
Abstract
Intragastric growth of Helicobacter pylori and non-Helicobacter microorganisms is thought to be associated with elevated levels of pro-inflammatory cytokines and the production of NO these effects can lead to chronic inflammation. Microorganisms can activate the expression of iNOS and the production of NO by macrophages through stimulation with bacterial LPS. Helicobacter pylori can evade these vigorous immune responses, but the underlying mechanism remains unknown. In this study, we used a murine model of macrophage infection to demonstrate that H. pylori inhibits LPS-induced expression of iNOS and production of NO by macrophages. Suppression of LPS-induced NO production by macrophages led to elevated survival of H. pylori in a trans-well system. This effect was abrogated in macrophages from iNOS–/– mice. Analysis of iNOS mRNA and protein levels revealed that H. pylori inhibits iNOS expression at both transcriptional and post-transcriptional levels, and that these effects occurred with live bacteria. Furthermore, the effect of H. pylori involved down-regulation of the mitogen-activated protein kinase pathway and the translocation of active NF-κB into the nucleus. Taken together, our results reveal a new mechanism by which H. pylori modulates the innate immune responses of the host and maintains a persistent infection within the stomach.
Introduction
Helicobacter pylori is the most common causative agent of gastrointestinal disease in humans. Infection with this pathogen usually occurs in childhood and the bacteria can persist in the stomach for the lifetime of an individual.1,2 Persistent infection with H. pylori in the gastric mucosa induces the expression of NF-κB and the secretion of pro-inflammatory cytokines, including IL-1β, IL-6, IL-8, and TNF-α.3,4 Other inflammatory mediators, such as NO, a bactericidal agent generated by inducible nitric oxide synthase (iNOS) during the conversion of
Nitric oxide is derived from iNOS in LPS-activated macrophages during inflammatory responses. Following treatment of macrophages with LPS, the NF-κB heterodimer rapidly translocates to the nucleus where it activates the transcription of target genes, including iNOS and several pro-inflammatory cytokines. 7 In addition, p38 MAPK, protein kinase C and ERK are also involved in the activation of NF-κB and the expression of iNOS in response to LPS.8,9 Several microorganisms disrupt the activation of MAPKs or the NF-κB signalling pathway in macrophages to evade immune attack.10–14 The effect of H. pylori on the modulation of LPS-activated molecules in macrophages remains unknown.
In addition to H. pylori, non-Helicobacter microorganisms are found in the gastric environment.15,16 One study reported that H. pylori-associated gastritis was associated with the presence of several other microbes in the stomach, including Enterococcus, Pseudomonas, Streptococcus, Staphylococcus and Stomacoccus. 17 A more recent study identified 128 phylotypes in 23 gastric biopsy samples; however, the presence of H. pylori did not affect the composition of microbiota in the gastric microbial community. 18 These findings indicate that H. pylori and non-Helicobacter microorganisms are present in the microbiota of the human stomach, and these microbes can elicit pro-inflammatory mediators and induce vigorous immune responses. 19 These findings also raise the question of how H. pylori persists in the microbial ecosystem under the harsh environment of the stomach.
The aim of the present study was to address thequestion of how H. pylori evades the vigorous antimicrobial activities of macrophages. We established an invitro murine model system and an ex vivo murine model system to examine whether this bacterium could suppress LPS-induced NO production through theMAPK or the NF-κB signalling pathways. We showed that H. pylori inhibits iNOS expression and NO production by murine macrophages stimulated with a high dose of LPS. We further demonstrated that H. pylori down-regulates the LPS-induced activation of phosphorylated p38, ERK1/2 and NF-κB, and it subsequently suppresses LPS-induced macrophage responses. Thus, our study revealed that H. pylori attenuates LPS-induced NO production inmacrophages and consequently evades early host immune responses.
Materials and methods
Antibodies and reagents
Polyclonal rabbit anti-iNOS, anti-phosphorylated c-Jun-N-terminal kinase (p-JNK), and anti-α-tubulin Abs were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Mouse mAbs specific for p38 MAPK, JNK, and p44/42 (ERK1/2) were purchased from Cell Signaling (Beverly, MA, USA). Mouse monoclonal anti-phosphorylated p38 MAPK, and the anti-phosphorylated ERK1/2 (Thr185/Tyr187) Abs were purchased from Upstate (Billerica, MA, USA). LPS (Escherichia coli O55: B5) and aminoguanidine hemisulfate (AG) were purchased from Sigma-Aldrich (St Louis, MO, USA). SB203580 (p38 inhibitor), PD98059 (ERK inhibitor) and SP600125 (JNK inhibitor) were purchased from Calbiochem (San Diego, CA, USA). The activator protein (AP)-1-Luc and NF-κB-Luc plasmids were purchased from Stratagene (San Diego, CA, USA). The iNOS promoter construct (piNOSLuc) was a kind gift from Dr E. A. Ratovitski (Johns Hopkins University, Baltimore, MD, USA). The pSV-β-galactosidase vector and the luciferase assay kit were purchased from Promega (Madison, WI, USA). All other reagents were obtained from Sigma-Aldrich.
Bacterial strains, cell culture and mice
Helicobacter pylori 26695 (ATCC 700392) was used as a reference strain. The cagA or vacA isogenic mutants derived from H. pylori 26695 were constructed as described. 20 Helicobacterpylori strains were recovered from frozen stocks on Brucella agar plates (Becton Dickinson, Franklin Lakes, NJ, USA) containing 10% sheep blood. Helicobacterpylori strains were stored and cultivated as described, 21 and H. pylori extracts were prepared as described. 22 Heat-killed H. pylori was obtained by boiling 1 × 109/ml of bacteria suspended in PBS for 30 min. Crude H. pylori extracts were prepared by sonicating 1 × 109/ml of bacteria suspended in PBS for 5 min on ice. Crude extracts were then centrifuged at 16,000 g for 5 min at 4 °C. The supernatant was filtered through a 0.22 µm filter and used for further analysis.
RAW 264.7 cells (ATCC TIB-71) were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Invitrogen, Carlsbad, CA, USA). De-complement fetal bovine serum (10%; HyClone, Logan, UT, USA) was added to the culture medium. For bacterial infection experiments, the cell culture medium was not supplemented with antibiotics.
Male wild-type C3H/HeN and TLR4-deficient C3H/HeJ mice aged 6–8 wk were kindly provided by Dr Ai-Li Shiau (Departments of Microbiology and Immunology, National Cheng Kung University Medical College, Taiwan). C57BL/6 iNOS knockout (C57BL/6-Nostm1Lau) (iNOS–/–) and wild-type mice aged 6–8 wk were kindly provided by Dr Ming-Chei Maa (Graduate Institute of Basic Medical Science, China Medical University, Taiwan). Mice were maintained at the animal centre of China Medical University (Taichung, Taiwan). All procedures were performed according to the ‘Guide for the Care and Use of Laboratory Animals’ (National Research Council, USA) and were approved by theanimal experiment committee of China Medical University.
Preparation of murine peritoneal exudate macrophages
C57BL/6 iNOS knockout (C57BL/6-Nostm1Lau) (iNOS–/–) and wild-type mice of the same age and gender were used to assess the role of iNOS in H. pylori-induced suppression of LPS-induced NO production by macrophages. Murine peritoneal exudate macrophages (PEMs) were obtained after euthanasia by lavaging each mouse with 10 ml of cold PBS three days after intraperitoneal injection of 2 ml of 3% thioglycolate in PBS. Two h after seeding the cells in culture plates, the non-adherent cells were removed by washing with PBS and the adherent cells were used for further experiments.
Mouse inoculations
C3H/HeN (n = 6) and C3H/HeJ (n = 6) mice aged 6–8 wk were intragastrically inoculated with H. pylori. All mice were maintained under fasting for 24 h before inoculation. The protocol of administration of mouse with LPS was performed as described with slight modifications. 23 Each mouse was administered 1 × 109 CFU/ml of H. pylori and purified LPS (75 µg, phenol extracted from E. coli O55: B5, Sigma-Aldrich) by intragastric gavage for three consecutive days. Six h after the final inoculation with H.pylori, the mice were fed with standard food and water and housed for 1 wk. On the seventh day after infection, six mice in each group were sacrificed and the number of H. pylori in their stomachs was determined by plating on Brucella blood agar plates and expressed as CFU/g tissue.
Immunoblotting
H. pylori-infected cells were washed three times with PBS and boiled in SDS-PAGE sample buffer for 10 min. The samples were then resolved by 10% SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica, MA, USA). The membranes were incubated with primary Abs and then with horseradish peroxidase-conjugated secondary Abs (Invitrogen). The proteins of interest were visualized with ECLTM Western blotting reagents (GE Healthcare, Buckinghamshire, UK) and were detected by exposure to X-ray film (Kodak, Boca Raton, FL, USA).
Reverse transcription and quantitative real time-PCR
Total RNA was extracted from PEMs using TRIzol reagent (Invitrogen), and 1 µg of total RNA was reverse transcribed into cDNA using the oligo(dT) primer. Quantitative real-time PCR using SYBR Green I Master Mix and a model 7900 Sequence Detector System was conducted according to the manufacturer’s instructions (Applied Biosystems, Foster City, CA, USA). After pre-incubation at 50 °C for 2 min and 95 °C for 10 min, PCR was performed with 40 cycles of 95 °C for 10 s and 60 °C for 1 min. The threshold was set above the non-template control background and within the linear phase of target gene amplification in order to calculate the cycle number at which the transcript was detected (denoted as CT). The oligonucleotide primers were: iNOS, forward, 5’-CCCAGAGTTCCAGCTTCTGG-3’, and reverse, 5’-CCAAGCCCCTCACCATTATCT-3’; and GAPDH, forward, 5’-CTCAACTACATGGTCTACATGTTCCA-3’, and reverse, 5’-CTTCCCATTCTCAGCCTTGACT-3’.
Bacterial survival assay
Bacterial survival was assessed in cultures of H.pylori-exposed, LPS-treated macrophages using a trans-well system, as described 22 with slight modification. Briefly, murine PEMs were cultured in the bottom layer of trans-well plates (Corning, Corning, NY, USA). After 48 h, 1 × 106 H. pylori were added to the insert membrane (0.1 µm pore size) and co-incubated for an additional 6 h in culture. The bacteria on the insert membrane were then resuspended and cultured by serial dilution onto Brucella blood agar plates. Colonies were counted after 4 to 5 d of incubation. The CFUs were used to determine anti-bacterial effects.
Determination of NO production and cell viability assay
The NO production was estimated from the accumulation of nitrite (NO2−), a stable end product of NOmetabolism, in the culture medium, using the Griess reagent (Sigma-Aldrich).
24
The MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay was used to measure the effects of LPS and H. pylori on macrophage viability.
25
RAW 264.7 cells or PEMs were infected with various multiplicities of infection (MOI) of H. pylori for 24 or 48 h, respectively. Cell viability was then measured by examining the ability of viable cells to chemically reduce MTT to formazan
Transfection and reporter gene assay
RAW 264.7 cells were grown to 90% confluency in a 12-well plate and transfected with NF-κB-Luc, AP-1-Luc, or iNOS-Luc reporter plasmid using Lipofectamine 2000 (Invitrogen).26,27 After 24 h, cells were incubated without or with LPS and then infected with H. pylori during an additional 24 h culture. To prepare cell lysates, 100 µl of reporter lysis buffer (Promega) was added to each well and cells were scraped from dishes. An equal volume of luciferase substrate was added to all samples and luminescence was measured using a microplate luminometer (Biotek, Winooski, VT, USA). Luciferase activity was normalized to the transfection efficiency as determined by co-transfection of the β-galactosidase expression vector (Promega). 28
Immunofluorescence labeling of phosphorylated p65
To visualize H. pylori-induced inhibition of the translocation of phosphorylated p65 into the nucleus of macrophages, RAW 264.7 cells were seeded onto cover-slips and treated without or with LPS for 2 h and then with H. pylori for an additional incubation at 37 °C for 1 h. Cells were fixed in 3.7% (w/v) paraformaldehyde and permeabilized with 0.5% (v/v) Triton X-100 in PBS for 2 min. For labeling of p65, cells were incubated for 30 min with rabbit polyclonal anti-p65 (H-286; Santa Cruz Biotechnology) and propidium iodide (Calbiochem). Cells were then incubated with a secondary antibody, fluorescein isothiocyanate–conjugated anti-mouse IgG (Chemicon) and they were fixed in paraformaldehyde. Fixed cells were mounted and observed with a confocal laser scanning microscope (Zeiss LSM 510, Carl Zeiss, Göttingen, Germany). The quantification of fluorescence intensity for p65 was analyzed by ZEN software (Carl Zeiss).
Statistical analysis
The Student’s t-test was used to calculate statistical significance; a P value of <0.05 was considered significant.
Results
Helicobacter pylori inhibits LPS-induced NO productionby macrophages
To assess whether H. pylori inhibits LPS-induced NOproduction by macrophages, mouse macrophage RAW 264.7 cells were cultured with LPS (2 µg/ml) and infected with H. pylori at a MOI of 0 to 100 for24 h. NO production, measured by nitrite levels, was not suppressed when LPS-stimulated RAW 264.7cells were infected with H. pylori at a low MOI of 10 (Figure 1A). However, NO production by LPS-stimulated cells was decreased at a MOI of 50–100. When cells were infected with H. pylori ataMOI of 100, LPS-induced NO production wasreduced by approximately 50%. Macrophage viability, as determined by the MTT assay, did not change after 24 h of incubation with LPS and H.pylori (Figure 1B).
Helicobacter pylori–mediated inhibition of LPS-induced NO production in the RAW 264.7 cell line (A) and in murine primary PEMs; (C). Cells were treated without or with LPS (2 µg/ml) and infected with H. pylori at various MOIs from 0 to 100. After 24 h (RAW 264.7 cells) or 48 h (PEMs) incubation, the culture supernatants were collected for determination of nitrite levels using the Griess reagent. The MTT assay showed that there was no loss of cell viability in RAW 264.7 cells (B) or murine PEMs (D) during the incubation period. The data represent the mean ± standard deviation derived from three independent experiments. Statistical significance was determined using the Student’s t-test (*P<0.05; **P<0.01).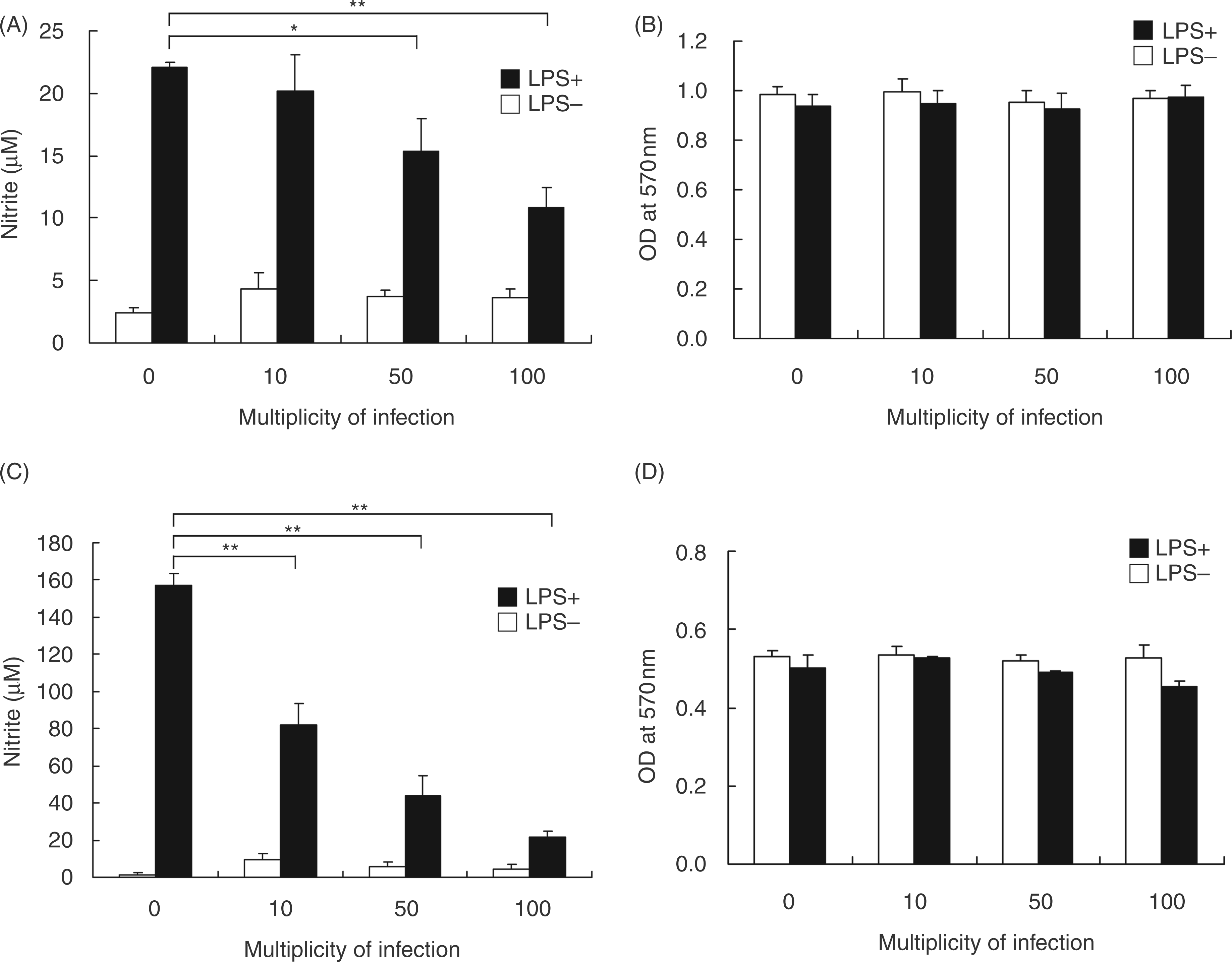
To further delineate the suppressive effects of H.pylori on LPS-induced NO production, PEMs were prepared and co-incubated with LPS and H.pylori for 48 h. This ex vivo approach showed that LPS treatment induced NO production in uninfected PEMs (MOI of 0; Figure 1C), and LPS-induced NO production was reduced in a MOI-dependent manner when PEMs were infected with H. pylori (Figure 1C). The viability of PEMs, as determined using the MTT assay, was hardly influenced by treatment with LPS and H. pylori at different MOIs (Figure 1D). Thus, H. pylori inhibited, in a MOI-dependent manner, LPS-induced NO production, not only in RAW 264.7 cells, but also in murine primary PEMs, and PEMs were more sensitive than the macrophage cell line to the suppressive effects of H. pylori.
Live Helicobacter pylori is essential for the inhibition of LPS-induced NO production
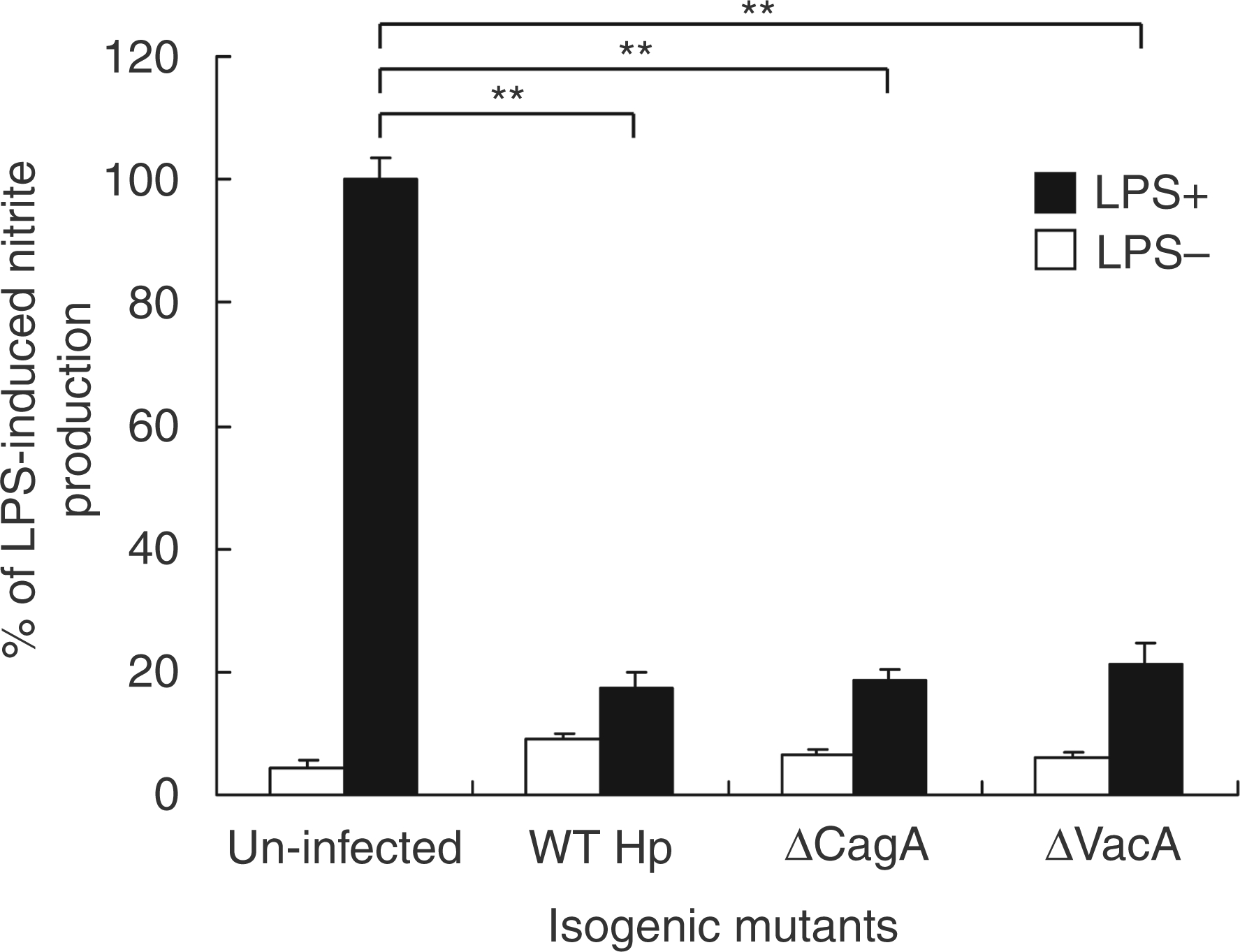
To determine the functional role of H. pylori in inhibiting LPS-induced NO production by RAW 264.7 cells,live bacteria, heat-killed bacteria and crude bacterial extracts were tested for their ability to inhibitLPS-induced NO production. As shown in Figure 2A, live H. pylori attenuated LPS-induced NOproduction by RAW 264.7 cells. In contrast to the effects observed with live bacteria, neither heat-killed bacteria nor crude bacterial extracts inhibited LPS-induced NO production. We also used PEMs to study the effects of H. pylori on the inhibition of LPS-induced NO production. Consistent with the results for RAW 264.7 cells, LPS-induced NO production by PEMs was inhibited by live H. pylori but not by heat-killed bacteria or crude bacterial extracts (Figure 2B). We further investigated the H. pylori-derived components that are responsible for these effects. Virulence factor isogenic mutants of H.pylori, ΔcagA and ΔvacA, were tested for their ability to inhibit LPS-induced NO production by RAW 264.7 cells. Both isogenic mutants suppressed LPS-induced NO production, similar to that observed with wild-type H. pylori (Figure 3).
Live H. pylori are essential for inhibition of LPS-induced NO production by macrophages. Cells were treated without or with LPS and were either infected or un-infected with live H. pylori (live Hp), heat-killed H. pylori (boiled Hp) or crude extracts prepared from H. pylori (Hp lysate). Live and heat-killed H. pylori were used at a multiplicity of infection (MOI) of 100. After RAW 264.7 cells were cultured for 24 h (A) or murine PEMs were cultured for 48 h (B), culture supernatants were collected to measure nitrite levels. Nitrite release from control cells treated with LPS alone was set as 100%. Results are expressed as the mean ± standard deviation derived from three independent experiments. Statistical significance was determined using the Student’s t-test (**P<0.01). Hp, H. pylori. Wild-type and isogenic mutants of H. pylori produce the same inhibitory effect on LPS-induced NO production by macrophages. RAW 264.7 cells were treated without or with LPS and infected with wild-type (WT) or isogenic mutants of H. pylori at a MOI of 100. After incubation for 24 h, culture supernatants were collected to determine levels of nitrite. The data represent the mean ± standard deviation derived from three independent experiments. Statistical significance was determined using the Student’s t-test (** P<0.05). Hp, H. pylori.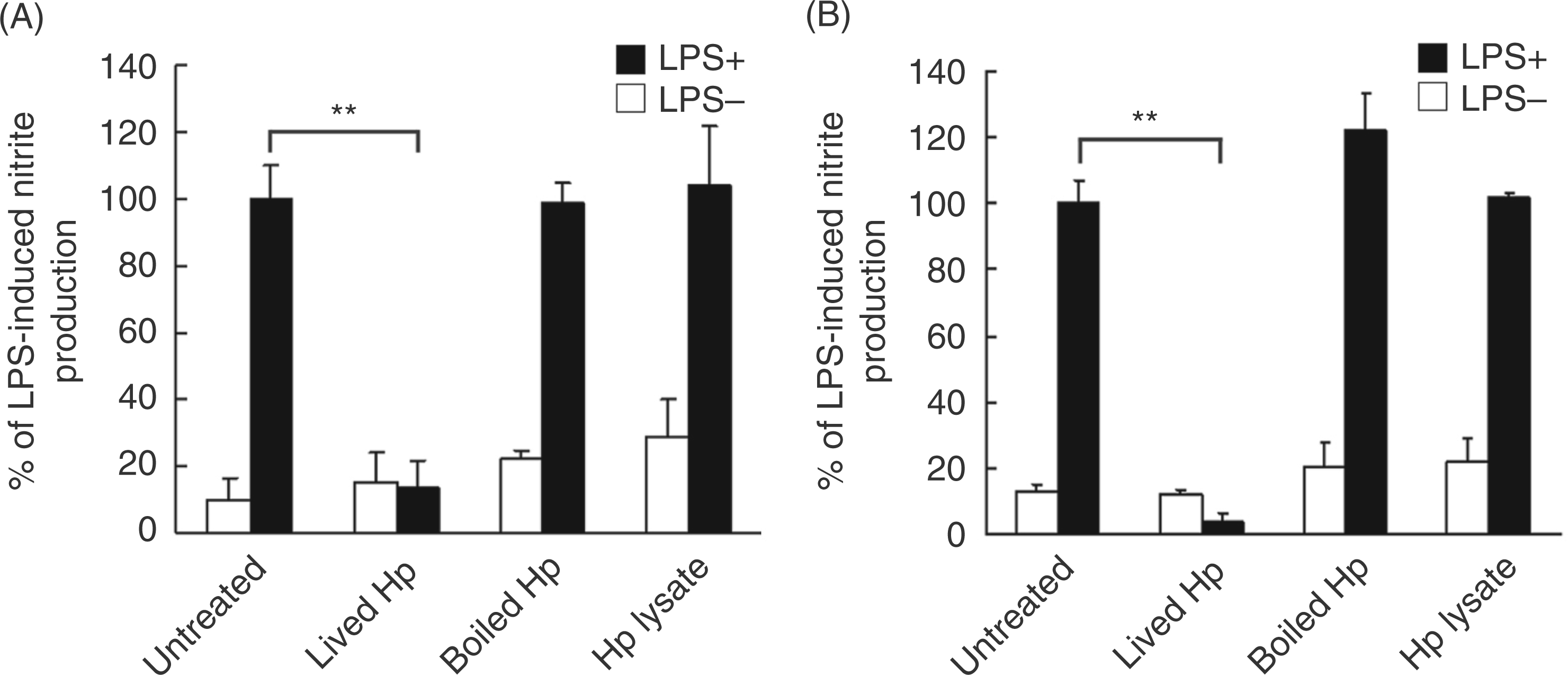
To mimic the environment of the bacterial infection, we further analyzed the ability of H. pylori to inhibit LPS-induced NO production and to enhance the survival of bacteria adjacent to infected macrophages invitro. Using a trans-well culture system to assay bacterial survival of LPS-stimulated murine PEMs, we found that H. pylori at a MOI of 100 increased bacterial viability (Figure 4A). To test the hypothesis that H. pylori triggers iNOS activity, which is necessary for NO production, we tested the effects of H. pylori in C57BL/6 iNOS–/– mice. Helicobacter pylori viability was similar with or without LPS and with H. pylori at a MOI of 0 or 100 in PEMs from iNOS–/– mice, unlike the effects observed in wild-type mice (Figure 4B). These results revealed that the ability of H. pylori to inhibit LPS-induced NO production resulted from a reduction in the antimicrobial activity of macrophages with subsequent enhancement of H. pylori survival.
Bactericidal effects of NO on H. pylori. Murine PEMs from C57BL/6 wild-type (A) or iNOS–/– (B) mice were grown on the bottom layer of trans-well culture plates and infected with H. pylori at a MOI of 0 or 100 for 48 h. The PEMs were then co-incubated with H. pylori in the trans-well insert membrane (0.1 µm) for another 6 h and CFU were counted. Bactericidal activity is expressed as the mean ± standard deviation derived from three independent experiments. Statistical significance was determined using the Student’s t-test (**P<0.01).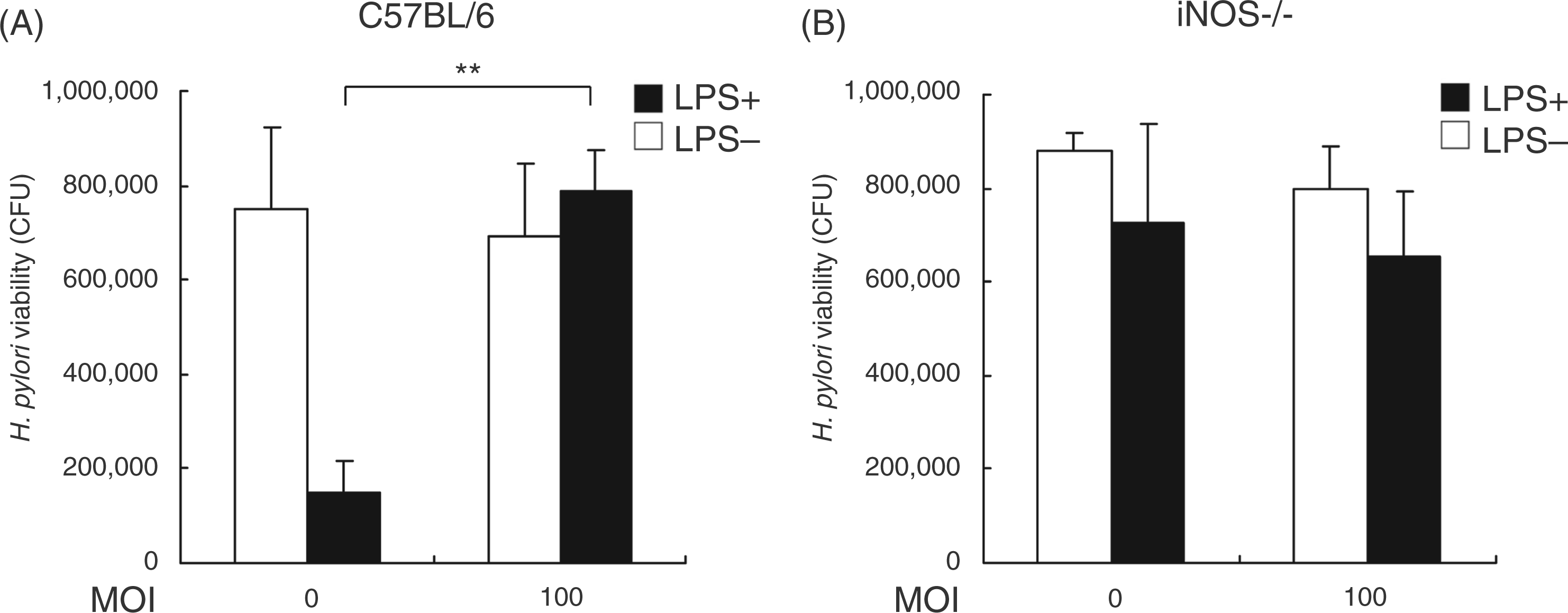
Helicobacter pylori inhibits LPS-induced iNOS expression at transcriptional and post-transcriptional levels
To investigate the effects of H. pylori on the regulation of iNOS expression, PEMs were treated (or left untreated) with LPS and exposed to H. pylori at various MOI for 48 h. Subsequently, iNOS protein levels were analyzed by Western blotting. Helicobacter pylori infection led to a significant decrease, in a MOI-dependent manner, in LPS-induced iNOS level (Figure 5A). When cells were infected with H. pylori at a MOI of 100, LPS-induced iNOS expression decreased by approximately 80%. We then measured expression of iNOS mRNA using reverse transcription and quantitative real-time PCR in PEMs that were co-cultured without or with LPS and with H. pylori atvarious MOI for 6 h. The expression of iNOS mRNA was decreased by H. pylori infection at a MOI of 50to 100 (Figure 5B). Taken together, theresultsindicated that H. pylori inhibited iNOS transcription, which subsequently influenced the translation of iNOS mRNA and NO production by macrophages.
Helicobacter pylori–mediated inhibition of iNOS expression in LPS-treated macrophages. Murine PEMs were treated without or with LPS and infected with H. pylori at different MOIs. (A) Cell lysates were prepared after 48 h of incubation to measure iNOS protein expression by Western blotting. Protein expression levels were quantified with densitometric analysis and normalized to β-actin (B) iNOS mRNA expression was measured by quantitative real-time PCR after 6 h of incubation, and GAPDH was used as an internal control. The data are presented as the mean ± standard deviation of three independent experiments. Statistical significance was determined using the Student’s t-test (*P<0.05; **P<0.01).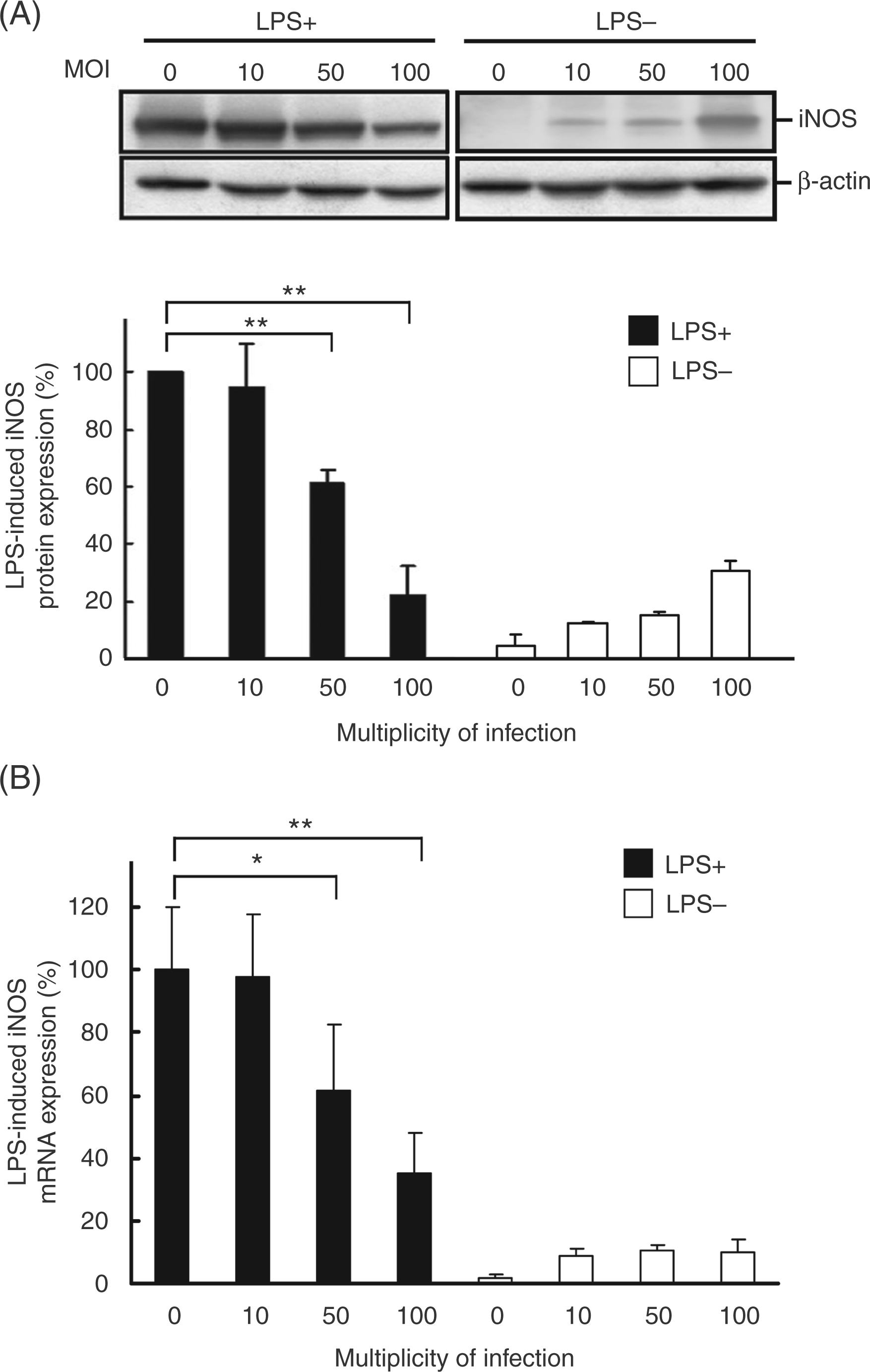
Helicobacter pylori attenuates LPS-induced NO production by macrophages through the p38 and ERK 1/2 signalling pathways
The LPS-induced NO production by macrophages involves several signalling pathways, including p38, ERK 1/2 (p42 and p44), and JNK. We used Western blotting to analyze the signal transduction pathways involved in the inhibitory effects of H. pylori on NO production by LPS-stimulated PEMs for 60 min of culture. The data showed that without treatment with LPS and H. pylori, the phosphorylated molecules involved in the MAPK signalling pathway were expressed at a basal level (Figure 6A). Helicobacter. pylori infection led to a decrease in phosphorylated p38 in LPS-stimulated macrophages within 5 to 10 min of culture (Figure 6B). Phosphorylation of ERK1/2 was also inhibited by H. pylori infection from 5 to 60 min of culture. In contrast to the results obtained for p38 and ERK1/2, phosphorylation of JNK1/2 in LPS-stimulated PEMs was not affected by H. pylori infection. The suppressive effect of H. pylori on LPS-induced iNOS expression was augmented by SB203580 and PD98059, specific inhibitors of p38 and ERK1/2, respectively, but not by SP600125, a specific JNK inhibitor (Figure 6D). Taken together, these results suggested that H. pylori inhibited LPS-stimulated NO production and iNOS expression in macrophages through the p38 and ERK1/2 signalling pathways.
Signalling pathways involved in the inhibition of LPS-induced macrophage activation by H. pylori. Murine PEMs were incubated without LPS (A) or with LPS (B) and infected (or not infected) with H. pylori at a MOI of 100 for the indicated times. Expression levels of phosphorylated p38 (p-p38), anti-phosphorylated extracellular signal-regulated kinases (p-ERK), and anti-phosphorylated c-Jun-N-terminal kinase (p-JNK) were determined by Western blotting. Representative Western blot results from one of three independent experiments are shown. (C) Protein expression levels were quantified with densitometric analysis, normalized to β-actin, and presented as the mean ± standard deviation derived from three independent experiments. Statistical significance was determined using the Student’s t test [*P<0.05; **P<0.01 as compared with LPS-stimulated cells in the absence of H. pylori infection (LPS+, Hp–), lower panel]. (D) Murine PEMs were pretreated for 30 min with SB203580 (p38 inhibitor), PD98059 (ERK inhibitor), or SP600125 (JNK inhibitor), followed by incubation with LPS and infection with H. pylori at a MOI of 100 for 48 h. Representative Western blot results from one of three independent experiments are shown. The data are expressed as the mean ± standard deviation determined from three independent experiments. Statistical significance was determined using the Student’s t-test (**P<0.01). Hp, H. pylori.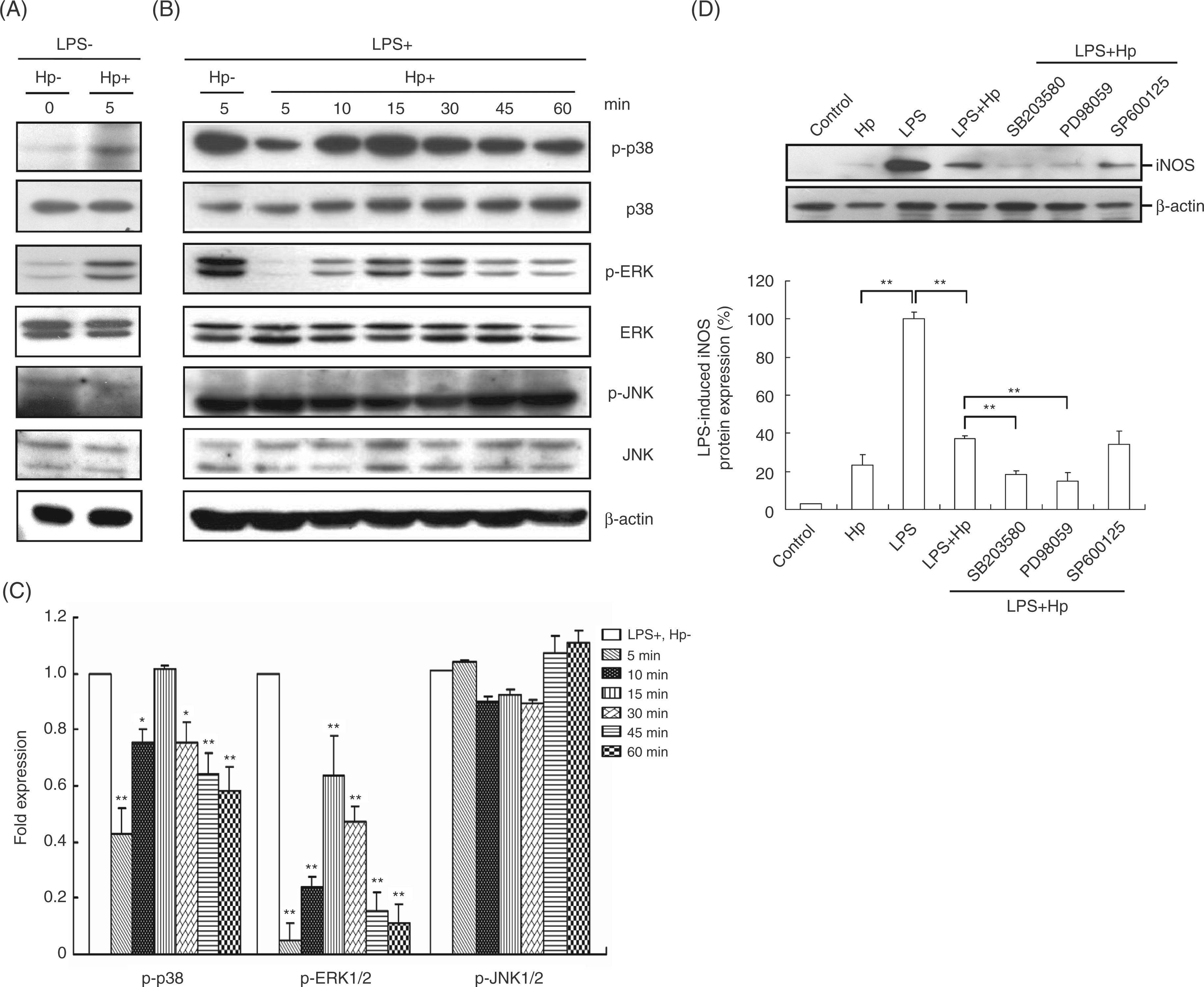
Suppression of LPS-induced NF-κB activation in macrophages by Helicobacter pylori
We next investigated the effects of H. pylori on the levels of two transcription factors, NF-κB and AP-1, which play essential roles in the regulation of iNOS expression.
29
We first examined the effects of Helicobacterpylori on NF-κB expression using the luciferase assay in RAW 264.7 cells transfected with the NF-κB-luciferase reporter. Helicobacter pylori inhibited LPS-stimulated activation of the NF-κB promoter (Figure 7A), but only slightly inhibited LPS-stimulated AP-1 activity; however, the change was not statistically significant (Figure 7B). We next examined p65 localization and observed that it was primarily located in the cytosol before LPS treatment. After 1 h of stimulation with LPS, p65 translocated into the nucleus of RAW 264.7 cells. When cells were co-cultured with LPS and H. pylori, however, p65 largely remained in the cytosol (Figure 7C). The quantitative data showed that the inhibition of p65 translocation into the nucleus in LPS-treated macrophages upon H. pylori infection (Figure 7D). These data suggested that H. pylori alone did not alter the distribution of p65 but rather prevented LPS-induced translocation of p65 into the nucleus.
Helicobacter pylori–mediated attenuation of LPS-induced NF-κB activation. RAW 264.7 cells transfected with a reporter gene for NF-κB (A), AP-1 (B), or β-Gal-lacZ (1 µg each) were treated without or with LPS and infected (or not infected) with H. pylori at a MOI of 100. Luciferase activity was normalized to the expression level of β-Gal-lacZ. The data are expressed as the mean ± standard deviation derived from three independent experiments. Statistical significance was determined using the Student’s t-test (**P<0.05). (C) The nuclear translocation of p65 after 1 h treatment with or without LPS and infection of H. pylori at a MOI of 100. Cells were washed and treated with anti-p65 followed by fluorescein isothiocyanate–conjugated anti-mouse IgG (green). Cells were co-stained with propidium iodide to visualize the nucleus (red). Cells were analyzed by confocal fluorescence microscopy. Regions of p65 nuclear localization appear in yellow in the overlay. Representative images from one of three independent experiments are shown. Scale bar, 10 µm. Ctrl, control; Hp, H. pylori.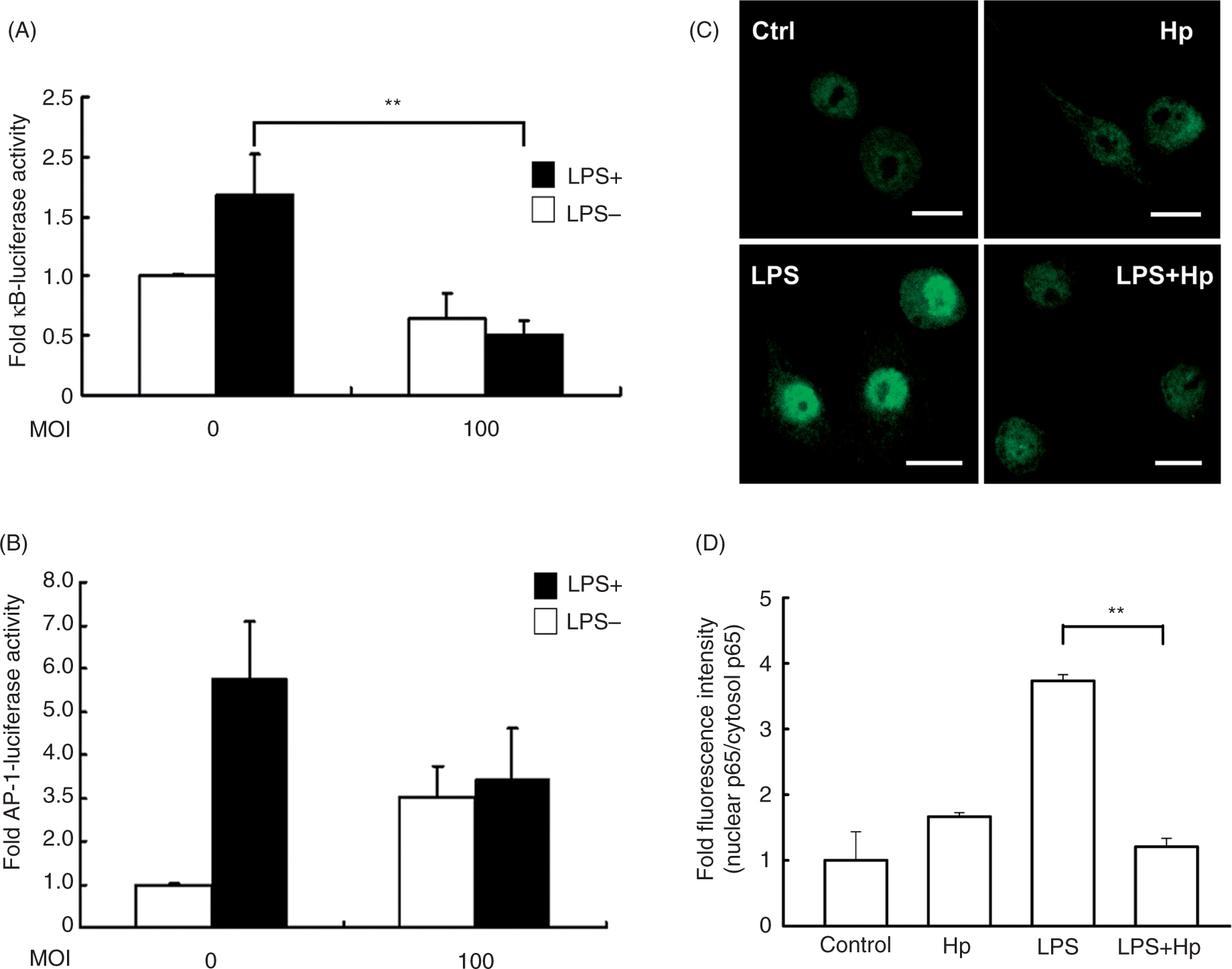
Discussion
Inducible iNOS and NO are two well known factors that play important roles in the antimicrobial response of macrophages. 30 Our results showed that infection of H.pylori elicited a small amount of NO production by macrophages (Figure 1). Live H. pylori and also heat-killed H. pylori and bacterial lysate slightly stimulated NO synthesis in the absence of bacterial LPS (Figure 2). In addition, without LPS treatment, H.pylori stimulated, in a MOI-dependent manner, the expression of iNOS mRNA and protein (Figure 5). Our results confirm the results of a previous study that showed that H. pylori can stimulate iNOS expression and activity in murine macrophages. 5 The release of NO appears to be stimulated by H. pylori-derived components because even heat-killed H. pylori can stimulate NO production. 31 Lipopolysaccharide derived from H.pylori produces only low biological activity as a stimulator of NO, 32 with estimates that it is 2000–30,000-fold less potent than LPS derived from E. coli. 33 Wilson et al. suggested that H. pylori induces iNOS expression through both LPS-dependent and -independent mechanisms. 5 The expression of iNOS and the accumulation of NO have been linked to H. pylori-associated gastritis.6,34 These results demonstrate that NO production plays an important role in gastric inflammatory responses elicited by H. pylori.
Apart from infection with H. pylori, the stomach is colonized with non-Helicobacter microorganisms. The most common non-Helicobacter bacteria of the gastric microflora are Streptococcus, Staphylococcus, Neisseria, Pseudomonas and Enterobacteriaceae.
15
In addition, endoscope-transmitted infections have been reported, including Salmonella spp., E. coli, Klebsiella pneumoniae and Pseudomonas aeruginosa.
35
Long-term infection with H. pylori
15
or acid-suppressive therapy
16
enables non-Helicobacter bacteria to colonize the host stomach. A diverse community of 128 phylotypes was identified using large-scale 16S rRNA sequencing in 23 gastric endoscopic biopsy samples, suggesting that the human stomach may be host to a distinct microbial ecosystem.
18
Both LPS and peptidoglycan (PGN), which is found in the cell wall of Gram-negative and Gram-positive bacteria, have been implicated in the production of NO and pro-inflammatory cytokines by macrophages.36–39 Patients with both H. pylori and non-Helicobacter bacteria in their gastric mucosa have higher levels of pro-inflammatory cytokines than patients without bacterial infection.
19
The issue that these findings raise, therefore, is how H. pylori can survive in a hostile gastric environment surrounded with profound inflammatory responses provoked by other microorganisms. To mimic the hostile gastric environment with activated macrophages and co-infection of H. pylori and non-Helicobacter bacteria, purified LPS or PGN were added to the macrophage culture system to determine the effects on NO production. In a preliminary study, we found that treatment with PGN (10 µg/ml) for 24 h resulted in an approximately four-fold increase in nitrite (8 µ
In the present study, we used in vitro and ex vivo murine model systems to reveal how H. pylori evades LPS-dependent killing by macrophages. To demonstrate that the rational design of our murine model systems mimic a real-life in vivo situation, we used wild-type C3H/HeN and TLR4-deficient C3H/HeJ mice to study the role of LPS in killing gastric H.pylori in vivo. As shown in Supplementary Figure 2, H. pylori was more significantly eradicated from the stomachs of LPS-administered C3H/HeN mice than from the stomachs of LPS-administered C3H/HeJ mice. The response of LPS-administered C3H/HeN mice was approximately 11-fold greater than that of LPS-administered C3H/HeJ mice. This result confirmed that LPS enhanced in vivo anti-H. pylori activity in LPS-responsive mice (C3H/HeN). In addition, the results of this experiment proved our assumption from the in vitro and ex vivo murine models as the responses were mimicked in the in vivo setting.
In this study, we demonstrated that LPS-induced NO production by macrophages was suppressed by H. pylori and that this effect was dependent on the presence of live bacteria (Figure 2). Neither heat-killed bacteria nor crude extracts of H. pylori suppressed LPS-induced NO production. Likewise, the work of von Bothmer et al. demonstrated that H. pylori water extract and whole-bacterial suspension produced an
Several reports have shown that NO can kill H.pylori in cell culture systems.22,41 Our data indicated that H. pylori could survive when H. pylori was co-cultured with LPS-activated macrophages in a trans-well system (Figure 4). This effect may be as a result of the suppressive effects of H. pylori on LPS-induced NO production by macrophages. This possibility is consistent with the results of previous studies that demonstrated that arginase produced by H. pylori
22
and arginase II released from macrophages
42
suppress NO production and lead to immune evasion by the bacteria. Another explanation for the mechanisms through which H. pylori inhibits NO production is a decrease in the concentration of
Lipopolysaccharide stimulates iNOS gene expression and NO production. Stimulation is positively regulated by NF-κB, which is normally bound to its inhibitor, IκB, in the cytoplasm. Phosphorylation of IκB by IκB kinase results in the degradation of IκB, which dissociates NF-κB and leads to the nuclear translocation of NF-κB and the up-regulation of downstream gene expression. In addition to NF-κB, LPS can activate MAPK pathways in macrophages, including p38, ERK-1/2, and JNK-1/2. A common strategy for pathogens to overcome host defense is interference with the activity of NF-κB or MAPKs.10,12,13,46 In the present study, H. pylori may have suppressed LPS-induced NO production by macrophages by inhibiting LPS-stimulated NF-κB activation. Our data demonstrated that the phosphorylation of p38 and ERK1/2 was attenuated by H. pylori infection at an early stage of infection, whereas JNK-1/2 was not affected. These results suggest that H. pylori targets p38 and ERK 1/2, but not JNK1/2. Our data are consistent with previous findings that pathogens can exploit NF-κB to manipulate cellular responses.10,13,14,47 Together, these findings support our hypothesis that H. pylori modulates hostsignalling to evade the host immune system. Inaddition, our results provide insight into the molecular mechanisms through which indigenous H.pylori survive commensurately in the stomach with non-Helicobacter microorganisms that induce potent immune responses.
In conclusion, we demonstrate that infection with live H. pylori attenuates LPS-induced iNOS gene transcription and NO production in a mouse macrophage model. We further find that H. pylori inhibits LPS-induced MAPK signalling and NF-κB activation in macrophages. Collectively, this study reveals a new mechanism through which H. pylori modulates host cell signalling to protect itself from inflammatory responses and to survive in the harsh environment of the stomach.
Footnotes
Acknowledgements
This work was supported by the National Science Council (NSC 97-2313-B-039-003-MY3); China Medical University (CMU98-S-09 and CMU99-S-09); and the Tomorrow Medicine Foundation. The authors thank Dr Wen-Ching Wang (National Tsing-Hua University) for valuable suggestions and comments on this work. We thank Dr Ai-Li Shiau (National Cheng Kung University Medical College) for providing the C3H/HeN and C3H/HeJ mice.
Conflicts of interest
None declared.