
Abstract
Avian influenza virus H5N1 is a potentially fatal disease not only in birds, but also in humans. The virus is able to induce apoptosis in many cell types including macrophages and dendritic cells. In the present study, we demonstrated that TNF-related apoptosis-inducing ligand (TRAIL) is involved in apoptosis-associated mechanisms of apoptosis downstream of the TRAIL receptor in H5N1 virus-infected human monocyte-derived macrophages (MDMs). Activation of caspase-10 was also observed in avian virus H5N1-infected MDMs. In the presence of caspase-10 inhibitor, Z-AEVD-FMK, the activation of Bid and a release of apoptotic-inducing factor (AIF) from mitochondria were markedly reduced, resulting in a significant decrease of apoptotic cells which suggested the involvement of caspase-10 activation in mitochondria leakage. Furthermore, neutralizing Ab against TRAIL significantly reduced caspase-10 activities, which paralleled with a decrease in the number of apoptotic cells. Together, this study demonstrated that apoptosis in avian virus H5N1-infected MDMs was induced by TRAIL-activated caspase-10, resulting in the activation of Bid and the release of AIF from mitochondria.
Keywords
Introduction
Since the re-emergence of H5N1 virus in 2003, a total of 552 confirmed cases of human H5N1 infection have been reported to World Health Organization (WHO) and the mortality rate of these patients is greater than 50%. 1 Clinical manifestations include lymphopaenia and severe pneumonia, which subsequently progresses to acute respiratory distress syndrome (ARDS). Thisprocess links with diffuse alveolar damage from desquamation of alveolar cells and haemorrhage. 2 – 4 Unlike most other influenza infections which are limited to the respiratory system, the H5N1 virus could spread beyond the primary site of infection in the lung causing blood-borne systemic infection.5,6 High levels of proinflammatory cytokines were demonstrated in primary human macrophages infected withavian H5N1. 6 This cytokine dysregulation may contribute to severity of the disease. Furthermore, the presence of lymphoid depletion, leukopaenia and lymphopaenia in H5N1-infected patients suggest that the H5N1 virus can evade host immunity by killing these immune cells.2,3,5 Several lines of evidence showthat the H5N1 virus is capable of inducing cell death in various cell types including lymphocytes, epithelial cells, dendritic cells and monocytes.4,7– 9 The ability of H5N1 virus to induce apoptosis in human cells may contribute to the pathogenesis of this infectious disease.
In general, apoptosis can be initiated by two different pathways: the intrinsic (or mitochondrial pathway) and the extrinsic pathway. The mitochondrial apoptotic pathway is regulated by proteins in the Bcl-2 family, such as Bax and Bid. 10 Following activation, these proteins trigger the release of intermembrane space proteins, for example cytochrome c, which facilitate caspase-9 activation. 11 Other molecules, such as apoptotic-inducing factor (AIF) and endonuclease G are also released from the mitochondria to the cytosol before translocating to the nucleus to induce chromatin condensation and DNA fragmentation. In contrast, the extrinsic pathway is initiated by interaction between death ligands, such as TNF-α, FasL (CD95L) and TNF-related apoptosis-inducing ligand (TRAIL/APO-2L) and corresponding receptors, thus resulting in recruitment of Fas-associated death domain containing protein (FADD) and caspase-8 to the death-inducing signalling complex (DISC). 11 – 13 Activation of caspase-8 then initiates the apoptosis-executing caspase cascade. In contrast, the information regarding the significance of caspase-10, an enzyme structurally closely related to caspase-8, in apoptosis induction is limited. Death ligands such as TRAIL and FasL can initiate apoptosis by the recruitment of FADD and caspase-10 to DISC, leading to activation of caspase-3.14,15 Caspase-10 could function independently of caspase-8 in initiating FasL and TRAIL. 16
TRAIL has been implicated in the induction of apoptosis in avian H5N1 virus-infected human monocytes-derived macrophages (MDMs). 17 Significant upregulation of TRAIL and TNF-α, but not FasL, was observed in avian H5N1 virus-infected MDMs. The induction of apoptosis was reduced by TRAIL Ab suggesting that TRAIL participates in apoptosis induction and contributes to disease pathogenesis. 17 Information regarding the molecular mechanisms of apoptosis induced by avian H5N1 in human MDMs is limited. It was demonstrated that this virus was able to activate caspase-3 and caspase-8 with a delayed onset. 18 Activation of these caspases also resulted in degradation of poly (ADP-ribose) polymerase (PARP). As caspase-10 is known to initiate apoptosis through TRAIL, we therefore analyzed the involvement of this enzyme in apoptosis induction of avian H5N1 in human MDMs.
Materials and methods
Virus isolation
An open-billed stork/Nakhonsawan/BBD0104F/04 strain of H5N1 used in the present study was initially isolated from cloacae swabs of Asian open-billed storks in 2004 and propagated in Madin-Darby canine kidney cells (MDCK). 8
Cell culture and viral infection
The MDMs were prepared from heparinized blood collected from healthy volunteers. Peripheral blood mononuclear cells were prepared by density gradient centrifugation using Unmarked set by EA37 Ficoll-Paque (Lymphoprep™, Axis-Shield PoC AS, Oslo, Norway) and purified by CD14+ MACS magnetic beads (MACS®, Miltenyi Biotech GmbH, Bergisch Gladbach, Germany) as described previously. 19 Differentiated macrophages were infected in the presence or absence of monoclonal anti-TRAIL (Abcam, Cambridge, UK), at a multiplicity of infection (MOI) of1. At times indicated, depending on experimental design, the cells were collected for analysis.
TUNEL staining
Apoptotic cells were examined after fixation of the cells for at least 30 min with 4% paraformaldehyde in PBS.The cells were stained with TUNEL reaction mixture (Roche Diagnostics, Mannheim, Germany) for 60 min at 37°C, washed twice with PBS and incubated with alkaline phosphatase–conjugated fluorescein isothiocyanate–labeled Ab for 30 min at 37°C. The apoptotic cells were observed under a fluorescence microscope and the percentage of apoptotic cells was determined by counting at least 500 cells per sample.
lmmunoblotting analysis
Human MDMs infected with H5N1were treated with lysis buffer [0.0625
Caspase-10 activity assay
A colorimetric assay for caspase-10 activity (Caspase-10 Colorimetric Protease Assay Kit, BioVision Inc., Mountain View, CA, USA) was used according to the manufacturer’s instructions.
Measurement of TRAIL production
Production of TRAIL was measured by ELISA (eBioscience, Vienna, Austria).
Statistical analysis
Statistical comparisons were analyzed by one-way ANOVA test. P-Values of <0.05 were considered statistically significant.
Results
H5N1 induces apoptosis in human MDMs via activation of caspase-10
Avian influenza H5N1 virus has been previously shown to infect human MDMs.
17
Under the conditions used here, viral replication, as detected by real time PCR, exponentially increased until 21 h after the infection (data not shown). To determine whether H5N1 induces cell death in human MDMs, the cells were infected with H5N1 at MOIs of 1 for 12, 18, 21 and 24 h before the number of apoptotic cells were analyzed TUNEL staining. The results illustrated in Figure 1 showed that levels of apoptosis were found to be 5%, 26%, 39% and 52% respectively. These results demonstrate that H5N1 could induce cell death by apoptosis in human macrophages.
Apoptosis of avian H5N1 virus-infected human MDMs. Percentage of apoptotic infected human MDMs after 24 h of infection was determined by TUNEL staining. Data are the mean ± SE from 3 independent experiments, *P<0.05.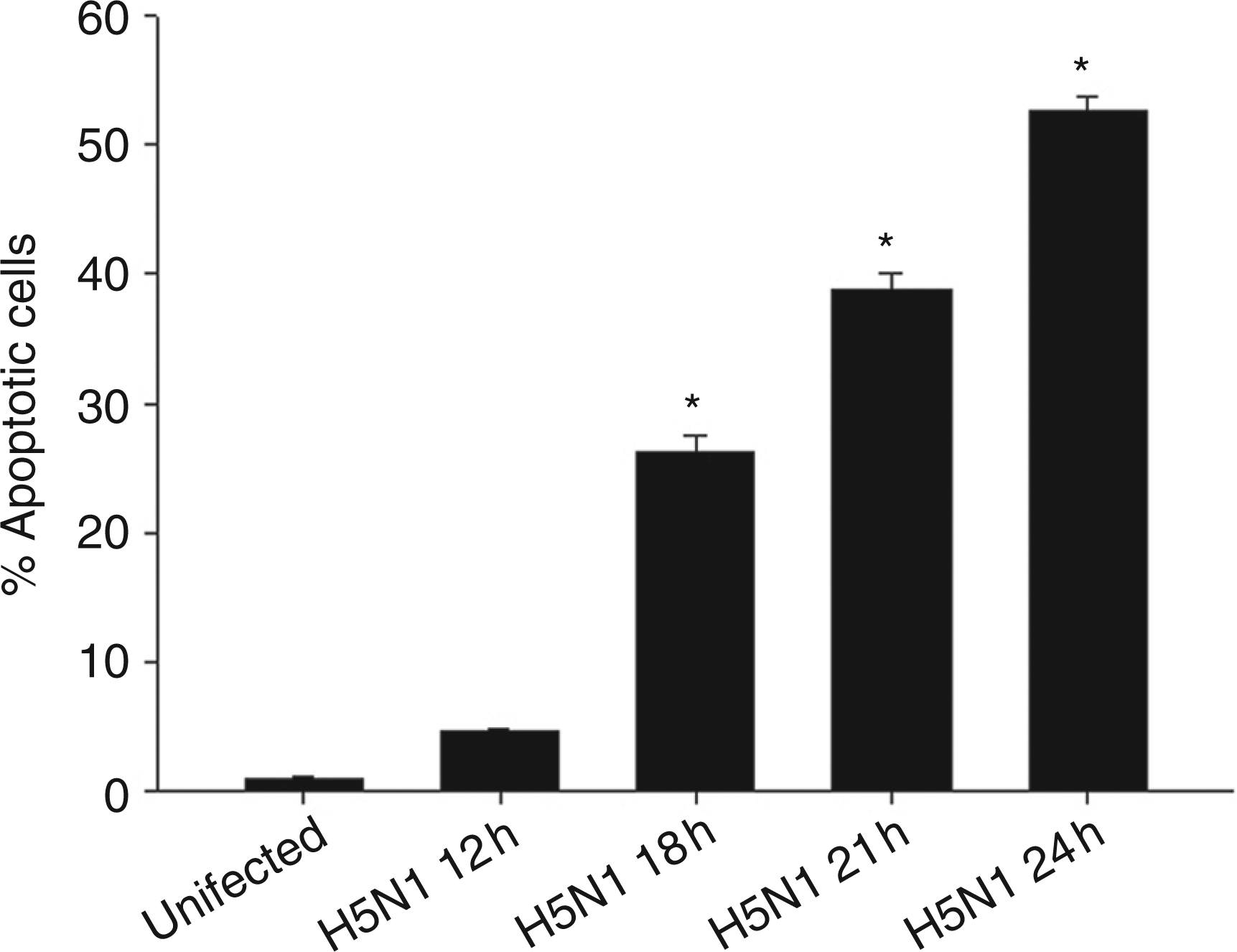
We further performed experiments to determine whether caspase-10 activity was involved in infected cells. Results presented in Figure 2A showed that caspase-10 activity in H5N1-infected cells (MOI of 1) increased more than 2-fold after 18 h of infection, reaching maximum activity by 21 h, suggesting that caspase-10 was activated in H5N1-infected MDMs. This result was also consistent with the degradation of procaspase-10 shown in Figure 2B. It should be noted that in the infected cells, procaspase-7 was also degraded, thus suggesting that H5N1 also induced caspase-7 activation in human MDMs. In order to demonstrate that activation of caspase-10 is involved in apoptosis in our model, the number of apoptotic cells induced by H5N1 was determined in the presence of caspase-10 inhibitor, Z-AEVD-FMK. Data presented in Figure 2C showed that enzyme activity in these virus-infected cells was completely abolished by Z-AEVD-FMK at a concentration of 10 μ Activation of caspase-10 in avian H5N1 virus-infected human MDMs. Human MDMs were infected with H5N1 at multiplicity of infection (MOI) of 1 for 18, 21 and 24 h. Caspase-10 activity was determined by colorimetric assay and expressed as fold increase relative to that of uninfected cells (A). Immunoblotting analysis was performed to detect procaspase-10 and -7. Actin was used as loading control (B). Human MDMs were treated with 10 µ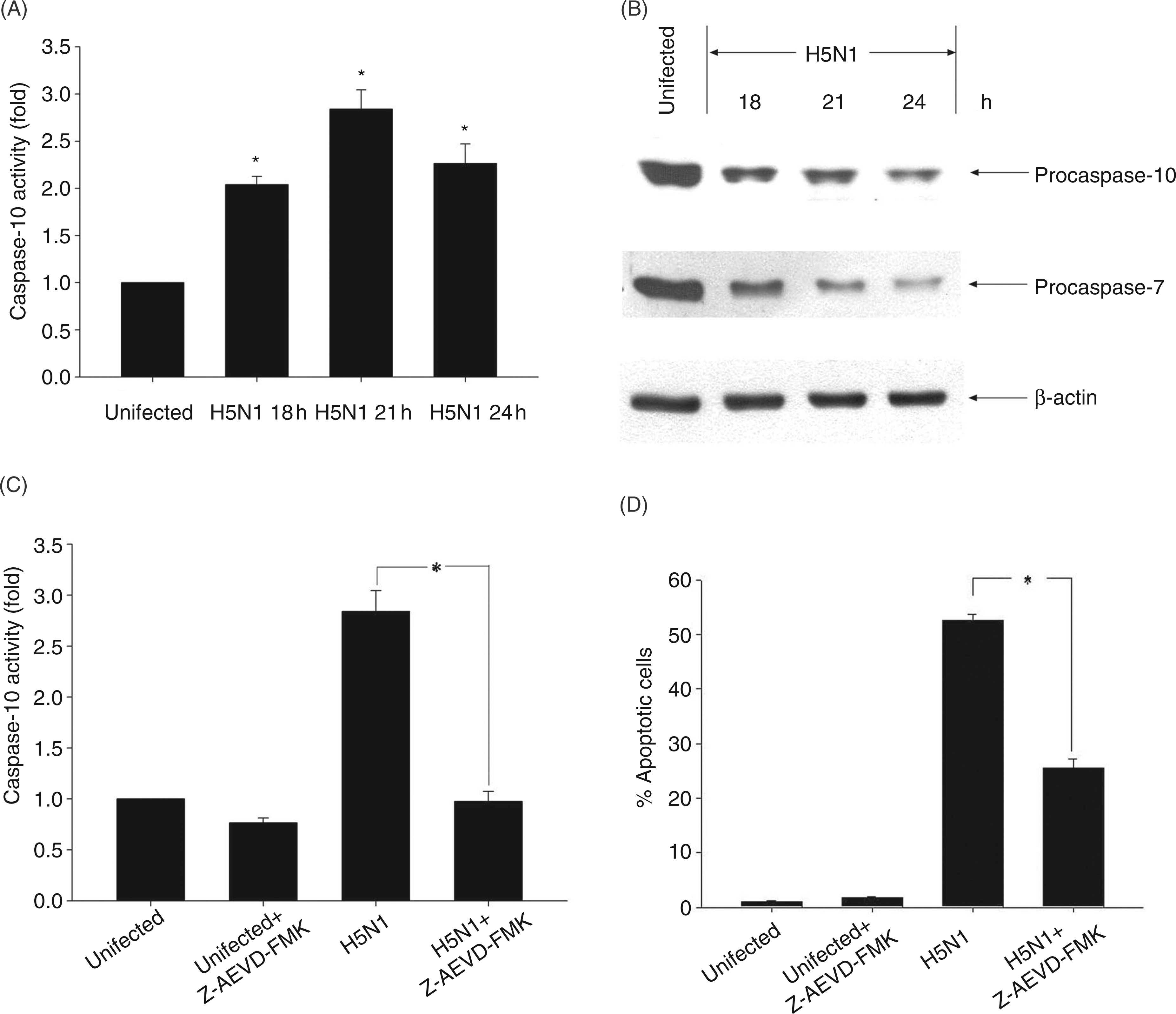
Activation of caspase-10 by H5N1 triggers Bid cleavage and release of AIF
As activation of caspase-10 is known to participate in the mitochondrial apoptotic pathway by cleaving the Bid molecule,
20
we analyzed Bid cleavage in avian H5N1 virus-infected MDMs. As shown in Figure 3A, inactive full-length Bid was gradually degraded in infected cells and was accompanied by the appearance of its cleavage product tBid. It has been previously documented that, after translocation to mitochondria, tBid triggers a change in mitochondrial membrane potential, thus causing it to release mitochondrial intermembrane components, such as AIF;
21
our results support this conclusion. Data presented in Figure 3A showed that AIF could be detected in the cytosol 21 h following infection. These results clearly suggest that avian H5N1 virus could activate Bid, causing infected cells to release AIF from mitochondria.
Bid cleavage and AIF release in H5N1-infected human MDMs. Human MDMs (A) or Z-AEVD-FMK-pretreated cells (B) were infected with H5N1 virus at multiplicity of infection (MOI) of 1. At the times indicated, cells were harvested and cytosolic proteins were prepared. Immunoblotting was performed to detect Bid, tBid and AIF. Actin was used as loading control. The results shown are representative of triplicate experiments.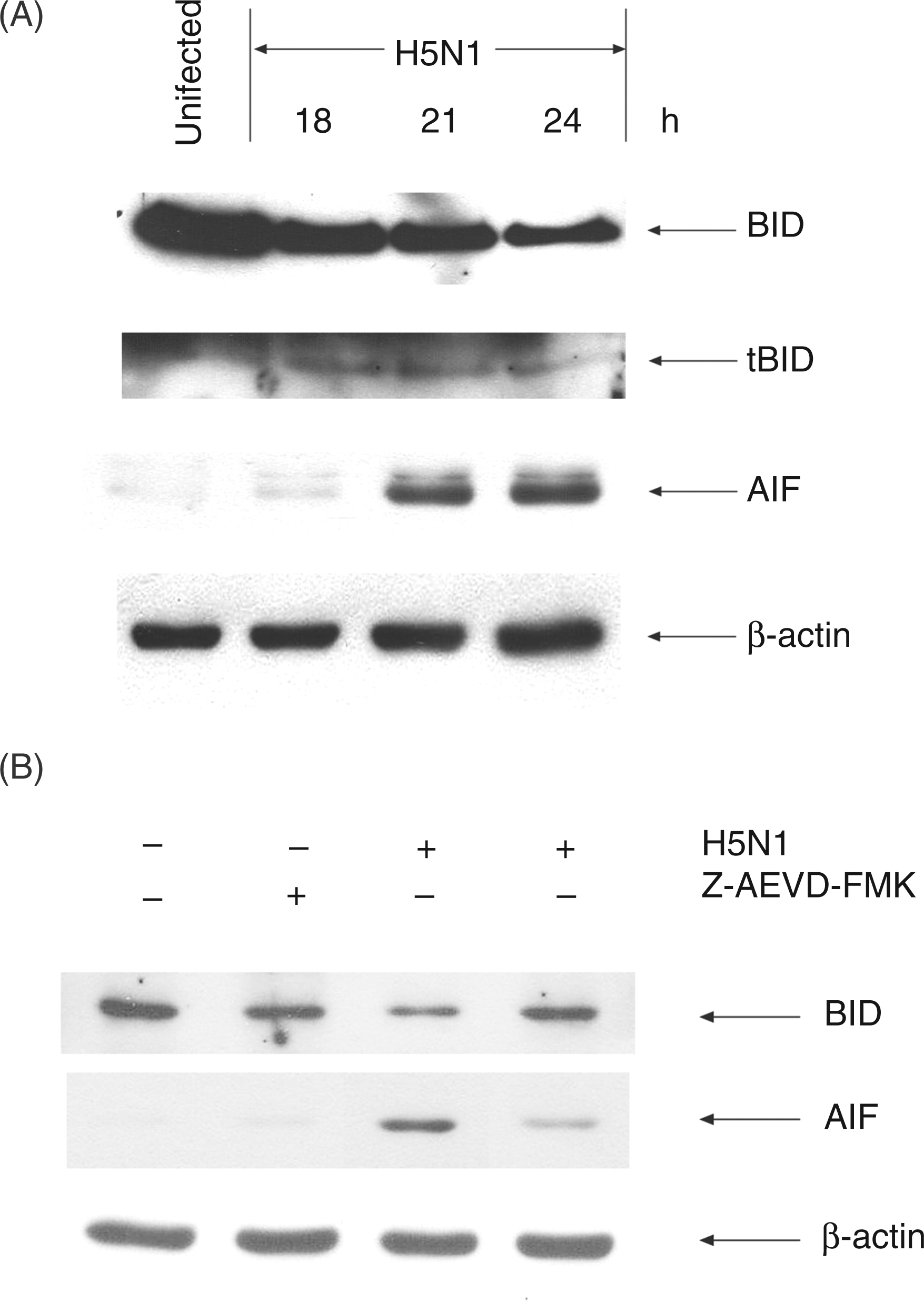
In order to investigate the involvement of caspase-10 in Bid cleavage and AIF release, caspase-10 inhibitor, Z-AEVD-FMK, was added to the test system as above. In the presence of this inhibitor, the degradation of Bid in H5N1-infected cells was not observed compared with that of infected cells that were not pretreated with the inhibitor (Figure 3B). These results suggested that the activation of Bid requires caspase-10 activation. In addition, Z-AEVD-FMK also suppressed the release of AIF from mitochondria, suggesting that this enzyme is also involved in AIF release, which may lead to apoptosis in H5N1-infected MDMs (Figure 3B).
Secreted TRAIL is required for caspase-10 activation
It was previously reported that the production of TRAIL was significantly upregulated in H5N1-induced apoptosis in avian H5N1 virus-infected MDMs.
17
In our system, the level of secreted TRAIL from H5N1-infected MDMs increased to 16 pg/ml at 24 h after infection (Figure 4A). To further investigate the relationship of TRAIL and caspase-10 with the ability to trigger cell death in H5N1 infection, the experimental protocol similar to that described earlier was performed in the presence or absence of TRAIL Ab (anti-TRAIL). A high dose of anti-TRAIL (1 µg/ml) was added to human MDMs at the time of infection. The number of apoptotic cells was determined after 24 h of infection. In the presence of anti-TRAIL Ab, the percentage of apoptotic cells was significantly lower when compared to the H5N1 infection alone or in the presence of isotype controls (Figure 4B). It should be noted that anti-TRAIL Ab by itself had no effect on the viability of uninfected cells. Our results suggest that TRAIL plays a role in inducing cell death in H5N1-infected macrophages.
Anti-TRAIL Ab interferes with induction of apoptosis in avian H5N1 virus-infected MDMs. Human MDMs were infected with H5N1 at multiplicity of infection (MOI) of 1 for 18, 21 and 24 h and secreted TRAIL in supernatant was measured by ELISA (A). MDMs were pretreated with 1 µg/ml anti-TRAIL before being infected with H5N1 at MOI of 1. The number of apoptotic cells (B) and caspase-10 activity (C) were determined by TUNEL staining and colorimetric assay respectively. Cleavage of procaspase-7, Bid activity and AIF release were determined by immunoblotting and actin was used as loading control (D). Data (A, B and C) represented mean ± SE from three independent experiments, *P<0.05. The results shown in (D) are representatives of triplicate experiments.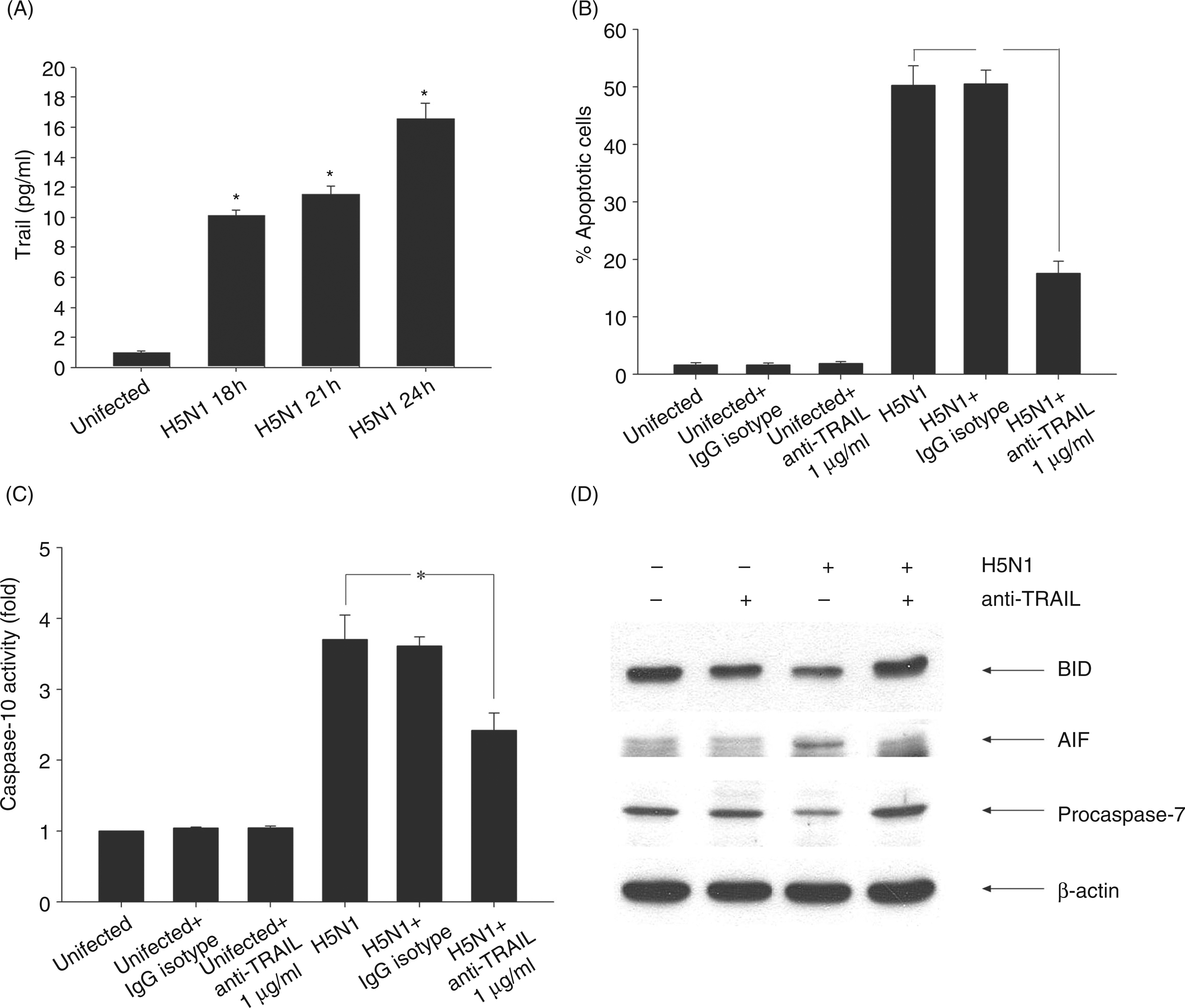
As TRAIL is also known to initiate apoptosis by the recruitment of FADD and caspase-10 to DISC in other infection models,14,16 it is of interest to further investigate whether TRAIL may be also involved in caspase-10 activation in H5N1-infected macrophages. Therefore, caspase-10 activity of H5N1-infected MDMs was determined in the presence of anti-TRAIL. As shown in Figure 4C, anti-TRAIL significantly reduced caspase-10 activity when compared to the H5N1-infected cells in the absence of the Ab or isotype control. These results suggest that TRAIL is involved in caspase-10 activation.
As the activation of caspase-10 by H5N1 could occur by the intrinsic pathway via Bid cleavage, or by the extrinsic pathway via caspase-7 activation, the experiments were set up to determine the pathway associated with this infection using anti-TRAIL Ab. As shown in Figure 4D, the levels of Bid and procaspase-7 degradation did not alter in the presence of anti-TRAIL Ab when compared to uninfected cells, but markedly increased compared with infected cells (Figure 4D). Under the same conditions, the level of AIF in cytosol appeared to be decreased in the presence of anti-TRAIL Ab (Figure 4D). The results of the activation of Bid and caspase-7 presented herein suggest that apoptosis of H5N1-infected MDMs involves both the intrinsic and extrinsic pathways.
Discussion
Infection with H5N1 virus is associated with a high mortality rate in humans. The virus not only infects the respiratory system, but can also induce blood-borne systemic infection. 5 Among the clinical manifestations, lymphopaenia has been suggested to contribute to disease pathogenesis.3,7 The H5N1 virus has also been reported to induce cell death in a variety of cell types, including alveolar epithelial cells, dendritic cells and macrophages.8,9,18 An autopsy from an H5N1-infected patient exhibited a high degree of macrophage cell death, suggesting, therefore, that the activation of macrophage cell death may contribute to the pathogenesis of the disease. 4 The mechanism involving H5N1-inducing cell death is still not fully understood. It was previously demonstrated that H5N1 virus isolated from infected patients was able to induce apoptosis by upregulating TRAIL and TNF-α, but not FasL, suggesting that TRAIL may play a role in the apoptosis of infected MDMs. 17 In the present study, we attempted to provide additional insights into the molecular mechanism of apoptosis in human MDMs infected with H5N1 isolated from A/open-billed stork. Our data demonstrated that the avian H5N1 virus could induce a significant degree of apoptosis in human MDMs (Figure 1). This result is compatible with a previous report showing that the virus is highly lethal to human monocytes. 8
Among the caspase enzymes, caspase-10 shares a similar structure to caspase-8 which functions as an initiator caspase in apoptotic pathway. Death ligands such as FasL and TRAIL have been shown to induce apoptosis by stimulating caspase-10 activation.14,15,20 In our study, we demonstrated that caspase-10 activation occurred in H5N1-infected cells (Figures 2A, B). Furthermore, the activation of caspase-7 and Bid cleavage in these infected human MDMs were also observed (Figures 2B, 3A). These results are in accordance with a previous study, suggesting that activation of caspase-10 led to activation of a pro-apoptotic molecule, Bid, and of a downstream caspase, caspase-7. 20 The cleavage form of Bid (tBid) is known to translocate to the mitochondria which then triggers change of mitochondrial membrane potential, thus resulting in the release of mitochondrial intermembrane components, such as AIF (Figure 3A). In addition, in the presence of caspase-10 inhibitor, the activation of Bid and AIF release were significantly reduced (Figure 3B). It should also be noted that in the presence of caspase-10 inhibitor, degradation of procaspase-7 was markedly reduced, suggesting that the activation of caspase-7 is downstream of caspase-10 (data not shown). Moreover, the number of apoptotic cells in avian H5N1-infected MDMs was also significantly depressed in the presence of caspase-10 inhibitor (Figure 2D). These results indicated that the activation of caspase-10 may contribute to apoptosis in avian H5N1-infected MDMs by the activation of Bid, resulting in the release of AIF from mitochondria.
TRAIL has been demonstrated to play an essential role in apoptosis in many host cell infection systems. 17 A previous study demonstrated that TRAIL secreted from the virus-infected human MDMs induced apoptosis in human Jurkat T cells, suggesting the ‘paracrine’ cytotoxic effect of TRAIL derived from the virus-infected cells on T cells. 17 The activation of TRAIL is known to trigger apoptosis by activating downstream caspases, such as caspase-8 and caspase-10. 14 In our study, we demonstrated that in the presence of the antibody against TRAIL, the number of apoptotic cells was significantly suppressed (Figure 4B). This result is consistent with the data showing that the Ab against TRAIL also interfered with the activation of caspase-10 activity (Figure 4C). However, it is possible that activation of caspase-10 may involve other pathways, as the anti-TRAIL Ab could not completely reduce caspase-10 activity to baseline level (Figure 4C). Although the role of FasL in caspase-10 activation in other systems is well established, this ligand was not upregulated in avian H5N1 virus-infected MDMs, 17 suggesting that it is not involved in H5N1 infection and that other components may be involved in caspase-10 activation.
Viral proteins have been demonstrated to be able to directly process procaspase, for example HIV-protease is able to cleave procaspase-8 which then activates downstream caspases, leading to apoptosis. 22 However, avian H5N1 possessing viral proteins directly activating caspase-10 remains to be investigated. In the present study, we have clearly demonstrated that, in the presence of anti-TRAIL Ab, activation of caspase-10, Bid and caspase-7 was inhibited, resulting in a reduction of AIF release (Figure 4D). These data implied that TRAIL, in autocrine fashion, regulates apoptosis in avian H5N1-infected MDMs through the activation of caspase-10.
Considered together, the molecular data in the present study allow us to propose a possible mechanism leading to apoptosis in avian H5N1 virus infection of MDMs. Exposure to avian H5N1 virus leads to the activation of TRAIL, resulting in an increase of caspase-10 activity. The latter then cleaves Bid to its truncated active form that translocates to mitochondria to induce the release of AIF. Moreover, caspase-10 activation also activates caspase-7 resulting in apoptotic cell death. The release of mitochondrial intermembrane space apoptotic proteins is known to activate several other caspases including caspase-3. 23 The activation of caspase-3 in avian H5N1 virus-infected MDMs has also been reported. 18 Moreover, activation of caspase-7 has also been reported to participate in caspase-3 activation. Therefore, it is also possible that the activation of caspase-7 observed in our study may also contribute to caspase-3 activation Furthermore, inhibition of the cleavage of caspase-7 and inactivation of Bid degradation, together with the release of AIF to cytosol in H5N1-infected cells by anti-TRAIL Ab presented in this study, implied that the extrinsic pathway may also activate the intrinsic pathway in avian H5N1 virus-infected MDMs.
Our results suggest that the influenza virus is capable of activating cell death via both extrinsic and intrinsic apoptotic pathways. Understanding the mechanisms of how H5N1 virus induces apoptosis in host cells may lead to developing novel therapeutic agents that can alleviate the severity, and reduce the mortality rate, of this infectious disease.
Conclusion
This study elucidated a possible mechanism of apoptosis in avian H5N1-infected MDMs. The data are consistent with the conclusion that apoptosis was initiated by TRAIL, which has an autocrine cytotoxic effect, and induced caspase-10 activation, which led to Bid cleavage and AIF release.
Footnotes
Funding
This work was supported by National Institute of Health and National Institute of Allergy and Infectious Disease (YI-AI-5026-01).