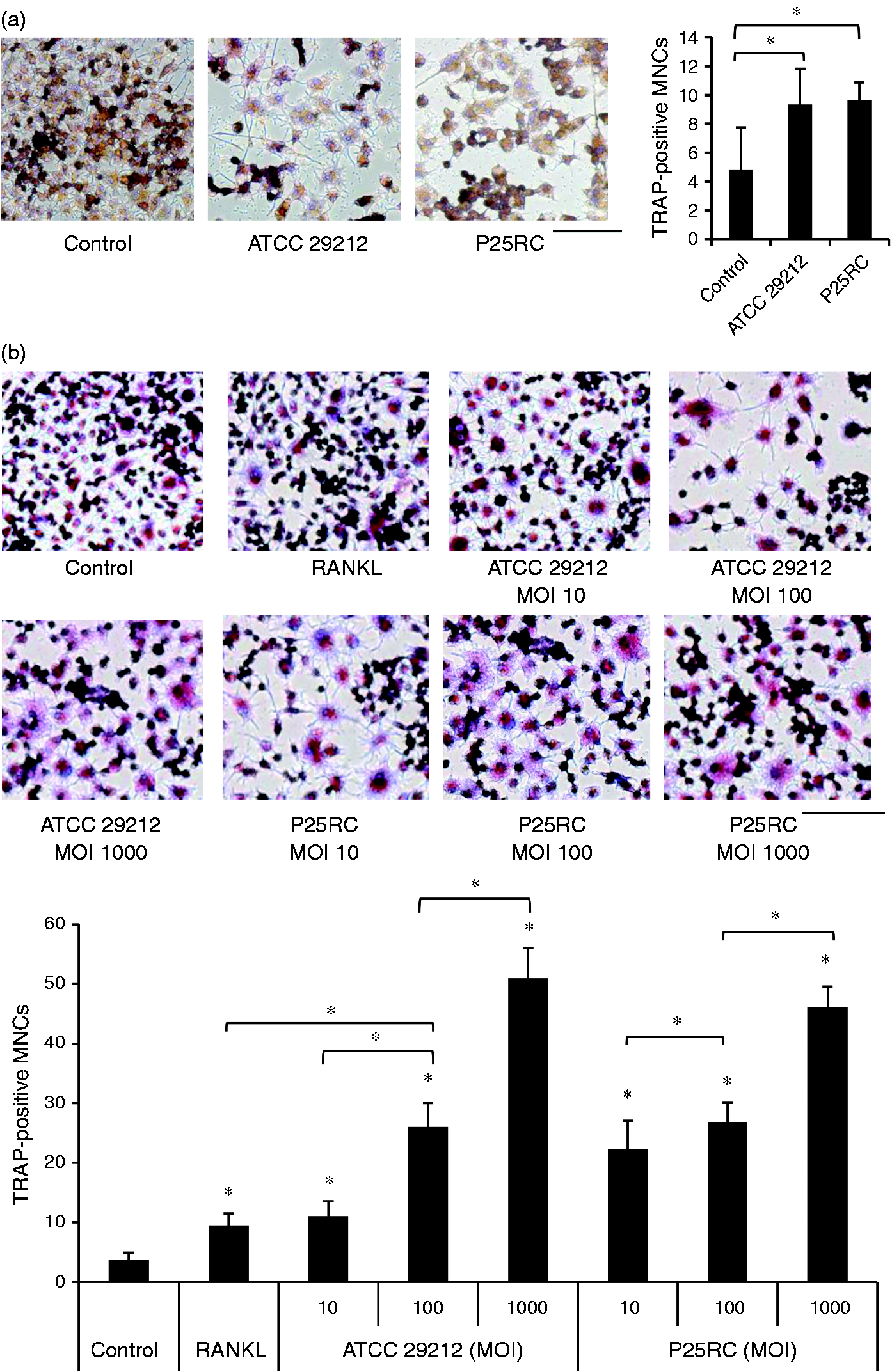
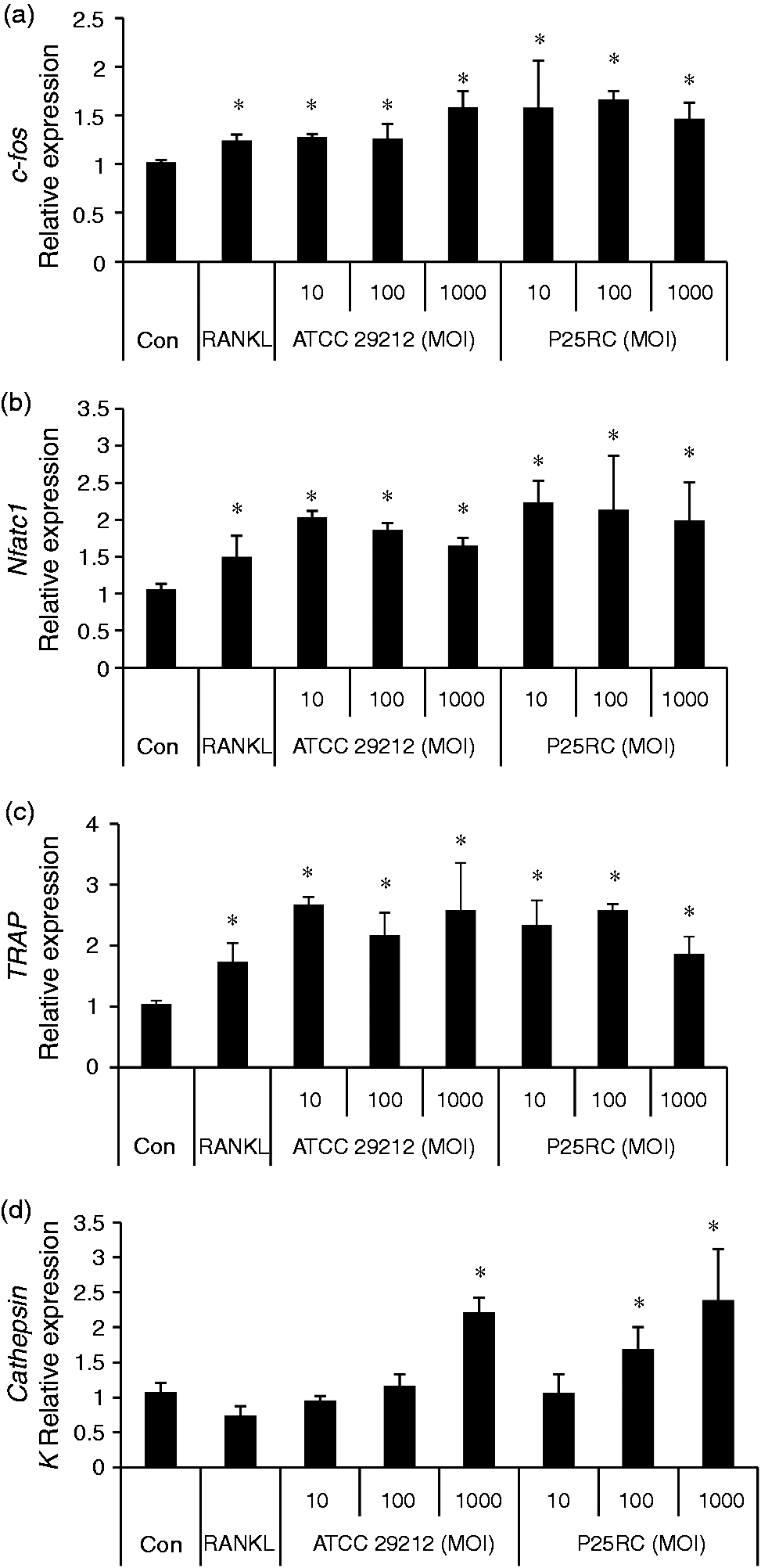
Next, RAW264.7 cells were pretreated with low-dose RANKL for 4 d followed by the treatment of two E. faecalis strains at different MOIs for 2 d. RAW264.7 cells in the absence of E. faecalis were cultured for 6 d as a control. The last 2-d culture supernatants were collected and the levels of TNF-α and IL-6 were significantly up-regulated with exposure to E. faecalis, according to the ELISA assays (Figure 4A, C), but the expression of IL-1β was not promoted (Figure 4B). Gene expressions of COX-2, c-Fos, NFATc1 and TRAP were significantly up-regulated in the presence of E. faecalis at different MOIs (Figures 4D and 5A–C). In addition, the gene expression of cathepsin K was up-regulated in RANKL-primed cells with the treatment of E. faecalis in a dose-dependent manner, which was significantly higher at the MOI of 1000 compared with the control group (Figure 5D).
The pro-inflammatory effects of E. faecalis on RANKL-primed RAW264.7 cells. RAW264.7 cells were primed with low-dose RANKL (20 ng/ml) for 4 d and then treated with the two strains of E. faecalis at different MOIs for 2 d. The last 2-d culture supernatants were collected and the levels of (A) TNF-α, (B) IL-1β and (C) IL-6 were detected using ELISA assays. (D) The gene expression of COX-2 was assayed using real-time PCR. The mean and SD are shown. *P < 0.05 was considered statistically significant. Con: untreated RAW264.7 cells (control). Gene expression of osteoclast differentiation induced by E. faecalis in RANKL-primed RAW264.7 cells. RAW264.7 cells were treated with low-dose RANKL (20 ng/ml) for 4 d and then treated with the two strains of E. faecalis at different MOIs for 2 d. The expressions of osteoclast-related genes (A) c-Fos (A), (B) NFATc1, (C) TRAP and (D) cathepsin K were assayed using real-time PCR. The mean and SD are shown. *P < 0.05 was considered statistically significant. Con: untreated RAW264.7 cells (control).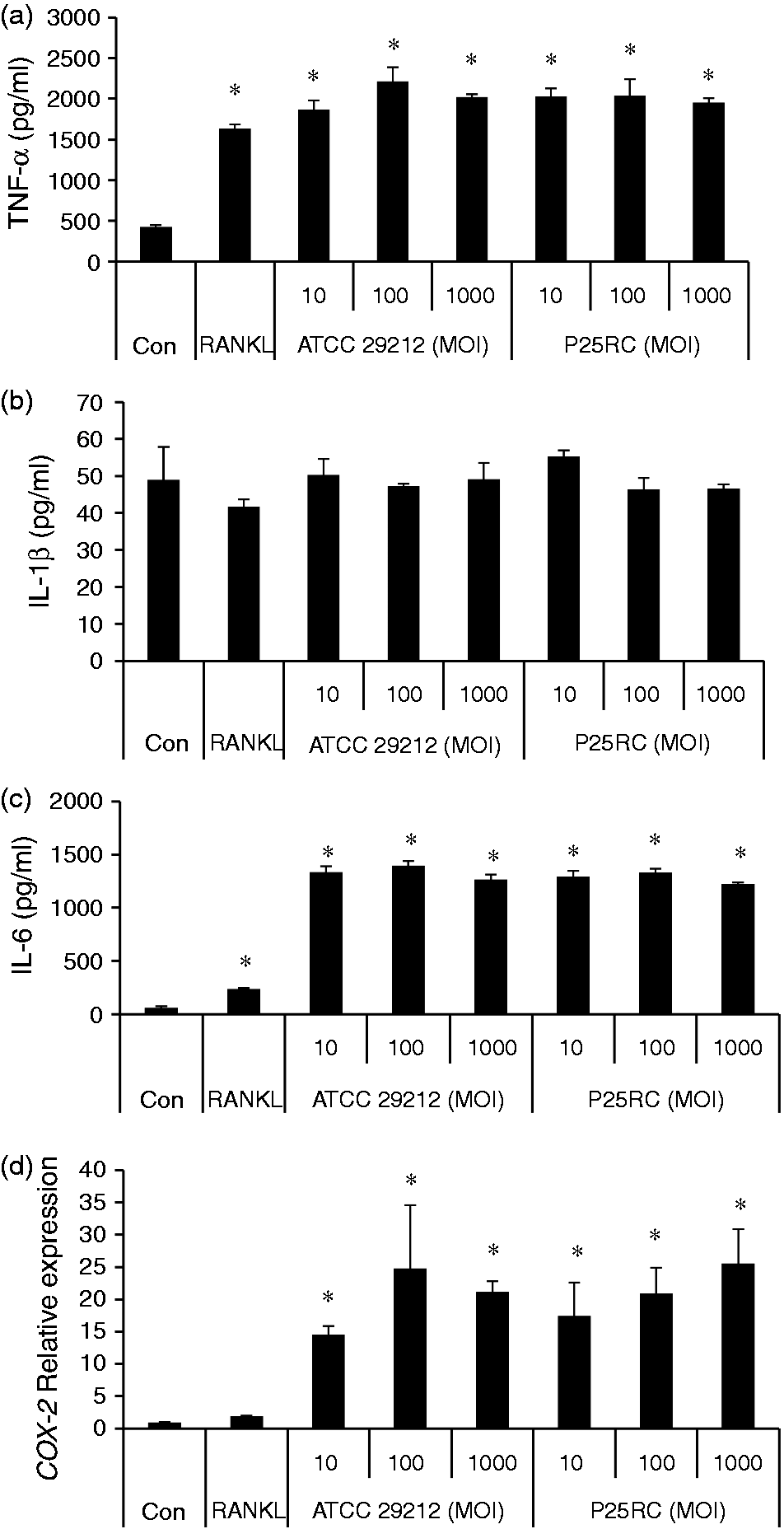
Activation of MAPK signaling pathways in RANKL-primed cells
RAW264.7 cells were pretreated with RANKL for 4 d followed by the treatment with two strains of E. faecalis at different MOIs for 2 d. RAW264.7 cells cultured for 6 d in the absence of E. faecalis served as the control. Enterococcus faecalis increased the activation of p38 and ERK1/2 compared with the control. Band density analysis indicated that E. faecalis ATCC 29212 and P25RC increased phosphorylated p38 up to 1.4- and 1.7-fold, respectively, at an MOI of 1000 compared with control. E. faecalis ATCC 29212, and also markedly increased phosphorylated ERK1/2 up to 2.1- and 3.9-fold, respectively, at an MOI of 1000 compared with the control (Figure 6).
MAPK signaling pathways of osteoclast differentiation induced by E. faecalis in RANKL-primed RAW264.7 cells. RAW264.7 cells were treated with low-dose RANKL (20 ng/ml) for 4 d and then treated with the two strains of E. faecalis at different MOIs for 2 d. The phosphorylations of p38 and ERK1/2 with regard to MAPK signaling pathways were analyzed by Western blotting. The numbers above the bands denote the relative density value.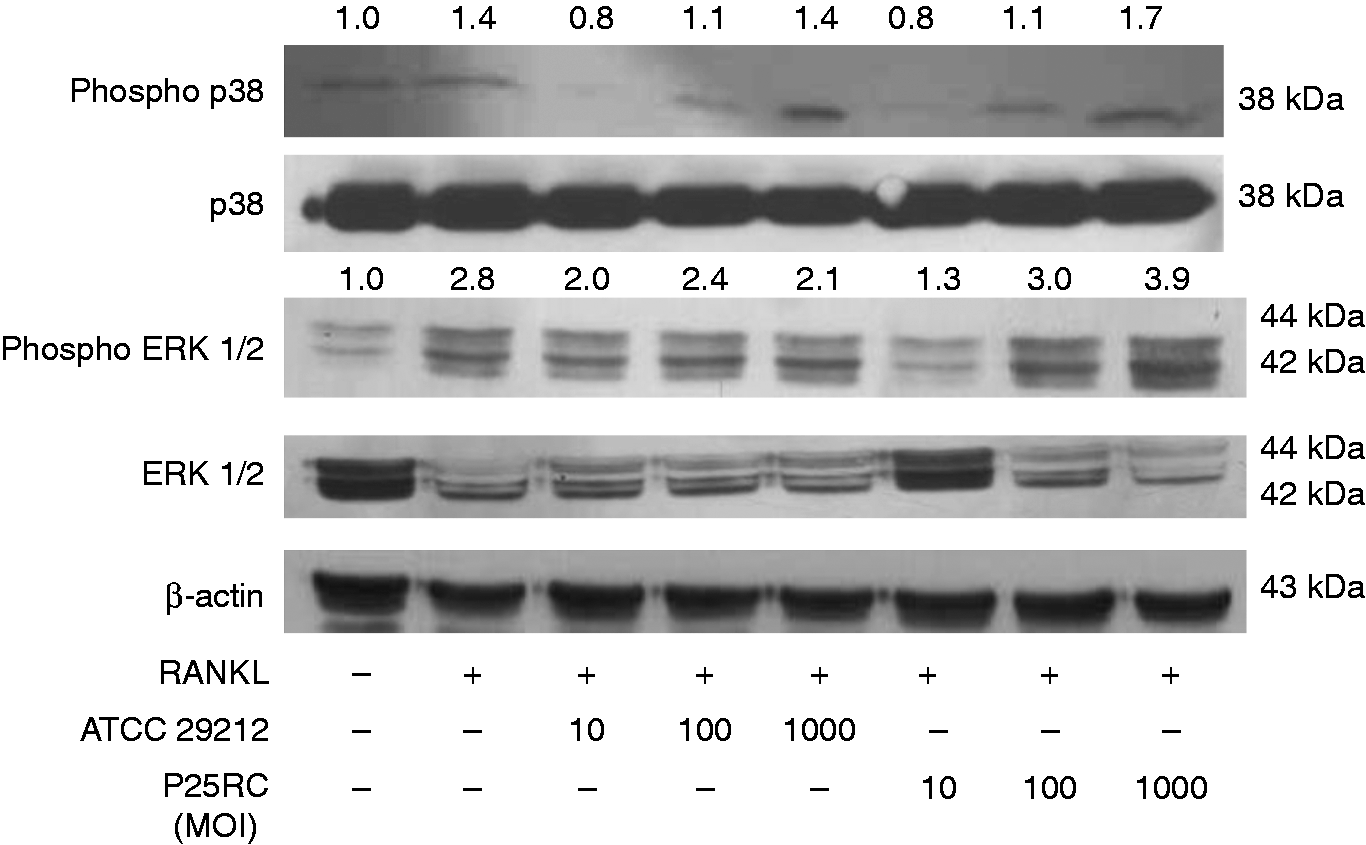
Expression of Sema4D in RANKL-primed cells
Western blotting assay revealed that the expression of Sema4D in RAW264.7 cells was down-regulated when treated with RANKL. However, E. faecalis treatment significantly enhanced Sema4D expression in RANKL-primed RAW264.7 cells. The highest expression of Sema4D was found when cells were treated with E. faecalis P25RC at an MOI of 1000 (1.1-fold compared with control) (Figure 7A). The ELISA results also showed that soluble Sema4D increased with the improvement of MOI of E. faecalis in RANKL-primed cells (Figure 7B).
Sema4D expression of osteoclast differentiation induced by E. faecalis in RANKL-primed RAW264.7 cells. RAW264.7 cells were treated with low-dose RANKL (20 ng/ml) for 4 d and then treated with the two strains of E. faecalis at different MOIs for 2 d. (A) The expression of Sema4D was assayed by Western blotting. The numbers above bands denoted the relative density value. (B) The soluble Sema4D was detected by ELISA. The mean and SD are shown. *P < 0.05 was considered statistically significant.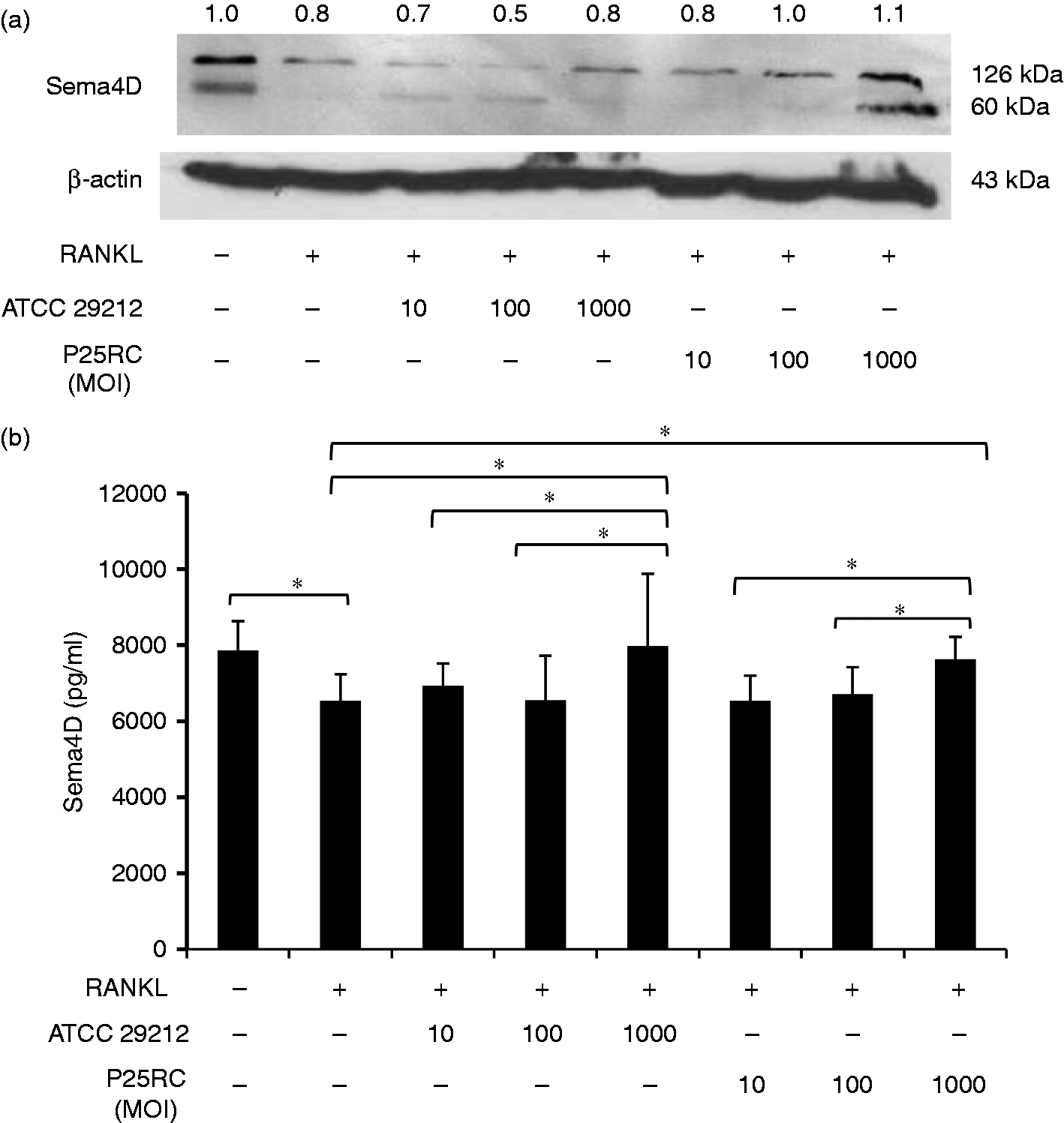
Discussion
The present study shows that E. faecalis markedly stimulated RAW264.7 cells to undergo osteoclast differentiation through a RANKL-dependent mechanism. Previous studies have demonstrated that pro-inflammatory cytokines such as TNF-α and IL-1β play a role in osteoclastogenesis and bone resorption.29,30 In addition, the COX-2 pathway is crucial for inflammatory processes by contributing to the synthesis of the pro-inflammatory mediator prostaglandin E2 (PGE2), which is responsible for osteoclastogenesis and bone resorption.24,31,32 Therefore, bacteria or their products may mediate osteoclast differentiation via pro-inflammatory mediators such as TNF-α, IL-1β, IL-6 and PGE2.33–35 It has been reported that TNF-α and IL-6, but not IL-1β, are responsible for lipopeptide-induced osteoclast differentiation. 36 Concordantly, our study indicates that E. faecalis could significantly enhance the expressions of TNF-α, IL-6 and COX-2 in RANKL-primed RAW264.7 cells, thereby contributing to the bone resorption by inducing pro-inflammatory mediators involved in osteoclastogenesis and bone resorption. In addition, Gram-positive bacteria are commonly recognized by TLR2, and the activation of TLR2 can cause an immune response via multiple signaling pathways. 37 In our study, the blocking assay showed that the expression of TNF-α induced by E. faecalis was attenuated by blocking TLR2 in RAW264.7 cells.
There are four phases of osteoclast development: (i) cell–cell fusion of osteoclast precursors, (ii) osteoclast formation, (iii) osteoclast activation and (iv) osteoclast survival. 38 Only mature osteoclasts have the capacity to resorb bone.38,39 In fact, the interaction of RANKL with its receptor RANK plays an important role in osteoclast differentiation. 40 It has been reported that c-Fos can regulate the expression of NFATc1 through binding to the promoter sequence of NFATc1 gene. 41 NFATc1 is an osteoclast marker involved in cell fusion, and plays an essential role in osteoclastogenesis.9,42 Our results demonstrate that the expression of c-Fos and NFATc1 was higher in RANKL-primed RAW264.7 cells treated with E. faecalis. However, RANKL resulted in the expression of TRAP in RAW264.7 cells. 43 We found that RANKL-primed RAW264.7 cells treated with E. faecalis could form more TRAP-positive multinucleated cells. However, treatment with E. faecalis at an MOI of 10 significantly enhanced TRAP expression in RANKL-primed RAW264.7 cells. There was no significant difference in the gene expression of cathepsin K between RAW264.7 cells with or without RANKL. Those with an MOI of 1000 resulted in prominent expression of cathepsin K in RANKL-primed RAW264.7 cells. It has been reported that the expression of TRAP and cathepsin K could be up-regulated in the early stage of osteoclastogenesis. 43 TRAP and cathepsin K are good indicators of active bone resorption.15,16 Hence, our current findings indicate that E. faecalis could promote osteoclast differentiation and resultant bone resorption. The up-regulation of NFATc1, TRAP and cathepsin K suggests that E. faecalis may have a potent effect on osteoclastogenesis, especially with pretreatment of RANKL in osteoclast precursors.
MAPK signaling pathways are essential for osteoclast differentiation and activation. 44 It has been shown that p38 and ERK1/2 are partially involved in osteoclast differentiation in RANKL-primed precursors. 45 In the present study, we also found that p38 and ERK1/2 signaling pathways could be activated by E. faecalis in RANKL-primed RAW264.7 cells, which may contribute to the osteoclast differentiation from RAW264.7 cells.
Sema4D is a transmembrane protein released by osteoclasts and osteoclast precursors. It is worthy to note that both strains of E. faecalis enable promotion of the expression of transmembrane and soluble Sema4D. Sema4D suppresses bone formation via binding to its receptor Plexin-B1 derived from osteoblasts. Our study shows that when RAW 264.7 cells were induced by RANKL into osteoclast-like cells, the expression of Sema4D was transiently decreased. This phenomenon may result from the reduction of the number of total cells by cell–cell fusion during the early stage of osteoclast differentiation. Collectively, E. faecalis could induce osteoclastogenesis while suppressing osteoblast differentiation by up-regulating Sema4D expression, subsequently leading to bone resorption.
In conclusion, the present study unravels for the first time an undescribed mechanism by which E. faecalis may contribute to the common bone resorption observed in periapical periodontitis, by promoting RANKL-dependent TRAP-positive osteoclastogenesis and expression of Sema4D through activation of the p38 and ERK1/2 MAPK signaling pathways. This novel mechanism may shed light on the underlying pathogenic mechanism behind periapical periodontitis, and contribute to developing a new treatment strategy for effective management of this common oral disease.
Footnotes
Acknowledgements
We thank Raymond Tong, Edith Tong and Wendy Yu for their technical assistance.
Funding
This work was supported by RFCID/HHSRF Grant No.12110772.
Conflicts of interest
The authors do not have any potential conflicts of interest to declare.