
Abstract
This study determined the influence of physiologically relevant concentrations of C-reactive protein (CRP) on reactive oxygen species (ROS) production by neutrophils. Neutrophils from healthy individuals were incubated with soluble pentameric CRP prior to TLR stimulation with Fusobacterium nucleatum, or FcγR stimulation with IgG-opsonised Staphylococcus aureus or heat-aggregated IgG. ROS generation by unstimulated cells and those after stimulation were determined using luminol, isoluminol and lucigenin chemiluminescence, detecting predominantly intracellular hypochlorous acid (HOCl), extracellular hydrogen peroxide (detected as HOCl) and extracellular superoxide respectively. Baseline (unstimulated) neutrophil ROS generation and release was reduced compared with vehicle control by 10 µg/ml CRP. There was no consistent effect of CRP on FcγR-stimulated HOCl production, but the extracellular superoxide response was reduced by 10 µg/ml CRP. By contrast, CRP reduced intracellular (10 µg/ml) and extracellular (3 and 10 µg/ml) HOCl generation, but increased superoxide release (1–10 µg/ml) in response to TLR stimulation. Physiologically relevant concentrations of CRP inhibited baseline ROS generation and reduced FcγR-stimulated extracellular superoxide and TLR-stimulated HOCl release, suggesting that CRP may offer some degree of host protection from neutrophil-associated, low-level oxidative stress. However, CRP enhanced TLR-mediated superoxide release from neutrophils, potentially increasing oxidative stress but aiding host protection from infection.
Introduction
Neutrophils are short-lived defence cells 1 classically characterised by their ability to phagocytose, to release lytic enzymes and other antimicrobial peptides, and to produce reactive oxygen species (ROS). 2 Crucial to the production of superoxide (O2−) by activated neutrophils is NADPH oxidase, a highly regulated enzyme complex composed of cytosolic (e.g. p40phox, p47phox, p67phox) and membrane-bound (e.g. p22phox, gp91phox) proteins. 3 O2− is short-lived and dismutates to hydrogen peroxide (H2O2) and forms other downstream secondary ROS. 4 Although H2O2 alone is microbicidal, its bactericidal activity is greatly potentiated by the enzyme myeloperoxidase, resulting in the formation of hypochlorous acid (HOCl). 5 Hypochlorous acid is also essential for neutrophil extracellular trap (NET) formation, 6 a novel defence mechanism whereby neutrophils extrude their nuclear chromatin in complex with granular and cytosolic components forming an extracellular DNA fibrillary network which localises their antimicrobial peptides and subsequently traps and destroys microbes. 7
While neutrophil ROS generation is primarily a defensive response, excess production can lead to oxidative stress and collateral host tissue damage, which is important in the pathology of many chronic inflammatory diseases, including atherosclerosis, 8 cardiovascular disease, 9 diabetes mellitus, 10 rheumatoid arthritis, 11 stroke 12 and chronic periodontitis. 13 The latter is the most common inflammatory disease of humans which, apart from its primary impact on oral health and healthcare costs, also has significant importance as a risk factor for many other inflammatory diseases, including atherogenic vascular disease, diabetes and rheumatoid arthritis.14–16 The pathogenesis of periodontitis involves a host response to subgingival plaque bacteria (e.g. Porphyromonas gingivalis and Fusobacterium nucleatum) 17 characterised by exaggerated or ‘hyper’ inflammation, which is ineffective in removing the causative organisms resulting in chronic non-resolving inflammation and tissue damage due to excessive or prolonged release of neutrophil proteolytic enzymes, pro-inflammatory cytokines and ROS.13,18 Peripheral blood neutrophils in periodontitis patients are both hyperactive and hyper-reactive with respect to ROS generation19,20 and, although Fcγ-receptor (FcγR)-mediated hyper-reactivity is reduced by successful periodontal therapy, spontaneous, unstimulated ROS release is not, indicating that both reactive and constitutive mechanisms contribute to tissue damage and disease progression in susceptible individuals. 21 These data are supported by the recent demonstration that shorter peripheral blood leukocyte telomere lengths are associated with a diagnosis of periodontitis, and their measures correlate with oxidative stress and severity of periodontal disease. 22 Chronic periodontal disease is also strongly associated with elevated plasma C-reactive protein (CRP) levels 23 with a consensus view that underpinning mechanisms likely relate to chronic entry of periodontal bacteria and their products into the circulation during eating and tooth cleaning. 24
The ability of neutrophils to produce ROS may also be influenced by the host via exposure to priming agents in the plasma. 25 Priming neutrophils for an enhanced respiratory burst when exposed to an activating second stimulus, could increase neutrophil-derived ROS release, both locally in tissues and also within the systemic circulation. Priming agents include pro-inflammatory cytokines, 26 platelet activating factors27,28 as well as other components resulting from the systemic acute phase response that characterises many inflammatory processes.29,30 One possible candidate is CRP, levels of which are regarded as a sensitive, systemic, non-specific marker of inflammation.31–33
CRP in human plasma is composed of five identical, non-covalently-linked subunits. 34 In health, the median concentration of CRP is 0.8 µg/ml but, following an acute phase stimulus, values may increase to >500 µg/ml. 35 Relatively moderate increases in plasma CRP are considered a risk factor for atherosclerosis, cardiovascular disease, myocardial infarction and stroke, independently of established common risk factors.36,37 It is suggested that lowering the burden of subgingival periodontal bacteria, and the associated chronic local and systemic inflammatory response through effective management of chronic periodontitis, may reduce the associated risk of atherosclerotic complications.38,39 Plasma CRP concentrations in periodontitis patients (mean high sensitivity CRP range ∼1–4 µg/ml)40–42 fall within the ‘medium cardiovascular risk range’, as defined by the American Heart Association guidelines (0.1, 1 and 3 µg/ml plasma CRP represent low, medium and high cardiovascular risk respectively).36,43
Soluble, pentameric CRP has been shown to bind rapidly and specifically to all three classes of human FcγR (FcgRI, RII and RIII) in a calcium- and magnesium-dependent manner.44–46 However, in terms of CRP binding to neutrophils, which normally only express low/medium affinity receptors for IgG (FcγRII and FcγRIII), most studies have highlighted the role of FcγRIIA. 47 Soluble, pentameric CRP has been shown to have multiple effects on neutrophils, including inhibiting chemotaxis,48,49 promoting phagocytosis50,51 and reducing the stimulated respiratory burst.44,51,52 These latter studies have shown that pre-incubation of neutrophils with CRP (1–100 µg/ml) causes a reduction (5–55%), but not complete inhibition, of O2- release after stimulation with phorbol myristate acetate (PMA), concanavalin A (Con A) and N-formyl-methionyl-leucyl-phenylalanine (fMLP). The effects of CRP on intracellular changes in ROS generation have not been investigated. Furthermore, no studies have investigated the direct effect of native CRP on baseline, unstimulated neutrophil ROS generation and release.
Neutrophil hyperactivity and reactivity, in terms of ROS generation and release, characterise individuals susceptible to periodontitis, which is associated with small increases in circulating CRP levels and known to be a risk factor for many inflammatory diseases where oxidative stress is thought to be important in their pathogenesis. To date, the ability of native CRP, at levels comparable with those detected in chronic low level inflammatory conditions, such as periodontal disease (∼1–4 µg/ml),40–42 and equivalent to those associated with increased risk of cardiovascular disease (1–3 µg/ml),36,43 to modulate ROS production by unstimulated neutrophils or those stimulated with biologically relevant stimuli has not been reported. We hypothesised that soluble, native CRP alters the level of neutrophil ROS generation in the absence of deliberate stimulation, as well as changing TLR and/or FcγR-mediated ROS responses, and modulates oxidative stress and/or innate immune responses.
Materials and methods
CRP
Human plasma CRP (2 mg/ml) was obtained commercially supplied as a solution in 20 mMTris, pH 7.8–8.2, containing 280 mM sodium chloride, 5 mM calcium chloride and 0.1% sodium azide as a preservative (C4063; Sigma-Aldrich Ltd, Gillingham, Dorset, UK). In order to remove azide, CRP (diluted 1:1 in Tris buffer minus azide; 1 mg/ml final concentration) was dialysed against sodium azide-free buffer (20 mMTris, pH 7.8, containing 280 mM sodium chloride, 5 mM calcium chloride; 1 l), as was a corresponding blank control (buffer plus 1 mg/ml sodium azide diluted 1:1 in buffer minus sodium azide). Dialysis was performed over 3 d with three 1-l buffer changes per day at 4℃ with stirring. CRP concentrations post-dialysis were determined by spectrophotometry. All procedures were performed aseptically, and both dialysed CRP and dialysed buffer blank were filter-sterilised (Sigma CLS431212; regenerated cellulose membrane, membrane diameter 4 mm, pore size 0.2 µm) and stored at 2–8℃ for a maximum of 2 wks prior to use.
Heat-aggregated IgG
Freeze-dried IgG purified from human serum (Sigma I4506) was reconstituted with 150 mM NaCl to give a 4.8 mg/ml solution which was heated in a glass tube in a water bath at 63℃ for 60 min, 53 and allowed to cool to room temperature prior to use. Heat-aggregated IgG was prepared immediately prior to use.
Bacterial culture and suspensions
F. nucleatum is a Gram-negative anaerobe present at high frequency and number in the subgingival plaque biofilm in periodontal lesions and is central to periodontal disease pathogenesis as the microflora develops from a non-pathogenic to a pathogenic state.17,54 F. nucleatum [American Type Culture Collection (ATCC) 10953] was grown anaerobically at 37℃, as described previously. 55 Bacteria were isolated from broth cultures by centrifugation, washed three times in sterile PBS and heat treated (80℃ for 10 min) before dilution with sterile PBS to give a final stock suspension of 4 × 108 cells/ml, which was stored at −30℃.
S. aureus [National Collection of Type Cultures (NCTC) 6571] was grown in air on mannitol salt agar and inoculated into tryptone soy broth. Bacteria were washed three times in sterile PBS, and heat-treated (100℃ for 10 min) prior to dilution with sterile PBS to give a final suspension of 5 × 109 cells/ml, which was stored at −80℃. Opsonised S. aureus was prepared as previously described 56 and stored as a 1.2 × 109 cells/ml suspension at −80℃.
Analytical ultracentrifugation data and analyses
Analytical ultracentrifugation was performed using an 8-piece An-60 Ti cell rotor in a Beckman-Optima XL-I using absorbance optics (Beckman Coulter (UK) Ltd, High Wycombe, UK). CRP samples in buffer (20 mM Tris, pH 7.8, containing 280 mM sodium chloride, 5 mM calcium chloride) or IgG samples (heated and unheated) in 0.15 M saline were loaded into two-sector cells for sedimentation velocity experiments and centrifuged at 32,256 g for 8 h at 20℃. The absorbance of the sample was measured at a wavelength of 220 nm throughout the cell. A total of 100 measurements were taken throughout each 8-h run. Sedimentation velocity data were analysed using direct boundary Lamm fits from the continuous c(s) distribution model implemented within SEDFIT (version 11.0). SEDFIT resulted in size-distribution analyses c(s) that assume that all macromolecular species have the same frictional ratio, f/fo.
Liquid chromatography-tandem MS
After reduction and alkylation of disulphide bonds using dithiothreitol and iodoacetamide, CRP was digested overnight (18 h) with trypsin Gold (500 ng; Promega, Madison, WI, USA) at 37℃ and the resultant peptides prepared for analysis as described previously. 57 Peptides were loaded onto a 150-mm Acclaim PepMap100 C18 column (LC Packings, Sunnyvale, CA, USA) in the mobile phase A (0.1% formic acid; JT Baker, Sigma Aldrich, Deventer, Holland) and were separated over a linear gradient from 3.2% to 44% mobile phase B (acetonitrile + 0.1% formic acid; JT Baker, Sigma Aldrich) with a flow rate of 350 nl/min. The column was then washed with 90% mobile phase B before re-equilibrating at 3.2% mobile phase B. The column oven was heated to 35℃. The LC system was coupled to an AdvionTriversaNanomate (Advion, Ithaca, NY, USA), which infused the peptides with a spray voltage of 1.7 kV.
The mass spectrometer performed a full Fourier-transformed MS scan (m/z 380–1600) and subsequent collision induced dissociation (CID) MS/MS scans of the five most abundant ions above a threshold of 1000. Survey scans were acquired in the Orbitrap with a resolution of 60,000 at at m/z 400. Precursor ions were subjected to CID in the linear ion trap. Width of the precursor isolation window was m/z 2 and only multiply-charged precursor ions were subjected to CID. CID was performed with helium gas at a normalised collision energy of 35% (target 5 × 104, maximum fill time 100 ms). CID activation was performed for 10 ms. Dynamic exclusion repeat count was set to 1 with duration of 30 s. Data acquisition was controlled by Xcalibur 2.1 (Fisher Scientific, Loughborough, UK). The purity of CRP was assessed by comparing the intensity of the peptide precursor ions identified from CRP with the peptides which were identified from contaminants.
Neutrophil isolation
Periodontally and systemically healthy volunteers (n = 14) were recruited from the staff of the Birmingham Dental Hospital (10 men, 4 women; mean age = 26 ± 8 yrs, range 21–49 yrs). Ethical approval was obtained from the South Birmingham Local Research Ethics Committee (West Midlands Research Ethics Committee number 10/H/1208/48). Exclusion criteria included a course of non-steroidal anti-inflammatory drugs or antimicrobial drugs within a 3-month period before enrolment, pregnancy or use of vitamin supplements within the previous 3 months.
Following informed consent, venous blood was collected from the ante-cubital fossa into Vacutainer (Greiner, Bio-One, Stonehouse, UK) lithium heparin (17 IU/ml) tubes between 09.00 am and 10.00 am. Neutrophils were isolated from blood as described previously 20,21 using a discontinuous Percoll gradient (δ = 1.079:1:098) followed by erythrocyte lysis (0.83% NH4Cl containing 1% KHCO3, 0.04% Na2EDTA·2H2O and 0.25% BSA). Isolated cells were re-suspended in in PBS supplemented with glucose (1 mM) and cations (1 mM MgCl2, 1.5 mM CaCl2; GPBS) at 1 × 106 cells/ml. Previous studies have demonstrated that both glucose and cations are required for optimal O2- production by neutrophils. 58 Although glucose alone can transiently activate neutrophils, its presence at 1 mM has no effect on neutrophil metabolism and oxidant release. 59 Cell viability, typically >98%, was determined immediately before assay using trypan blue dye exclusion (1:1 mix of cells with trypan blue) and visualisation by light microscopy.
Neutrophil viability in the presence of CRP
The viability of neutrophils treated with commercial CRP (1–6 µg/ml; 37℃; 60 min) was assessed by trypan blue dye exclusion and cell metabolic activity analysed with CellTiter-Glo Luminescent Cell Viability Assay Kit (Promega, Southampton, UK) with a microplate reader (Bertold microplate luminometer LB96v, Berthold, Bad Wildbad, Germany).
Antioxidant capacity of soluble CRP
The ROS scavenging capacity of soluble CRP was determined using the luminol-based enhanced chemiluminescence assay described previously.60,61 The ability of azide-free CRP (final concentrations of 0.2 and 4 µg/ml) to reduce luminol chemiluminescence generated by HRP/H2O2 was determined and compared with vehicle control (azide-free Tris buffer). Data are presented as the percentage reduction in light output

Chemiluminescence assays with neutrophils
Chemiluminescence assays were performed using luminol, to detect mainly intracellular ROS generation (HOCl formed intracellularly by the action of myeloperoxidase on H2O2 generated by dismutation of O2-), and isoluminol and lucigenin to detect extracellular H2O2 (detected as HOCl after the action of HRP within the assay mixture) or O2- release respectively. Cell membranes are permeable to luminol, but impermeable to isoluminol and lucigenin. All assays were performed as described previously.20,21 In brief, neutrophils (1 × 105 in GPBS) were placed in pre-blocked (PBS BSA 1%, overnight at 4℃) white microwells (Microlite 2; Fisher Scientific, Loughborough, UK) with GPBS (30 µl), luminol (3 mmol/l; pH 7.3; 30 µl) and PBS (15 µl) (total volume 75 µl) or isoluminol (3 mmol/l; 60 µl) with 1.5 U HRP in PBS (P8415, 15 µl; Sigma) (total volume 75 µl). The total volume of each well was made up to 165 µl using PBS, and the plate transferred to a microplate reader (37℃; Berthold microplateluminometer, LB96v) and baseline light output was recorded for 1 s per well in relative light units (RLU) for 30 min. CRP from human plasma (pre- and post-dialysis, 10 µl; Sigma C4063) was added (final concentrations: 1, 3 and 10 µg/ml) diluted in dialysed buffer (i.e. 0 µg/ml CRP equates to dialysed buffer) and light output recorded in RLU. After a further 30 min, neutrophils were stimulated with either F. nucleatum [ATCC 10953; multiplicity of infection (MOI) = 100:1; 25 µl], opsonized S. aureus (NCTC 6571; MOI = 300:1; 25 µl), heat-aggregated IgG (25 µl; 600 µg/ml final well concentration; Sigma I4506) or PBS (25 μl) as control to give a total volume of 200 µl per well, and read for a further 120 min. All analyses were performed in triplicate. Peak light emission, in RLU, was determined for the equilibration phase (‘unstimulated’, baseline radical generation), after addition of CRP, and again after FcγR and TLR2/4 stimulation.
Preliminary experiments (n = 5) compared the effect of addition of Tris buffer and PBS on otherwise unstimulated neutrophils. Essentially, Tris buffer and PBS (10 µl) were added in place of CRP after the initial 30-min incubation period and light output monitored for 120 min.
Statistics
Data were recorded automatically in Microsoft Excel, manipulated in Excel and evaluated using GraphPad Prism 5 (version 5.04; GraphPad Software, La Jolla, CA, USA). Non-parametric statistics were used for all tests of statistical significance because the Kolmogorov–Smirnov test demonstrated that not all datasets were normally distributed. Differences between multiple groups were determined by Friedman test followed, where appropriate, by Dunn’s multiple comparisons test to determine differences between control and test groups. The Mann–Whitney test was used to compare the effects of Tris buffer and PBS on neutrophil ROS generation. A level of P ≤ 0.05 was used for assigning statistical significance.
Results
Preparation and characterisation of CRP
Initial studies demonstrated that although the commercial CRP, supplied containing 1 mg/ml sodium azide, had no effect on neutrophil viability, it significantly inhibited ROS generation by neutrophils in a dose-dependent manner. This inhibition could be reproduced using azide alone (data not shown). Therefore, CRP was extensively dialysed to remove azide, filter sterilized, and its molecular mass and purity determined prior to further study. Analytical ultracentrifugation confirmed that the dialysed, filtered protein had a sedimentation coefficient of ∼ 6.2 S and molecular mass of 117.6 kDa, corresponding to pentameric CRP. 62 Liquid chromatography-MS/MS of the two different batches of commercial CRP used in the study confirmed the predominant presence of isoform 1 CRP (>98%) plus 21 micro-contaminant proteins, 11 of which were derived from epidermis or saliva in origin, and commonly present in proteins purified from biological samples. 63
Unstimulated neutrophil ROS generation in the presence of CRP
Addition of CRP (1, 3 and 10 µg/ml) caused significant changes in baseline (unstimulated) ROS generation by neutrophils detected using all chemiluminescent substrates (P ≤ 0.0006) with levels significantly reduced compared with vehicle control (20 mMTris buffer) by 10 µg/ml CRP (P ≤ 0.01; Figure 1). The mean percentage reductions in unstimulated ROS with 10 µg/ml CRP were 41.8%, 11.9% and 28.8% for luminol, isoluminol and lucigenin respectively. Both lucigenin-detected O2- release and luminol-detected intracellular ROS (HOCl) were also significantly reduced by the presence of 3 µg/ml CRP (P ≤ 0.05; Figure 1A, B). Although there were large variations in the responses of cells from individual donors, the effect of CRP in reducing the minimal stimulatory effect of Tris buffer on ROS generation appeared to be dose-dependent.
Unstimulated ROS generation in the presence of CRP as detected by luminol (A), isoluminol (B) and lucigenin (C). Peak chemiluminescence (median, interquartile range, maximum and minimum RLU; n = 14) attained during the 30 min after addition of CRP (1, 3 and 10 µg/ml) or vehicle control (0 µg/ml CRP). Significant differences (Dunn’s test) from vehicle control-treated cells. *P < 0.05; ** P < 0.01; ***P < 0.001.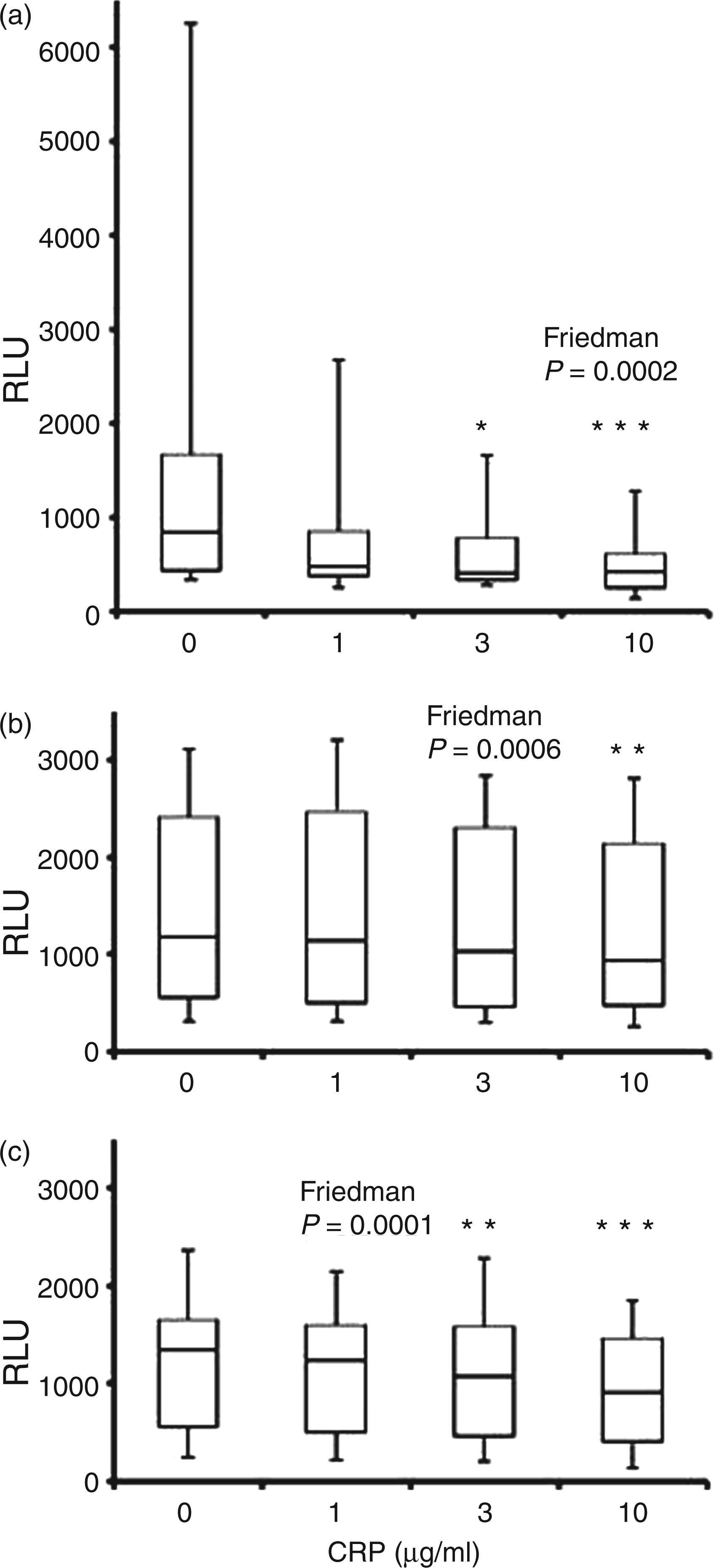
Fcγ and TLR-stimulated neutrophil ROS generation in the presence of CRP
Pre-incubation of neutrophils with CRP prior to, and in its presence during, stimulation with FcγR ligands did not show any obvious relationship between luminol and isoluminol chemiluminescence and concentration of CRP (Figures 2A, C, Figures 3A, C). Although median levels of luminol-detected intracellular HOCl stimulated with IgG-opsonised S. aureus were highest in CRP-treated cells, this increase was only significant for 3 µg/ml CRP and was not seen with aggregated IgG. By contrast, small, but significant, changes in lucigenin-detectable O2− were detected in neutrophils stimulated with both FcγR ligands (P < 0.001; Figure 4A, C), with 10 µg/ml CRP reducing O2− release by 13.9% (P < 0.001) and 17.9% (P < 0.05) in response to IgG-opsonised S. aureus and aggregated IgG respectively.
Luminol-detected neutrophil ROS (detected predominantly as intracellular HOCl) generation following 30 min pre-incubation with CRP and subsequent stimulation with IgG-opsonised S. aureus (A), F. nucleatum (B) and aggregated IgG (C). Peak chemiluminescence (median, interquartile range, maximum and minimum RLU; n = 14) attained after stimulation. Significant differences (Dunn’s test) from vehicle control/0 µg/ml CRP-treated cells. NS: not significant.*P < 0.05. Isoluminol-detected neutrophil ROS (extracellular peroxide, detected predominantly as HOCl) generation following 30 min pre-incubation with CRP and subsequent stimulation with IgG-opsonised S. aureus (A), F. nucleatum (B) and aggregated IgG (C). Peak chemiluminescence (median, interquartile range, maximum and minimum RLU; n = 14) attained after stimulation. Significant differences (Dunn’s test) from vehicle control/0 µg/ml CRP-treated cells. **P < 0.01. Lucigenin-detected neutrophil ROS (extracellular O2−) generation following 30 min pre-incubation with CRP and subsequent stimulation with IgG-opsonised S. aureus (A), F. nucleatum (B) and aggregated IgG (C). Peak chemiluminescence (median, interquartile range, maximum and minimum RLU; n = 14) attained after stimulation. Significant differences (Dunn’s test) from vehicle control/0 µg/ml CRP-treated cells. *P < 0.05; ***P < 0.001.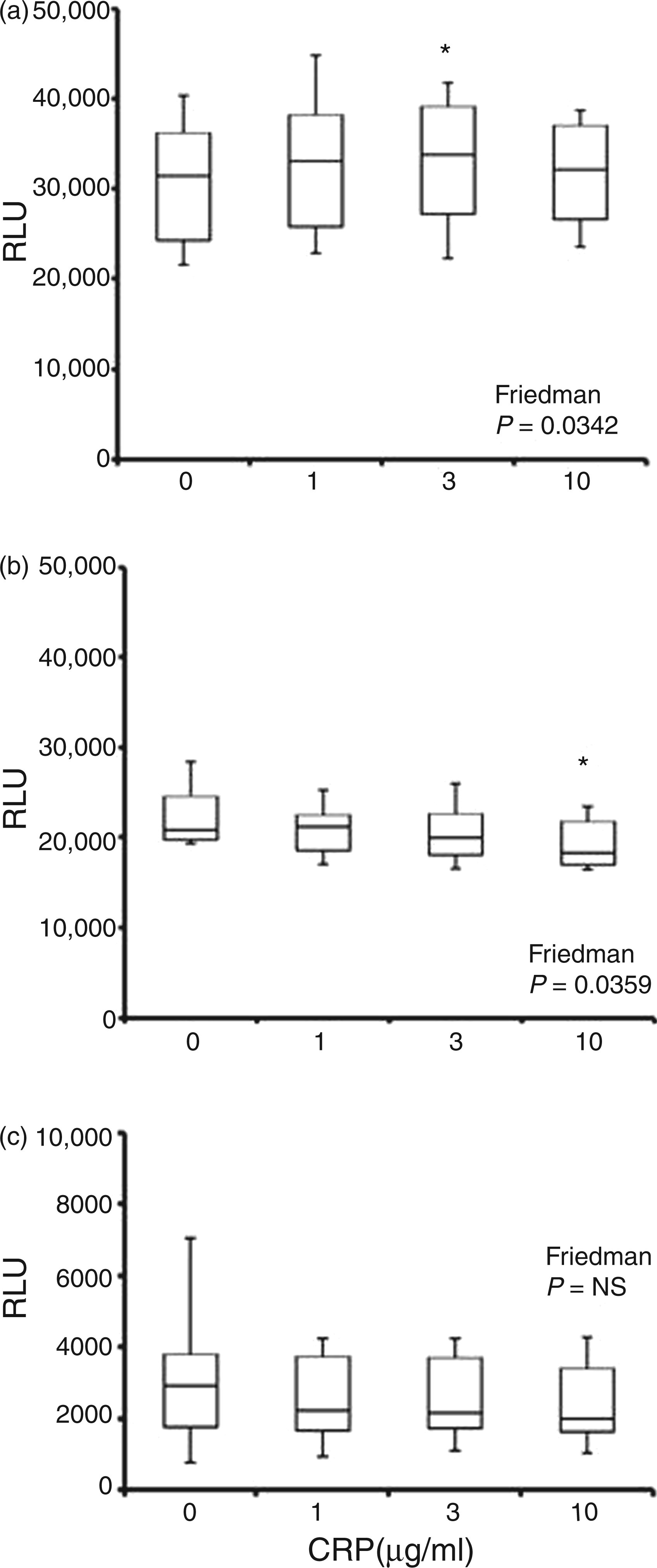
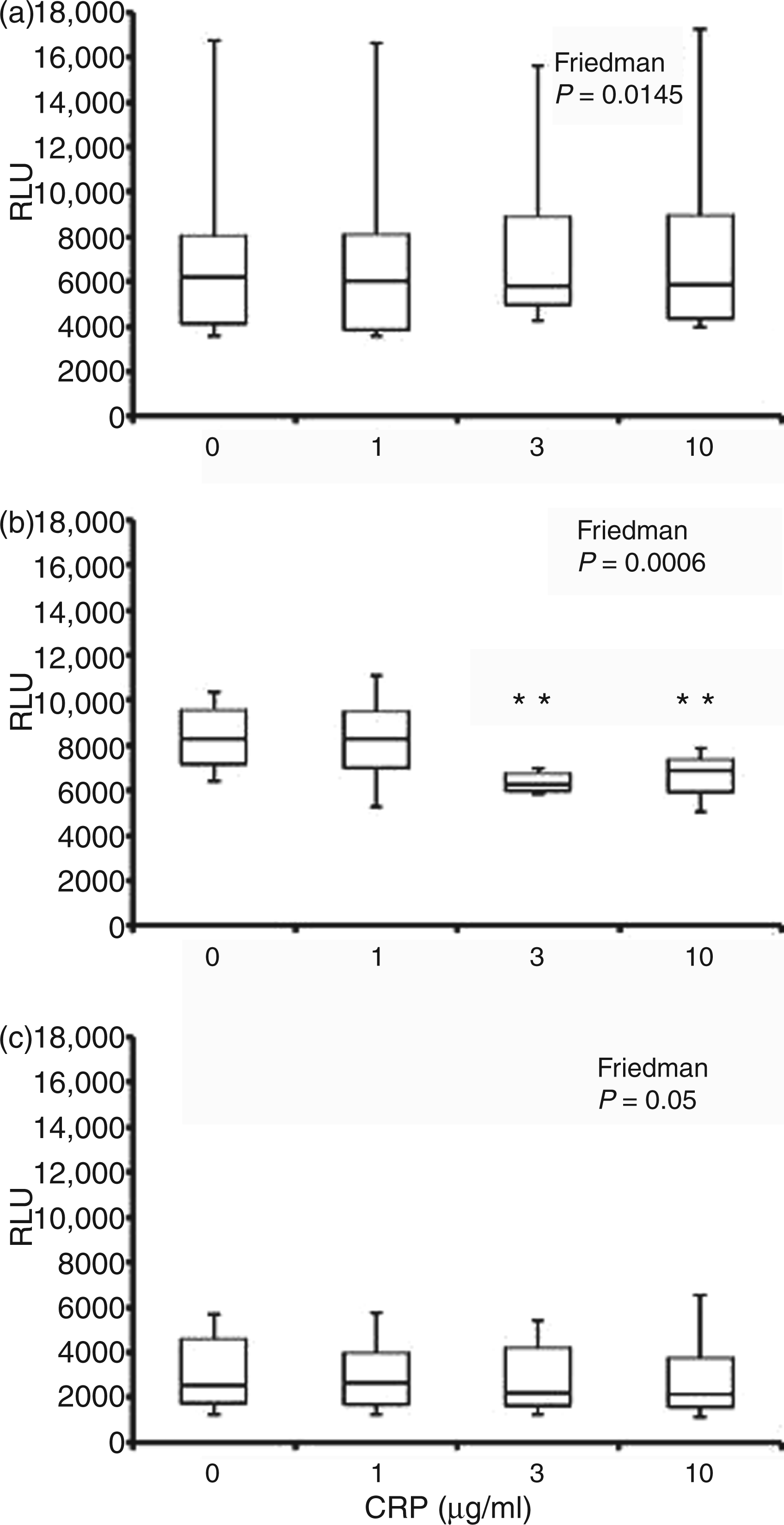
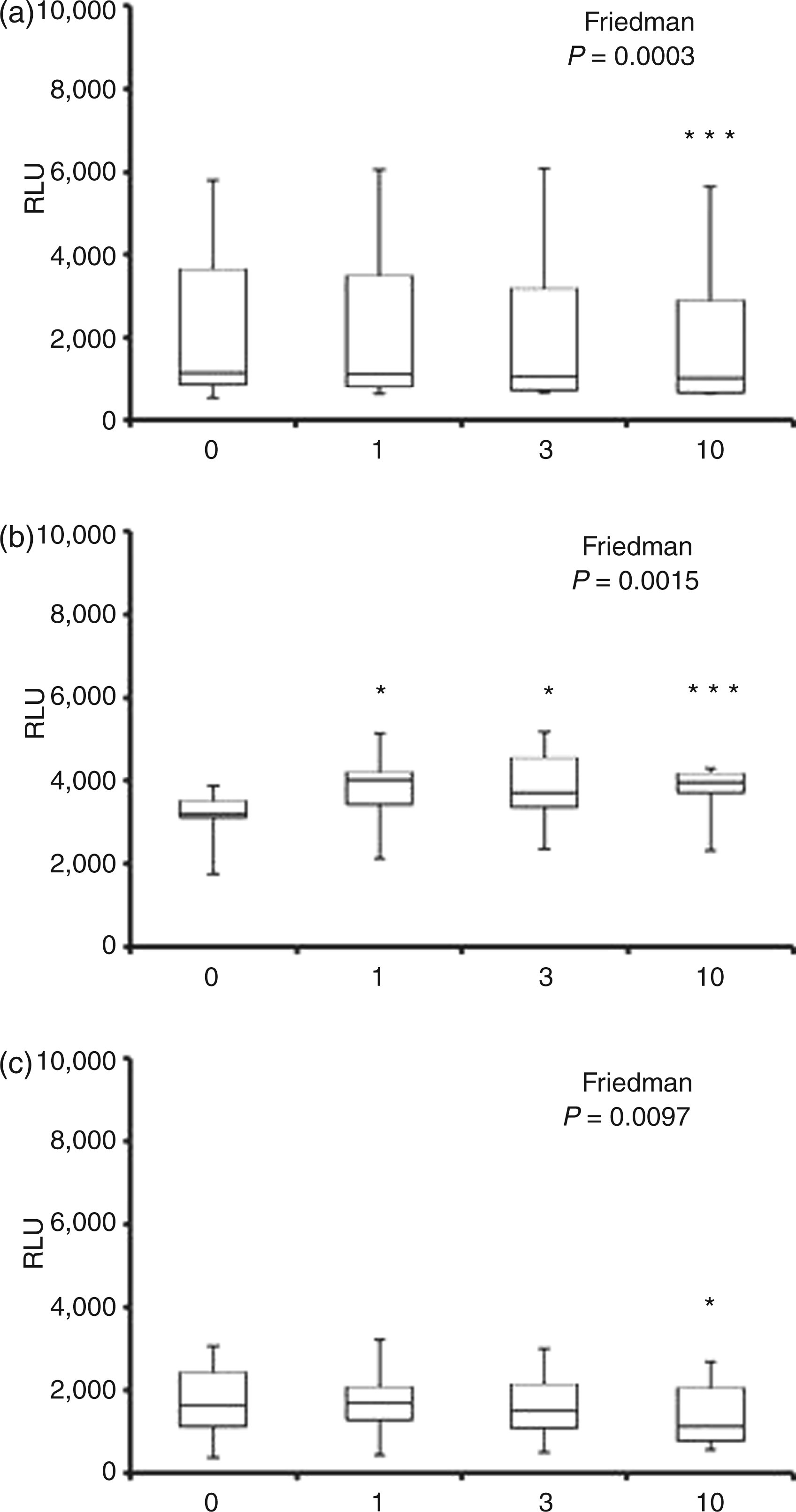
Neutrophil ROS generation in response to stimulation with F. nucleatum was significantly altered by CRP, as measured by all three chemiluminescent substrates (P < 0.04; Figure 2B, Figure 3B and Figure 4B). Although CRP at 3 and 10 µg/ml showed reduced median levels of luminol-detected intracellular HOCl, the change was only significant at the highest CRP concentration (P < 0.05; 13.4% reduction; Figure 2B). The predominant effect of CRP on F. nucleatum-stimulated neutrophil ROS was on extracellular levels (Figure 3B and Figure 4B). CRP decreased extracellular H2O2 release by 22.5% and 19.2% at 3 and 10 µg/ml respectively (P < 0.01). By contrast, lucigenin-detectable extracellular O2− showed a mean increase of 21.5% due to CRP at all concentrations tested (P < 0.05 for 1 and 3 µg/ml; P < 0.001 for 10 µg/ml; Figure 4B).
Antioxidant capacity of CRP
Proteins have some radical scavenging ability and some of the effects of CRP, particularly on ROS generation by unstimulated neutrophils, could have been due to addition of increasing amounts of protein to the test system. The antioxidant capacity of CRP was therefore determined using enhanced luminol chemiluminescence. CRP at 0.2 and 4 µg/ml caused 0.45 ± 2.5% and 0.78 ± 1.08% reduction in light output, respectively, equating to a reduction of the median luminol-dependent RLU for unstimulated neutrophils (844 RLU) by between 3.8 and 6.6 RLU only.
Discussion
This study, for the first time, has demonstrated that low levels of soluble, native, pentameric CRP (≤10 µg/ml) are able to lower baseline unstimulated ROS generation and release by neutrophils. Furthermore, the data show that soluble CRP also down-regulates the neutrophil respiratory burst after FcγR stimulation and, while decreasing intracellular and extracellular ROS as detected by luminol/isoluminol, up-regulates O2− release in response to F. nucleatum.
Chronic periodontitis is characterised by the presence of ‘hyperactive’ peripheral blood neutrophils, which have a greater ability to release potentially tissue damaging ROS in vitro, in the absence of overt stimulation (and CRP), than cells isolated from healthy control individuals.20,21 Furthermore, periodontitis is a common and frequently undiagnosed inflammatory disease, often active across many decades within susceptible individuals, which is associated with a higher prevalence of bacteraemia (including periodontal pathogens) after chewing, brushing, flossing or scaling than detected in gingivitis or periodontal health. 24 The long-term impact of the resulting low-grade systemic inflammation across a patient’s lifespan upon the chronic inflammatory diseases of ageing has a strong evidence base. For example, systemic inflammation precedes type-2 diabetes,64,65 and emerging evidence associates severe periodontitis with incident diabetes. 66 Periodontitis is also a recognised independent risk factor for diabetes complications (e.g. cardio-renal complications) 66 in a dose-dependent manner, and further associates with elevated HbA1c and low-density lipoprotein cholesterol concentrations. 41 It is also independently associated with atherogenic cardiovascular disease 15 and ‘all-cause mortality’. 67 In addition, chronic periodontitis is associated with shorter telomere lengths in peripheral blood leukocytes, with measures correlating to oxidative stress and severity of periodontal disease. 22
Our reported data on unstimulated ROS generation suggest that modest increases in the circulating levels of CRP could reduce baseline ROS generation and release by neutrophils, and have an overall antioxidant/anti-inflammatory effect, especially in chronic, low-grade inflammatory diseases such as periodontitis. Despite the relatively small reductions (12–42%) in baseline ROS production and release demonstrated here, the role of soluble CRP in modulating oxidative stress in vivo may have significant biological relevance. It is possible that the increased circulating CRP levels associated with periodontitis (1–4 µg/ml)40–42 are sufficient to neutralise, or reduce, the low-level, 16–90% excess baseline ROS chronically generated and released by neutrophils in these patients, 20 and thereby reduce both periodontal disease severity and the likelihood of developing another inflammatory disease for which periodontitis is a known risk factor.14–16
The direct antioxidant properties of CRP in our assay system are very low and could not account for the reduction in unstimulated ROS generation detected in the neutrophil assays. Furthermore, the reduced intracellular ROS generation in the presence of CRP could not be due to simple protein scavenging because CRP cannot enter the cell 46 and interfere with the luminol chemiluminescence reaction. This being the case, the reduced, unstimulated ROS generation detected in the presence of CRP is presumably due to a reduction in the activation of NADPH oxidase in the cell membrane and within intracellular granule membranes by CRP-FcγR interaction.
The effects of CRP on ROS generation and release by stimulated cells varied according to stimulus, site and nature of the ROS assayed. Neither intracellular nor extracellular HOCl levels stimulated by FcγR ligands showed a consistent or significant relationship with the presence of CRP whereas O2− release was reduced. That the data for isoluminol-detectable extracellular HOCl showed no relationship with CRP is probably owing to the fact that this ROS product is derived from both cell membrane-released O2- and diffusion of H2O2 from within the cell, with the latter masking the small changes in the former seen with lucigenin. The data on O2−, using pathologically relevant stimuli, are consistent with reports that soluble CRP reduces O2- release after stimulation with PMA, ConA and fMLP44,51,52 and, therefore, potentially acts to moderate neutrophil oxidative responses and reduce oxidative stress. Although it is not possible to know the true biological significance of this reduction (12.5–23.0%) in FcγR-stimulated O2− release, it may suggest a protective effect in moderating extracellular oxidative stress and limiting collateral tissue damage. However, the fact that this reduction was not reflected in reduced intracellular luminol-detected HOCl levels suggests retention of defence functions requiring intracellular ROS (e.g. microbial killing in the phagosome, 68 NET production). 6
As soluble CRP is known to bind FcγRs,45,46 the reduced O2− release after FcγR stimulation could be due to receptor competition. Indeed, competition between IgG and CRP for FcγR binding sites resulting in dose-dependent inhibition of phagocytosis by macrophages has been demonstrated. 45 However, this may not explain our data as similar inhibition of stimulated neutrophil O2− release has been reported with PMA, conA and fMLP, which do not activate cells via FcγRs.
ROS generation and release stimulated by F. nucleatum, and detected as HOCl, was reduced in the presence of CRP, but O2− release was consistently increased. These novel data suggest that stimulation with F. nucleatum in the presence of CRP results in an increased outer cell membrane expression of activated NADPH oxidase, and therefore extracellular release of more O2−, with a concurrent decrease in intracellular membrane expression (e.g. the membranes of secondary and tertiary granules) and intracellular oxidant levels. In normal, unstimulated neutrophils, NADPH oxidase is localized predominantly in intracellular compartments, in particular secondary (i.e. specific)69,70 and tertiary (i.e. gelatinase-containing) granules. 71 When stimulated, NADPH oxidase is translocated to the surface as granules fuse with phagosomal or surface membranes.72–74
Receptor cross-linking is probably the mechanism whereby CRP increases surface expression and activity of NADPH oxidase stimulated by F. nucleatum. The primary binding specificity of CRP in the presence of Ca++ is phosphorylcholine, which is known to be expressed on the cell surface of many bacteria, including F. nucleatum.75,76 It is therefore likely that during stimulation with F. nucleatum in the presence of CRP, cross-linking of TLRs, binding F. nucleatum, with FcγR, binding CRP, would occur and potentially up-regulate the downstream activation events leading to increased translocation of NADPH oxidase to the cell surface and the observed increased release of O2−. Although the importance of receptor cross-linking/receptor cross-talk in lymphocyte activation has been known for a long time, similar interactions between TLR and FcγRs are now being shown to be important in the activation of neutrophils, macrophages and dendritic cells.77,78
This study has focused on the effects of low levels of soluble, pentameric CRP on the neutrophil respiratory burst. However, altered or modified CRP (mCRP) has been detected in a variety of tissues. It is naturally expressed at vascular sites, including sites of neutrophilic inflammation, and is considered to relate to the vascular damage associated with cardiovascular disease. 79 This insoluble form of CRP is antigenically distinct from pentameric CRP and, until recently, could only be produced using heat, acid, or urea dissociation and chelation. 80 However, it has now been shown that mCRP can be generated on cell membranes and activated platelets.33,81 Modified CRP binds neutrophils via FcγRIII (CD16) and inhibits fMLP-mediated chemotaxis. 47 Evidence from a single research group indicates that mCRP has no direct effect on baseline neutrophil ROS generation or ROS release after stimulation but, at high concentrations (>50 µg/ml), appears to increase intracellular ROS responses (1.4-fold) to aggregated IgG, but not PMA or opsonised zymosan.82,83 Pentameric CRP and mCRP exhibit distinct biological activities, and mCRP tends to be pro-inflammatory in nature whereas, consistent with our data, the pentameric form appears to show both anti- and pro-inflammatory properties depending upon the function assayed. 84
In summary, our data indicate, for the first time, that soluble, pentameric CRP, at physiologically relevant concentrations, inhibited baseline neutrophil ROS generation and reduced FcγR-stimulated extracellular O2− release, suggesting that low levels of CRP offer a degree of host protection from background or antibody-mediated oxidative stress. Such an effect could reduce an individual’s susceptibility to periodontitis and reduce the associated systemic chronic inflammation that underlies the risk of developing another inflammatory disease. However, CRP enhanced TLR-mediated O2− release from neutrophils, which could increase oxidative stress, but aid host protection from infection in situations where acquired immunity had not been developed.
Footnotes
Funding
This work was supported by the British Society of Periodontology (grant number 14944); and the Royal College of Surgeons of England (grant number 16209).
Conflict of interest
The authors have no conflicts of interest to declare.