
Abstract
Neisseria meningitidis causes fulminant meningococcal sepsis with a massive activation of the coagulation and complement cascades. Bacterial cell envelope molecules from N. meningitidis, particularly lipopolysaccharide (LPS), induce tissue factor (TF) expression. In meningococcal sepsis, TF can be detected on circulating monocytes and microparticles (MPs) within the bloodstream. During infection, Nm activates C5 and C5a, which also is able to induce TF. We evaluated the effect of eculizumab, a C5-blocking monoclonal antibodies (mAb), on cell- and MP-associated TF. Using a lepirudin-anticoagulated whole blood model, we activated the coagulation and complement cascades by N. meningitidis, and investigated the interaction between the cascade systems with special focus on cell-associated TF-expression (mRNA and protein) and MP-associated TF-dependent thrombin and fibrin generation in platelet-free plasma. We also examined the ability of TF-positive MPs to support clot formation in whole blood. In addition, the effect of corn trypsin inhibitor and time-dependent changes on MP-associated functional TF activity was examined. Inhibition of C5 reduced cell-associated TF expression at both gene and protein level, and reduced MP-associated TF-dependent thrombin and fibrin generation in platelet-poor plasma, MP-induced TF-dependent clot formation in whole blood, implying that the complement and coagulation cascades are interplayers in N. meningitidis-mediated activation of these cascades.
Introduction
The Gram-negative bacterium Neisseria meningitidis causes fulminant meningococcal sepsis with a massive inflammatory response induced by activation of the ‘first line of defence’, including the coagulation and complement cascades. The clinical picture is explained by rapid proliferation of N. meningitidis to levels as high as 108 bacteria per ml plasma and endotoxin (LPS) levels up to 2150 endotoxin units per ml in plasma.1,2 Multiple hemorrhagic skin lesions are the hallmark of the infection and they often evolve to purpura fulminans and thrombosis of peripheral extremities. 1 Bacterial cell wall molecules from N. meningitidis, particularly LPS, induce tissue factor (TF) expression. 1 Tissue factor, a 47-ku transmembrane glycoprotein, is the main physiologic initiator of blood coagulation, 3 where the extracellular N-terminal domain contains functional sites for binding the serine protease factor VIIa (FVIIa). At the site of vascular damage, the extravascularly located TF/FVIIa cell surface complex initiates blood coagulation by limited proteolysis and activation of factor IX (FIX) and factor X (FX), which subsequently leads to the generation of thrombin, fibrin deposition and platelet activation. In meningococcal sepsis, TF can be detected on human circulating monocytes and also on cell-derived microparticles (MPs) within the blood stream. 4 MPs are small phosphatidylserine (PS)-exposing membrane vesicles (<1 µm) released from cells upon activation or apoptosis. 5 MPs may disseminate TF in addition to procoagulant phospholipids, and can be incorporated into developing thrombi. 6 Owing to its possible role as a pro-thrombotic biomarker in a variety of diseases such as meningococcal sepsis and cancer, there is an increasing clinical interest for measuring MP-associated TF activity.5,7 However, to be able to use TF-positive MPs as biomarkers for identifying thrombotic complications, accurate and standardized measurements of blood-borne TF are required. 8
During infection, N. meningitidis activates the serine proteases involved both in the coagulation and the complement cascades. 9 The activation of the complement cascade is evident in severe meningococcal sepsis and is related to disease severity. 10 Generation of the powerful complement anaphylatoxins C3a and, in particular, C5a, contributes substantially to the inflammatory reaction.11–13 A final assembly of the activated components from the complement system leads to the formation of the terminal C5b-9 complement complex (TCC) that inserts into the bacterial membrane and kills the N. meningitidis. C5a has been shown to exert many harmful effects that may contribute to immunoparalysis, multi-organ failure and septic cardiomyopathy, as well as imbalance in the coagulation system.14,15 C5a may also induce up-regulation of TF. Ikeda et al. 16 found that recombinant human C5a induced TF in a dose-dependent way in HUVEC cells, thereby probably contributing to the activation of the extrinsic coagulation pathway. Thus, C5a may represent one of the many potential crosstalk links between the coagulation and the complement systems.14,16,17 Blocking the C5a receptor in experimental sepsis in rodents significantly improved survival, indicating an important role of complement activation in sepsis. 18 The recombinant mAb eculizumab specifically binds to C5, inhibiting its cleavage by the C5 convertase, thereby preventing the generation of C5a and TCC. This Ab has been shown to reduce thrombotic complications in diseases such as paroxysmal nocturnal hemoglobinuria and atypical hemolytic uremic syndrome.19,20
The aim of this study was to evaluate the effect of complement on cell- and MP-associated TF using the C5 inhibitory mAb eculizumab, which is in clinical use. For this purpose we incubated N. meningitidis in lepirudin-anticoagulated human whole blood and investigated the interaction between the two systems with special focus on TF expression and function.
Material and methods
Ethics statement
The project was approved by the Regional Committee for Medical Research Ethics, South-Eastern Norway Regional Health Authority, Oslo, Norway.
Reagents and equipment
Equipment such as tubes, tips, and solutions were endotoxin-free. CryoTubes (polypropylene tubes; Nalgene Nunc, Roskilde, Denmark) were used to obtain low background activation of complement. Lepirudin (Refludan 50 mg) was obtained from Celgene (Munich, Germany). Eculizumab (Soliris), a mAb that specifically binds to the complement protein C5, was purchased from Alexion Pharmaceuticals (Cheshire, CT, USA). Sterile Tris-buffered saline (TBS) and BSA were purchased from Sigma Aldrich (St. Louis, MO, USA). The FluCa-kit and Thrombin Calibrator were from Thrombinoscope BV (Maastricht, the Netherlands). Anti-TF mAb-producing cells [TF8-5G9 and TF9-5B7 (both IgG1)] were kind gifts from Professor James H. Morrissey at University of Illinois, College of Medicine (Urbana, IL, USA) and the cells were further processed for production of anti-TF Abs by Dr Elisabeth Paus, Department of Medical Biochemistry, Oslo University, Norway. A mixture of 1 µl TF9-5B7 (0.84 mg/ml) and 2 µl TF8-5G9 (0.40 mg/ml) was used in the experiments. The N. meningitidis strain H44/76 is a serogroup B (B:15:p1.16:L,3,7,9) reference strain initially isolated from a Norwegian patient with fulminant septicaemia. 21 The meningococci were grown as previously described. 22 The bacterial suspension was heat-inactivated at 56℃ for 30 min, and the number of bacteria determined by quantitative PCR. 2 A stock solution of 1 × 1011/ml bacteria was stored at –80℃ and used at a final N. meningitidis concentration of 1 × 108 bacteria/ml blood.
N. meningitidis whole blood model
The lepirudin-based human whole blood model, leaving the complement system active and able to mutually interact with the rest of the inflammatory systems, has been described in detail elsewhere. 23 The model was adapted for incubation with N. meningitidis in the present study. Whole blood from consenting, healthy donors was collected in sterile polypropylene tubes containing lepirudin (50 µg/ml). Aliquots of whole blood were pre-incubated for 5 min at 37℃ with eculizumab (100 µg/ml final concentration) or TBS. Then the samples were incubated ± 108 N. meningitidis/ml for 4 h at 37℃ under constant rotation [12 rpm, (MACSmix; Miltneyi Biotec, Bergish Gladbach, Germany)]. After incubation, 500 µl of whole blood was transferred to 1.38 ml of PAXgene solution and stored frozen until qRT-PCR (TF mRNA expression). For analysis of TCC, lepirudin-anticoagulated plasma was obtained from whole blood after centrifugation [2000 g, 15 min, room temperature (RT; 22–25℃)]. Then, 5 µl EDTA (0.5 M Ultrapure, pH 8.0; Gibco, Paisley, UK) was added to aliquots of 250 µl plasma and stored frozen until analysis.
To measure MP-associated TF-activity, aliquots of 550 µl lepirudin-anticoagulated plasma were transferred to new tubes and stored frozen. Subsequently, at the day of analysis, the aliquots were thawed (10 min, 37℃), centrifuged (2000 g, 15 min, RT) and the top layer (500 µl) transferred to new Eppendorf tubes. To pellet MPs, plasma was further centrifuged (17,000 g, 30 min, RT), the top layer removed and the remaining MP-enriched pellets were washed twice (17,000 g, 30 min, RT). Then, MP pellets were resuspended in either 150 µl barbital buffer diluted in 0.9% (v/v) saline (1:10) with 0.1% (v/v) BSA for the fibrin generation assay (FGA) or in 150 µl Tris-buffer with 0.5% (v/v) BSA (TBSA) for the thrombin generation assay (CAT) and for clot formation in whole blood [rotational thromboelastometry (ROTEM)].
Preparation of pooled normal plasma
To prepare pooled normal plasma (PNP) for use in the CAT assay, whole blood was collected into citrate-anticoagulated SCAT tubes [0.109 M Na-citrate and 50 µg/ml corn trypsin inhibitor (CTI); Haematologic Technologies, Essex Junction, VT, USA] from five healthy volunteers by venipuncture (21-G needle). The first tube was discarded and the plasma prepared by centrifugation (2000 g, 15 min, RT), followed by pooling of the plasma samples. Then, PNP was subjected to sequential centrifugations 2000 g, 15 min, RT and 48,000 g, 1 h, RT. The PNP was aliquoted and stored at –80℃ until use.
For the FGA assay, blood was collected into citrate-anticoagulated tubes (0.109 M; Greiner Bio-One, Frickenhausen, Germany) and PNP further prepared as described above.
FGA (fibrin generation assay)
25 µl of MP-enriched pellets were transferred to 96-well plates in duplicates (Cell culture cluster, Costar 3595, Corning, Tewksbury, MA, USA). PNP (75 µl) was added to the wells (4 min, 37℃), and the clotting reaction initiated with CaCl2 (100 µl, 10 mmol/l final concentration). Optical density (OD) changes (340 nm) were recorded kinetically every 9 s for 20 min (VersaMax, Molecular Devices, Sunnyvale, CA, USA), giving the clot formation time in seconds. To inhibit TF-dependent clot forming activity, 3 µl of anti-TF mAbs were incubated with the MP-enriched pellets 15 min before PNP was added.
CAT (calibrated automated thrombogram)
10 µl of MP-enriched pellets were added to separate wells containing either 20 µl TBSA ± anti-TF Abs or 20 µl calibrator. Then, 70 µl PNP was added to all wells and incubated for 15 min at 37℃ before the reaction was initiated by automated addition of Fluobuffer containing calcium and a fluorogenic substrate. The samples were run in duplicate wells and the calibrator as singlet (Thermo Immulon 2HB plate; Thermo Scientific, Billerica, MA, USA). The fluorescence was read by a Fluoroscan Ascent (Thermo Scientific), and the thrombin generation parameter lag time was calculated in minutes by the Thrombinoscope software (Thrombinoscope BV).
ROTEM (rotational thromboelastometry)
12 µl of washed MP-enriched pellets were mixed with 3 µl TBSA or 3 µl anti-TF Abs and incubated for 10 min in a 1.5 ml Eppendorf tube. Then, the MP pellets were added to 335 µl citrate/CTI-anticoagulated whole blood drawn from healthy donors in tubes containing 0.109 M citrate and 50 µl/ml CTI (Haematologic Technologies). Owing to time-dependable changes in the clotting time (CT), the measurements were performed between 1 and 4 h after the blood was drawn (see below). The ROTEM experiments were also analyzed using a CTI tube never exposed to air for each donor. To start the clotting reaction, 300 µl of the mixture of whole blood and MPs was added to pre-warmed ROTEM cups (Cup & Pin pro, Tem Innovations, Munich, Germany) containing 20 µl Star-tem (0.2 M CaCl2). The CT in seconds was automatically calculated by the ROTEM software (v. 1.5.0).
Effect of storage and exposure to air on citrate/CTI-anticoagulated whole blood in ROTEM
MPs obtained from whole blood exposed to ± 108 N. meningitidis/ml were added to citrate/CTI-anticoagulated whole blood as described above. To study the effect of storage of whole blood to be used in ROTEM, MPs were added to citrate/CTI-anticoagulated whole blood stored for up to 4 h. The effect of repeated uncapping of the tubes (exposure of air) containing citrate/CTI-anticoagulated whole blood was also examined. Thus, in these time-course experiments, the clotting reactions were initiated in either whole blood (stored at 37℃) from tubes only uncapped at the time of analysis or from tubes subjected to repeated capping/uncapping. The CT was calculated by the ROTEM software as described above.
Flow cytometry of monocyte TF expression
50 µl of whole blood was stained with anti-CD45-allophycocyanin (common leukocyte marker; Beckman Coulter, Brea, CA, USA), anti-CD14-Horizon V450 (monocyte marker) and anti-CD142 phycoerythrin (TF marker), both purchased from BD Biosciences (San Jose, CA, USA), and sheltered from light for 20 min at RT in 5 ml polypropylene tubes (VWR, Radnor, PA, USA). Then, samples were lysed and fixed by adding 500 µl 1 × BD FACS Lysing solution (BD Biosciences) and sheltered from light for 15 min at RT. Cells were kept on ice until analyzed with a FACSAria flow cytometer (BD Biosciences). Mononuclear cells were selected in a dotplot with sidescatter (SSC) (linear scale) on the y-axis and CD45 (log-scale) on the x-axis. A CD14 (log-scale) versus SSC dotplot was used to select the CD14-positive monocytes. Aggregated monocytes were excluded in a dotplot with SSC-pulse width on the x-axis versus CD14 (log-scale) on the y-axis. The remaining CD14 positive population was further analyzed in a dotplot displaying CD142 (log-scale) on the x-axis versus SSC (linear scale) on the y-axis. TF expression was reported as median fluorescence intensity.
TF mRNA expression (RT-qPCR)
Total RNA from whole blood lysates was extracted using the PAXgene Blood RNA Kit, according to the manufacturerś instructions (PreAnalytix, Qiagen, Crawley, UK). The isolated total RNA was quantified using the Nano Drop spectrophotometer (Thermo Scientific, Erembogem, Belgium) and quality controlled using the Agilent 2100 Analyzer (Agilent, Santa Clara, CA, USA).
Equal amounts of total RNA (100 ng in 13 µl H2O) were reverse transcribed in a total volume of 20 µl using oligo d(T) as primers and the reverse transcriptase Omniscript (Qiagen). Gene expression of TF in whole blood was quantified using the Applied Biosystems ViiA 7 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) and TaqMan probes (F3-Hs 01076029_m1). The geometric mean of β2microglobulin (B2M-Hs 00984230_m1), PPIB(PPIB-Hs 00168719_m1) and TMBIM4 (TMBIM4-Hs 00187842_m1) mRNAs were used as reference genes. 24 The relative change of the TF transcript was calculated using the ΔΔ Ct method. 25 The results were expressed as relative quantities, i.e. fold-change of TF in whole blood incubated with N. meningitidis (108/ml), normalized to the reference genes, and relative to whole blood incubated with only TBS (unexposed samples).
The TCC
The fluid-phase TCC, was analyzed in an ELISA based on a mAb (aE11).26,27
Statistics
Comparison of means was performed using paired samples t-test (SPSS PASW Statistics 18). Data were shown as mean ± SD, and P-values < 0.05 were considered statistically significant.
Results
The effect of eculizumab on N. meningitidis-induced MP-associated TF-dependent fibrin generation
MP-enriched pellets from whole blood (n = 6) incubated with TBS (unexposed), N. meningitidis (108 bacteria/ml), or N. meningitidis (108 bacteria/ml) and eculizumab were analyzed in a plasma-based FGA (Figure 1). MPs released from whole blood exposed to 108/ml N. meningitidis gave a significant shortening of the clotting time (CT) compared with MPs obtained from unexposed whole blood [N. meningitidis-exposed: 74 s ± 20 s (mean ± SD), unexposed: 202 s ± 22 s (P < 0.001)]. Eculizumab significantly lengthened the N. meningitidis-induced CT from 74 s ± 20 s to 105 ± 22s (P = 0.019). MPs obtained from whole blood only incubated with eculizumab had no effect on the CT (data not shown). Preincubating the MPs obtained from N. meningitidis-incubated whole blood with anti-TF Abs prolonged CT to the level of unexposed MPs (Figure 1), indicating a TF-dependent change in CT. In control experiments, applying the bacteria alone to the assay, the CT was at the same level as MPs obtained from unexposed blood, indicating that the bacteria did not interfere with the measurements (data not shown).
The effect of eculizumab on N. meningitidis (Nm)-induced MP-associated TF-dependent fibrin generation. Lepirudin-anticoagulated whole blood from six donors was exposed to N. meningitidis in the absence and presence of eculizumab for 4 h. TF-dependent fibrin formation was measured in washed MP-enriched pellets obtained from plasma (incubated ± anti-TF Abs), using a FGA. Data are shown as mean ± SD. *P < 0.05 compared with N. meningitidis.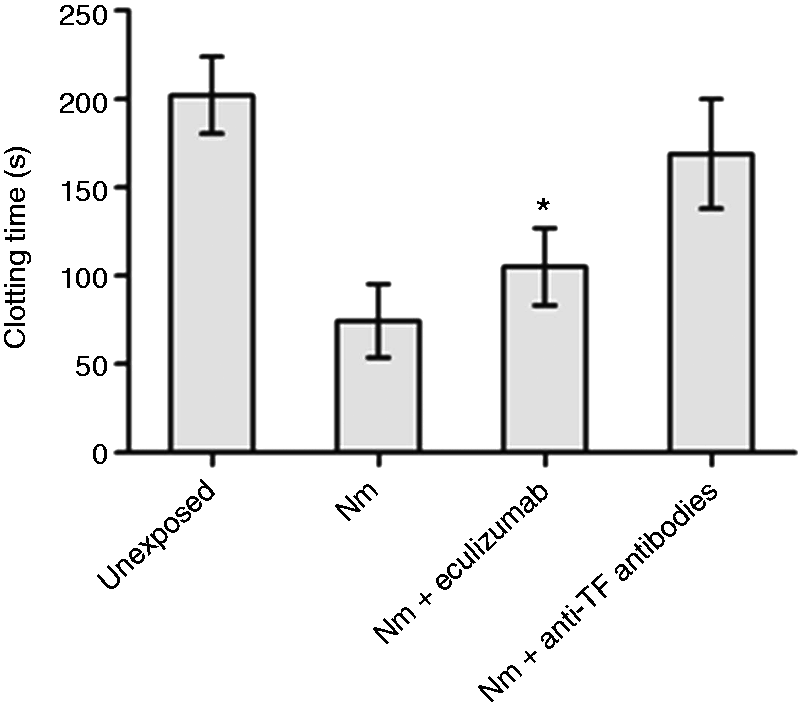
The effect of eculizumab on N. meningitidis-induced MP-associated thrombin generation
MP-enriched pellets from whole blood (n = 6) incubated with TBS (unexposed), N. meningitidis (108 bacteria/ml), or N. meningitidis (108 bacteria/ml) and eculizumab were analyzed in the CAT assay (Figure 2). MPs released from whole blood exposed to 108 N. meningitidis/ml gave a significant shortening of the lag times to 3.8 ± 1.2 min (mean ± SD) compared with lag times associated with MPs obtained from unexposed whole blood: 35.6 ± 9.9 min. Eculizumab significantly lengthened the N. meningitidis-induced lag times from 3.8 ± 1.2 min to 7.6 ± 4.5 min (P = 0.037). MPs obtained from whole blood only incubated with eculizumab had no effect on the lag times (data not shown). Pre-incubating anti-TF Abs with the MPs obtained from N. meningitidis-exposed whole blood prolonged all lag times to the level of MPs obtained from unexposed whole blood (Figure 2), indicating a TF-dependent change in lagtime. In control experiments, applying the bacteria alone to the assay, the lag time was at the level of MPs obtained from unexposed blood, indicating that the bacteria did not interfere with the measurements (data not shown).
The effect of eculizumab on N. meningitidis (Nm)-induced MP-associated thrombin generation (CAT). Lepirudin-anticoagulated whole blood from six donors was exposed to N. meningitidis in the absence and presence of eculizumab for 4 h. TF-dependent thrombin generation was measured in washed MP-enriched pellets obtained from plasma (incubated ± anti-TF Abs), using a CAT. Data are shown as mean ± SD. * P < 0.05 compared with N. meningitidis.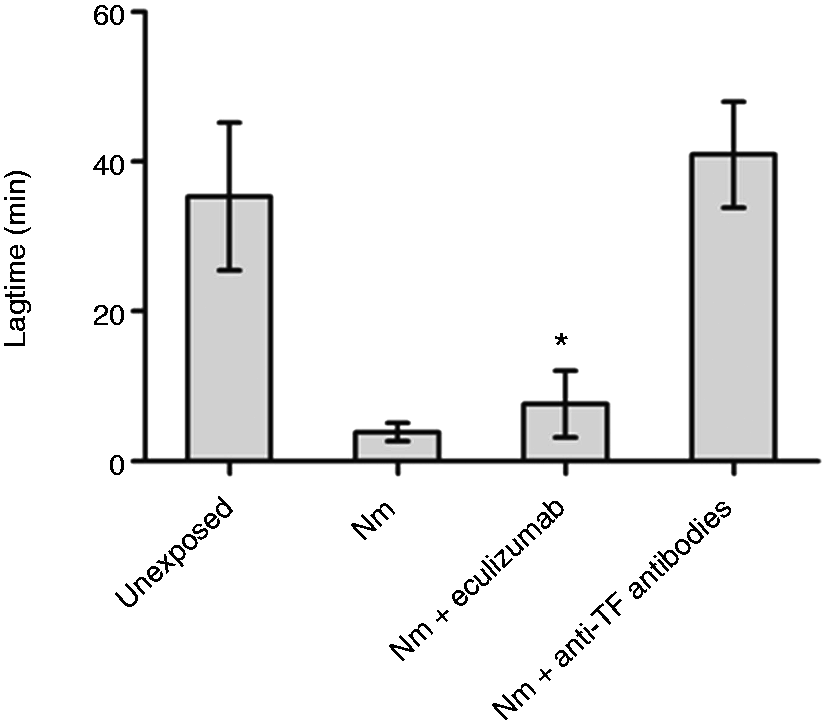
The effect of eculizumab on N. meningitidis-induced MP-associated whole blood clot formation
MP-enriched pellets from whole blood (n = 7) incubated with TBS (unexposed), N. meningitidis (108 bacteria/ml), or N. meningitidis (108 bacteria/ml) and eculizumab were added to citrated whole blood containing CTI. The CT was analyzed in a ROTEM (Figure 3). MPs released from whole blood exposed to 108 N. meningitidis/ml gave a significant (P = 0.001) shortening of CT to 303 s ± 99 s (mean ± SD) compared with MPs from unexposed blood 1015 s ± 210 s. Eculizumab significantly lengthened the N. meningitidis-induced CT from 303 s ± 99 s to 438 s ± 127 s (P = 0.012). MPs obtained from whole blood only incubated with eculizumab had no effect on the CT (data not shown). Pre-incubating the MPs from N. meningitidis-exposed blood with anti-TF Abs prolonged all the CTs (1049 s ± 122 s) to the level of MPs obtained from unexposed whole blood (Figure 3), indicating a TF-dependent change. In control experiments, applying the bacteria alone to the assay, the CT was at the level of MPs obtained from unexposed blood, indicating that the bacteria did not interfere with the measurements (data not shown).
The effect of eculizumab on N. meningitidis (Nm)-induced MP-associated whole blood clot formation (ROTEM). Lepirudin-anticoagulated whole blood from six donors was exposed to N. meningitidis in the absence and presence of eculizumab for 4 h. TF-dependent clot formation was measured in washed MP-enriched pellets obtained from plasma (incubated ± anti-TF Abs). MP-enriched pellets were added in citrate/CTI-anticoagulated whole blood and analyzed using thromboelastometry (ROTEM). Data are shown as mean ± SD. *P < 0.05 compared with N. meningitidis.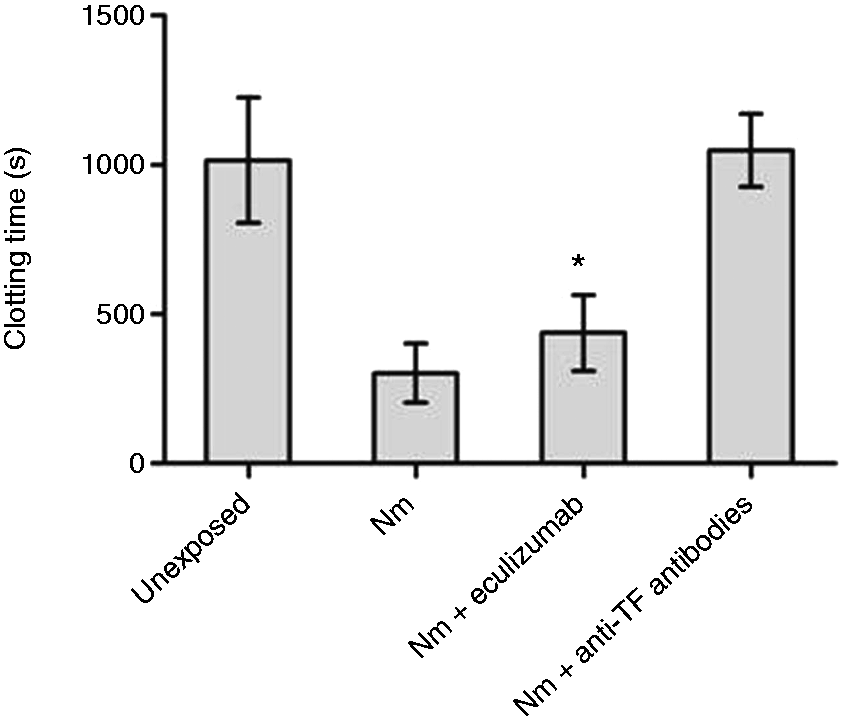
The effect on CT when citrate/CTI-anticoagulated whole blood was exposed to air
MPs obtained from whole blood (n = 4) exposed to ± 108 N. meningitidis/ml showed decreasing CT when analyzed in blood from SCAT tubes stored over a period of 0–4 h with the most profound change within the first hour. From 1 to 4 h the mean CT remained relatively stable [from 318 s to 309 s (N. meningitidis-exposed) and 1269 s to 1114 s (unexposed)] when analyzed in blood taken from a SCAT tube that had never been uncapped. Using blood from SCAT tubes that had been repeatedly exposed to air, i.e. using blood from the same tube for multiple measurements, the mean CT shortened from 309 s to 229 s (N. meningitidis-exposed) (P = 0.015 at 4 h compared with mean CT using new tubes for each measurement) and from 1352 s to 618 s (unexposed) (P = 0.002 at 4 h) (data not shown).
The effect of eculizumab on TF mRNA in whole blood
Whole blood (n = 6) incubated with TBS (unexposed), N. meningitidis (108 bacteria/ml), or N. meningitidis (108 bacteria/ml) and eculizumab were stabilized in PAXgene solution and analyzed using qRT-PCR (Figure 4). The relative quantity (RQ) TF mRNA increased significantly in all donors in the presence of N. meningitidis compared with unexposed blood (mean RQ increase from 1 to 28.2). The presence of eculizumab significantly decreased N. meningitidis induced RQ from 28.2 to 3.9 (P = 0.009). Incubation of whole blood with eculizumab alone had no effect on the RQ of TF mRNA compared with unexposed blood (data not shown).
The effect of eculizumab on TF mRNA in whole blood. Lepirudin-anticoagulated whole blood from six donors was exposed to N. meningitidis (Nm) in the absence and presence of eculizumab for 4 h. Whole blood was harvested for RNA isolation, and TF mRNA was quantified by qRT-PCR. Relative quantities of TF mRNA compared with unexposed mRNA were calculated using the ΔΔCt method. Data are shown as mean ± SD. *P < 0.05 compared with N. meningitidis.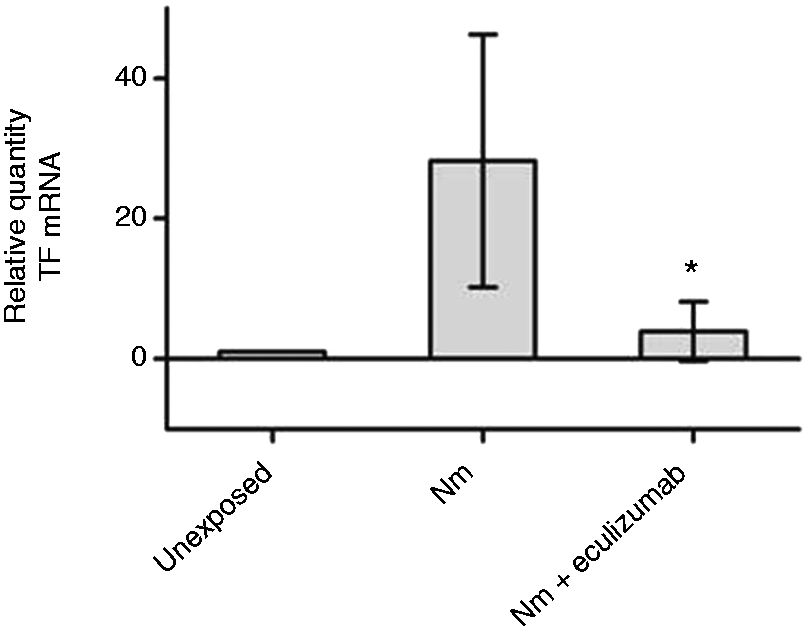
The effect of eculizumab on monocyte-associated TF expression in whole blood
Whole blood (n = 6) incubated with TBS (unexposed), N. meningitidis (108 bacteria/ml) or N. meningitidis (108 bacteria/ml) and eculizumab were analyzed using flow cytometry (Figure 5). The median fluorescence intensity (MFI) for TF expression on CD14-positive cells increased significantly in all donors in the presence of N. meningitidis compared with unexposed blood (MFI increased from 1917 to 6213, P = 0.0001). The presence of eculizumab significantly decreased the MFI (TF) of the N. meningitidis-exposed monocytes from 6213 to 4245 (P = 0.001). Incubation of whole blood with eculizumab alone had no effect on the MFI (TF) compared with unexposed blood (data not shown).
The effect of eculizumab on monocyte-associated TF expression in whole blood. Lepirudin-anticoagulated whole blood from six donors was exposed to N. meningitidis (Nm) in the absence and presence of eculizumab for 4 h. The cells were then labeled and analyzed by flow cytometry The MFI TF expression was measured on CD14-positive cells. Data are shown as mean ± SD. * P < 0.05 compared with N. meningitidis.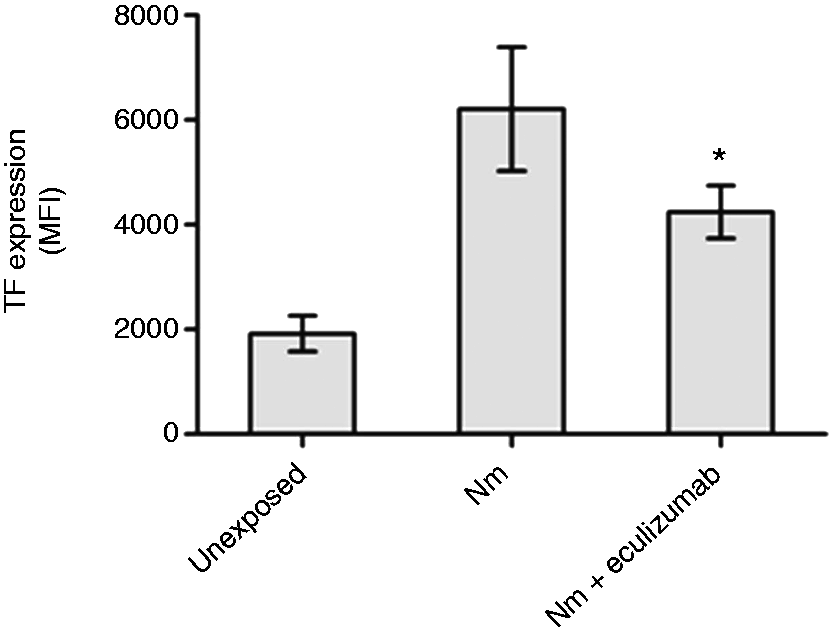
The effect of eculizumab on TCC formation
TCC was quantified in plasma samples from whole blood (n = 6) incubated with TBS (unexposed), N. meningitidis (108 bacteria/ml), or N. meningitidis (108 bacteria/ml) and eculizumab. Incubation with N. meningitidis in whole blood gave significantly higher levels of TCC than in unexposed from 16.0 to 28.2 AU/ml (P = 0.003). Eculizumab effectively inhibited the N. meningitidis-induced complement activation, reducing the levels of TCC from 28.2 to 1.2 AU/ml (P = 0.0001) (data not shown).
Discussion
In this study, we evaluated the effect of complement on cell and MP-associated TF using the C5 inhibitory mAb eculizumab. To our knowledge, we are the first to use a N. meningitidis-based whole blood model to study the interactions between the coagulation and complement cascades. We focused on functional activity of TF-positive MPs, and showed, for the first time, that the inhibition of the complement cascade reduced MP-associated TF-dependent procoagulant activity in both plasma and whole blood-based coagulation assays. In addition, eculizumab reduced cell-associated TF expression (mRNA and protein level). Our findings imply that complement activation plays an important role in N. meningitidis-induced TF mRNA regulation, and it modulates N. meningitidis-induced TF expression on monocytes and TF activity associated with MPs.
Interactions between the coagulation and the complement cascades have previously been proposed, 17 and therapeutic modulation of the human complement system is considered a promising approach for treating a number of pathological conditions. 20 Owing to their central positions in the complement cascade, complement factor C3 and C5 are attractive targets for complement-specific drugs. In a baboon model exposed to live Escherichia coli, the procoagulant response based on platelet counts and hemostatic parameters (fibrin degradation products, activated partial pro-thrombin time, soluble P-selectin and thrombomodulin) were reduced when complement was inhibited with the potent C3 convertase inhibitor, compstatin. 28 In a rat model of sepsis, treatment with anti-C5a Abs improved the hemostasis substantially, suggesting that the production of C5a caused hemostatic defects that could be reduced by blockade of C5a. 29 Furthermore, by using an E. coli-exposed whole blood model and blockage of complement factor 3, and the receptors for C5a and CD14, Brekke et al. 30 have shown that complement activation has an essential role in E. coli-induced TF mRNA up-regulation. In accordance with this study, we showed that inhibition of the complement cascade by the C5-inhibiting Ab eculizumab abolished TF mRNA up-regulation and reduced cell-associated TF expression also in N. meningitidis-induced whole blood. Brekke et al. 30 also showed that complement, together with CD14, contributes to E. coli-induced TF activity in plasma. After ultracentrifugation of MPs, TF activity in plasma was measured spectrophotometrically with pure coagulation factors in a two-stage amidolytic assay. In line with this, using plasma and whole-blood based coagulation assays, we found that TF-dependent procoagulant activity associated with MPs was significantly reduced when C5 is blocked in N. meningitidis-stimulated whole blood. Our plasma samples contain MPs from all the blood cells where the monocyte-derived MPs that express TF are most likely in rather low concentrations. Enumeration and characterization of the monocyte-derived MPs by flow cytometry is a challenging task, and even more challenging in this study owing to the fact that the bacteria, with the size of MPs, would interfere in such analysis. Thus, we are unable to conclude whether the reduction of MP-associated TF activity may be due to a reduced number of TF molecules on the microparticles or a reduced number of TF-positive microparticles.
Our TCC results showed that eculizumab effectively inhibited N. meningitidis-induced complement activation, and therefore, in our opinion, eculizumab is a valuable tool to distinguish between TF expression mediated by C5 and other pathways. Kanse et al. 13 have previously shown that C5 in polytrauma patients may form complexes with the serine protease factor seven activating protease, which may lead to generation of C5a. This may indicate that inhibition of C5 might also be an important approach in regulation of inflammation. 13
In vitro whole blood models that can be used for studying both the coagulation and the complement cascade simultaneously has been a challenge. We used lepirudin as an anticoagulant in the tube, which is the preferable anticoagulant when studying products from the complement system. However, lepirudin functions as a direct thrombin inhibitor and will therefore affect coagulation assays based on plasma or whole blood. To overcome the interference of lepirudin, we isolated MPs by pelletation, and washed the MP pellets carefully before MP-associated TF activity was measured. This isolation and washing procedure removed the thrombin-inhibiting effect of lepirudin and enabled us to study the interaction between the coagulation and the complement systems in one tube.
In contrast to the study of Brekke et al., 30 we performed MP-associated TF-dependent procoagulant activity in a plasma environment (CAT) and in whole blood (ROTEM) showing that the inhibition of C5 led to a significantly reduced MP-associated TF-dependent procoagulant activity in all assays. A challenge with such coagulation assays is the possible interference of the contact activation pathway, which may be eliminated by the use of CTI, an inhibitor of FXII.31,32 The inhibition of the contact activation pathway in the CAT assay has previously been shown to be important only when samples with low amounts of TF are analyzed. We performed the thrombin generation and whole blood clot formation experiments both with and without CTI. In accordance with previous studies, we found that the lag times and CTs were significantly prolonged in samples with low TF (MPs isolated from unexposed whole blood). However, working with high TF concentrations, such as samples stimulated with N. meningitidis 108/ml (with or without the addition of eculizumab), the lag times and CTs seemed to be almost unchanged after the addition of CTI (data not shown). Although not essential for the interpretation of our data on the effect of eculizumab, we chose to use CTI because of the effect on the samples with low TF. Furthermore, van der Meijden et al. 33 have recently shown that PS-positive MPs from erythrocytes and platelets, which are heavily abundant in whole blood, also are able to initiate coagulation via a FXII-dependent mechanism. The contribution of these PS-positive MPs to the overall thrombin generation may therefore be inhibited by the use of CTI. Thus, analyzing functional blood-borne TF, the use of CTI is debatable, but if the aim is to study only the contribution from the TF-bearing MPs, the report by van der Meijden et al. 33 may imply that CTI should be used.
Using the non-activated TEM (NATEM) approach in ROTEM enabled us to investigate the ability of TF-positive MPs to initiate thrombin generation and contribute to clot formation in an environment where platelets, leukocytes and erythrocytes are present. However, this analysis is sensitive to changes in time, pH and temperature. 34 Previous reports have shown that changes of sample pH and temperature in whole blood affect TEG (thrombelastograph) variables in a hypercoaguable direction.34,35 There have also been reported variances in TEG variables dependent on time between venipuncture of blood and analysis of sample. 36 By adding MPs to citrated/CTI-containing blood we found that the blood should rest for 1 h and be used within 4 h of analysis in the ROTEM. Previous reports have also shown that resting times from 30 min to 1 h are needed before analysis. 36 We also investigated the effect of whether citrate/CTI-anticoagulated blood repeatedly exposed to air (uncapped and capped tubes) influenced the thromboelastogram. We found that MPs subjected to citrate/CTI-anticoagulated whole blood from repeatedly uncapped/capped tubes showed a more evident increase in hypercoagulability with time than MPs put into citrate/CTI-anticoagulated blood not subjected to air before the time point of analysis. Thus, using the NATEM approach in ROTEM for MP analysis, MPs added into citrated/CTI whole blood should be analyzed within 1–4 h of venipuncture, and the tubes should only be uncapped at the time point of analysis to minimize artificial hypercoaguability.
In conclusion, by exposing whole blood anticoagulated with lepirudin to N. meningitidis, we were able to study the interaction between the coagulation and complement cascade. We showed that inhibition of C5 reduced cell-associated TF expression both at the gene and protein level. Inhibition of C5 also reduced MP-associated TF-dependent thrombin and fibrin generation in platelet-poor plasma and MP-induced TF-dependent clot formation in whole blood, reflecting the interplay between complement and coagulation in N. meningitidis-induced plasma cascade activation.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Acknowledgements
We thank Anne Pharo, Department of Immunology, Oslo University Hospital, for quantifying TCC by ELISA.