
Abstract
Nomilin is a triterpenoid present in common edible citrus fruits with putative anticancer properties. In this study, the authors investigated the antimetastatic potential of nomilin and its possible mechanism of action. Metastasis was induced in C57BL/6 mice through the lateral tail vein using highly metastatic B16F-10 melanoma cells. Administration of nomilin inhibited tumor nodule formation in the lungs (68%) and markedly increased the survival rate of the metastatic tumor–bearing animals. These results correlated with the biochemical parameters and histopathological analysis. Nomilin showed an inhibition of tumor cell invasion and activation of matrix metalloproteinases. Treatment with nomilin induced apoptotic response, characterized by an increase in the sub-G1 fraction of cells with chromatin condensation and membrane blebbing, a typical ladder of DNA fragmentation, and detection of apoptotic cells by TUNEL assay. Nomilin treatment also exhibited a downregulated Bcl-2 and cyclin-D1 expression and upregulated p53, Bax, caspase-9, caspase-3, p21, and p27 gene expression in B16F-10 cells. Proinflammatory cytokine production and gene expression were found to be downregulated in nomilin-treated cells. The study also reveals that nomilin could inhibit the activation and nuclear translocation of antiapoptotic transcription factors such as nuclear factor (NF)-κB, CREB, and ATF-2 in B16F-10 cells.
Keywords
Introduction
Cancer is a hyperproliferative disorder that involves transformation, dysregulation of apoptosis, proliferation, invasion, angiogenesis, and metastasis. Metastasis is the most lethal attribute of a cancer. It is the leading cause of cancer-related mortality and is the single most important obstacle to curative cancer therapy. During this process, tumor cells disseminate from the primary tumor, migrate through the basement membrane, survive in the circulatory system, invade into a secondary site, and start to proliferate. 1 The metastatic process is highly inefficient; very few of the cells that migrate from the primary tumor successfully colonize at distant sites. Recent in vitro and in vivo observations indicate that apoptosis is an important process in regulating metastasis. 2
Apoptosis, or programmed cell death, is an essential event that plays an important role in an organism’s developmental process, maintenance of homeostasis, and elimination of damaged cells. Its morphological characteristics include plasma membrane blebbing, cell shrinkage, nuclear condensation, chromosomal DNA fragmentation, and formation of apoptotic bodies. 3 Tumors are characterized by uncontrolled proliferation and reduced apoptosis. Inducing apoptosis is an efficient method of treating cancers. 4 Cell cycle control has been proven to be a major event in ensuring accurate cell division. Abnormalities of cell cycle regulators have been associated with many carcinogenic processes. Thus, cell cycle regulators of cancer cells could be targeted and altered as a chemoprevention treatment. 5
Natural products represent a rich source of compounds that have found many applications in various fields of medicines and therapy, including cancer therapy. Effective ingredients in several plant-derived medicinal extracts are terpenoid compounds, and many terpenes have biological activities that are used for the treatment of human diseases. Nomilin (supplementary Figure 1) is a triterpenoid present in common edible citrus fruits with putative anticancer properties. Immunomodulatory activity of this triterpenoid was already reported in our lab. Treatment with nomilin was found to enhance the total white blood cell count, bone marrow cellularity, α-esterase-positive cells, antibody titer, and the number of plaque forming cells in the spleen and also remarkably inhibited delayed-type hypersensitivity reaction in Balb/c mice. 6 It also possesses antiproliferative activity against a number of human cancer cell lines, including that for leukemia (HL-60) and cancers of the ovary (SKOV-3), cervix (HeLa), stomach (NCI-SNU-1), liver (Hep G2), and breast (MCF-7). 7 In the present study, the effect of nomilin on the inhibition of pulmonary metastasis induced by B16F-10 melanoma cells in C57BL/6 mice was evaluated with special emphasis on the mechanism of action, using in vitro models.

Chemical structure of nomilin (C28H34O9)
Materials and Methods
Animals
C57BL/6 mice (20-25 g body weight, 6- to 8-week-old males) were purchased from National Institute of Nutrition, Hyderabad, India. The animals were fed with mouse chow (Sai Feeds, India) and water ad lib. All the animal experiments were conducted according to the rules and regulations of the Animal Ethical Committee, Government of India.
Cell Line
Highly metastatic B16F-10 melanoma cells were obtained from the National Centre for Cell Science, Pune, India. The cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS) and antibiotics. DMEM was purchased from Himedia Laboratory, Mumbai, India.
Cells–cDNA kit was purchased from Ambion Inc (Austin, TX). CycleTEST PLUS DNA Reagent Kit (Becton-Dickson, Canada), ApopTag Peroxidase In Situ Apoptosis Detection Kit was purchased from CHEMICON International, Inc (Billerica, MA). Oligonucleotide primer sequences were purchased from Maxim Biotech Inc (San Francisco, CA; Table 1). All other reagents used were of analytical reagent grade.
Primer Sequences of Different Genes

Abbreviations: TNF, tumor necrosis factor; IL, interleukin; GM-CSF, granulocyte macrophage colony-stimulating factor.
Drug Administration
Nomilin was purchased from Sigma (St Louis, MO). It was suspended in light paraffin oil and administered at a concentration of 6 mg/kg body weight. The drug was administered starting from the same day as tumor inoculation and was continued for 10 consecutive days.
Determination of the Effect of Nomilin on Metastasis of B16F-10 Melanoma Cells In Vivo
C57BL/6 mice (20-25 g body weight, 6- to 8-week-old males) were divided into 3 groups (8 animals/group). All the animals were induced with metastasis by injecting highly metastatic BI6F-10 melanoma cells (106 cells/ animal) through the lateral tail vein. Group 1 mice were injected with 10 doses of nomilin (6 mg/kg body weight, intraperitoneally). Group II consisted of the vehicle controls who were injected with 10 doses of paraffin oil. Group III consisted of untreated tumor-bearing controls. All the animals were killed on the 21st day after tumor implantation; lungs were excised, and serum was collected. The lungs were used for assessing various parameters: (a) morphological examinations of metastatic tumor nodules, (b) histopathological analysis, and (c) estimation of collagen hydroxyproline, 8 hexosamine, 9 and uronic acid. 10 Serum was used for determining the levels of sialic acid 11 and gamma glutamyl transpeptidase (γGT). 12
A parallel set of experiments was conducted, and the death of animals resulting from tumor burden was recorded. The increase in life span as compared with the control animals was calculated using the formula ([T − C]/C) × 100, where T is the life span of treated animals and C the life span of control animals.
Determination of the Effect of Nomilin on Metastasis of B16F-10 Melanoma Cells In Vitro
The antimetastatic activity of nomilin in the in vitro system was assessed using the following parameters.
Cell viability assay
B16F-10 melanoma cells (5000 cells/well) were plated in a 96-well flat-bottomed titer plate and incubated at 37°C in 5% CO2 atmosphere. After 24 hours, escalating concentrations of nomilin (5-500 µg/mL) were added, and the incubation was continued for 48 hours under the same conditions. The percentage of viable cells was determined using the MTT assay.
Collagen matrix invasion assay
The invasion assay was carried out in Boyden chambers as described by Albini et al. 13 Briefly, the lower compartment of the chamber was filled with serum-free DMEM, and a polycarbonate filter membrane coated with type-I collagen was placed on it. B16F-10 cells were then seeded onto the upper chamber.
To test the effect on the invasion of B16F-10 melanoma cells, nomilin, at nontoxic concentrations (25, 10, and 5 µg/mL), was added along with the cells to the upper chamber, and this was incubated for 10 hours at 37°C. All the experiments were performed in triplicate, and the results are expressed as percentage inhibition of invasion.
Gelatin zymography
Gelatin zymography SDS-PAGE was performed with 5% gelatin incorporated in the separating gel. 14 B16F-10 melanoma cells of subconfluent cultures were incubated with serum-free medium for 24 hours at 37°C in 5% CO2 atmosphere. The conditioned medium was then collected and subjected to zymographic analysis; 50 µL of sample (equivalent to 100 µg protein) was activated with 5 µL trypsin solution (75 µg/mL) in the presence and absence of nomilin (25, 10, and 5 µg/mL) in 0.1 M Tris–HCl, 10 mM CaCl2 buffer (pH 8.0) and incubated for 1 hour at room temperature. Samples were mixed with an equal volume of 2× sample buffer and loaded on to 11% polyacrylamide gels containing 5% gelatin. Electrophoresis was carried out at 4°C with a constant current of 2 mA/tube until the tracking dye reached the periphery. The gels were then washed with 2% Triton X-100 in 0.1 M Tris–HCl, 10 mM CaCl2 at 37°C for 18 hours followed by staining with Gelcode Blue stain reagent for 2 hours. Gels were destained to visualize the clear area against the dark background.
The mechanism of action of nomilin in the inhibition of metastasis was studied by following parameters using in vitro systems.
Determination of the Effect of Nomilin on the Apoptosis of B16F-10 Cells
The effect of nomilin on the apoptosis of B16F-10 melanoma cells was analyzed using various parameters such as morphological analysis, DNA fragmentation analysis, cell cycle analysis, Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay, and expression studies of proapoptotic and antiapoptotic genes by reverse transcriptase polymerase chain reaction (RT-PCR).
Morphological analysis
B16F-10 melanoma cells (5000 cells/well) suspended in DMEM supplemented with 10% FCS, 100 µg/mL streptomycin and penicillin, and 2 mmol/L glutamine were plated in a 96-well flat-bottomed titer plate and incubated for 24 hours at 37°C in 5% CO2 atmosphere. After 24 hours, different concentrations of nomilin (25, 10, and 5 µg/mL) were added to the cells, and this was incubated further for 48 hours under the same conditions. The cells were then washed with phosphate-buffered saline (PBS; pH 7.4), fixed with 5% formalin and stained using hematoxylin and eosin, observed under phase-contrast microscopy, and photographs were taken. Apoptosis was characterized by examining the morphological changes, such as chromatin condensation, nuclear condensation, membrane blebbing, or presence of apoptotic bodies.
DNA fragmentation analysis
A total of 1 × 106 B16F-10 melanoma cells were treated with different concentrations of nomilin (25, 10, and 5 µg/mL) as described in the previous experiment. After incubation, the cells were treated with 0.1 mL lysis buffer (100 mmol/L Tris–HCl, pH 8.0, containing 0.2% Triton-X 100, and 1 mmol/L ethylenediaminetetraacetic acid [EDTA]) for 10 minutes at −20°C. DNA was extracted using the phenol–chloroform method, precipitated with chilled ethanol, and resuspended in Tris–EDTA buffer (10 mmol/L Tris–HCl, pH 8.0, and 1 mmol/L EDTA). DNA samples were separated by electrophoresis in 1% agarose gels. DNA was stained with ethidium bromide and photographed under UV light.
Determination of the Effect of Nomilin on Cell Cycling of B16F-10 Melanoma Cells
The cell apoptotic rate was measured by quantification of the percentage of cells with sub-G1 DNA levels, as determined via flow cytometry analysis. Briefly, 1 × 106 logarithmically growing B16F-10 cells, suspended in DMEM with 10% FCS, were seeded in a culture flask. These cells were treated with nomilin (25 µg/mL) and incubated for 10 hours at 37°C in CO2 atmosphere. After incubation, the cells were washed with PBS and stained with propidium iodide. DNA content was analyzed by a fluorescence-activated cell sorter (FACS, Becton Dickinson FACS Calibur) using the CycloTEST PLUS DNA Reagent Kit per manufacturer’s protocol.
TUNEL assay
TUNEL assay was done to detect apoptosis via DNA fragmentation using ApopTag Peroxidase In Situ Apoptosis Detection Kit, CHEMICON International Inc. B16F-10 melanoma cells (5000 cells/well) suspended in DMEM supplemented with 10% FCS, 100 µg/mL streptomycin and penicillin, and 2 mmol/L glutamine were plated in 96-well flat-bottomed titer plate and incubated for 24 hours at 37°C in 5% CO2 atmosphere. After 24 hours, 25 µg/mL nomilin was added to the cells, and this was incubated further for 48 hours under the same conditions. The cells were washed in PBS and stained per manufacturer’s protocol. The number of apoptotic cells was counted and expressed in terms of the percentage of cells.
Determination of the Effect of Nomilin on the Expression of p53, Caspase-9, Caspase-3, Caspase-8, Bcl-2, Bid, Bax, p21, p27, and Cyclin-D1 Using RT-PCR
B16F-10 melanoma cells (2 × 104 cells) suspended in serum-free DMEM (250 µL) were seeded in a 96-well titer plate and incubated for 24 hours at 37°C in 5% CO2 atmosphere. Nomilin at a concentration of 25 µg/mL was added, and incubation was continued for another 4 hours. cDNA was synthesized using a cell–cDNA kit (Ambion Inc.). Cells were washed with PBS and heated in cell lysis buffer (provided in the kit) to release the RNA into the solution followed by a heating step to inactivate endogenous RNases. The genomic DNA was further degraded by treating with DNase followed by inactivation of DNase by heating at 70°C. Reverse transcription was performed at 42°C for 50 minutes using Moloney murine leukemia virus RT (supplied along with the kit). Gene expression analysis was done by PCR. The mouse p53, caspase-9, caspase-3, caspase-8, Bcl-2, Bid, Bax, p21, p27, and Cyclin-D1 (Table 1) were amplified against the GAPDH standard. The cycling conditions used were as follows: 1 minute at 94°C, 1 minute at 58°C, and 1 minute at 72°C for 40 cycles, followed by a 10-minute extension at 72°C.
Amplified samples were subjected to electrophoresis in an agarose gel (1.5%) containing 0.5 µg/mL ethidium bromide and photographed under UV light.
Determination of the Effect of Nomilin on Gene Expression of Proinflammatory Cytokines and Granulocyte Macrophage Colony-Stimulating Factor (GM-CSF)
Proinflammatory cytokine gene expression studies were done using specific mouse primers of tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, GM-CSF, and the housekeeping gene GAPDH (Table 1). The following PCR reaction profile was followed in the minicycler: 1 minute at 94°C, 1 minute at 55°C, and 1 minute at 72°C for 39 cycles followed by a 10-minute extension at 72°C. Amplified PCR products were subjected to electrophoresis on a 1.8% agarose gel, stained with ethidium bromide, and photographed under UV light.
Determination of the Effect of Nomilin on Proinflammatory Cytokines and GM-CSF Levels
B16F-10 melanoma cells (5000 cells/well) suspended in DMEM supplemented with 10% FCS, 100 µg/mL streptomycin and penicillin, and 2 mmol/L glutamine were plated in a 96-well flat-bottomed titer plate and incubated for 24 hours at 37°C in 5% CO2 atmosphere. Nomilin at a concentration of 25 µg/mL was added to the cells, and this was incubated further for 48 hours under the same conditions. The supernatant was used for the estimation of cytokines such as IL-1β, IL-6, TNF-α, and GM-CSF using specific ELISA kits per manufacturer’s protocol (Pierce Biotechnology, USA).
Determination of the Effect of Nomilin on Nuclear Factor Kappa-B (NF-κB), c-fos, ATF-2, and CREB-1
Preparation of nuclear extracts
Nuclear extracts were prepared by a previously published method. 15 B16F-10 cells were exposed to nomilin (25 µg/mL) for 2 hours, washed with PBS twice, and then treated with TNF-α for 30 minutes and lysed using a lysis buffer for 15 minutes on ice. The cell suspension was centrifuged, cells were disrupted mechanically, and centrifuged at 10 000g to 11 000g for 20 minutes. The crude nuclear pellet obtained was suspended in a nuclear extraction buffer. Nuclei were disrupted using a fresh syringe, centrifuged, and the supernatant was collected. Protein concentrations of the nuclear extracts were estimated using the standard Bradford method and stored at −70°C.
Transcription factor profiling
Transcription factor profiling was done using a BD Mercury Transfactor Kit obtained from BD Biosciences (Palo Alto, CA). This kit provided rapid, high-throughput detection of specific transcription factors—namely, NF-κBp65, NF-κBp50, NF-κBc-Rel, c-Fos, ATF-2, and CREB-1—in the nuclear extract. Using an ELISA-based format, the transfactor kit detected the DNA-bound transcription factors 16 by specific primary antibody toward NF-κB p65, NF-κB p50, NF-κB, c-Rel, c-Fos, ATF-2, and CREB-1. A horseradish peroxidase–conjugated secondary antibody was then used to detect the bound primary antibody. The enzymatic product was measured with a standard microtiter plate reader at 655 nm. Percentage inhibition was calculated by the formula 100 − ([Optical Density of treated/Optical Density of control] × 100).
Statistical Analysis
The differences among experimental and control groups were determined using statistical software (GraphPad Instat software package). Comparisons among different groups were performed by analysis of variance using 1-way ANOVA. Significant differences between control and experimental groups were further analyzed by Bonferroni’s multiple comparison posttest. P values <.05 were considered as significant.
Results
Effect of Nomilin on Metastatic Lung Tumor Nodule Formation
The effect of nomilin on the inhibition of pulmonary tumor nodules is shown in Table 2. Untreated control animals developed a massive number of tumor nodules, which was given an arbitrary number of 250. 17 Administration of the triterpenoid nomilin inhibited tumor nodule formation remarkably. In the vehicle (paraffin oil)-treated group of animals, tumor nodule formation was similar to that of untreated controls. Histopathological analysis also supports the above observation, showing reduced lung fibrosis and accumulation of extracellular proteins in the alveolar walls of lungs of nomilin-treated animals when compared with untreated tumor controls (Figure 1). The effect of nomilin on the survival rate of metastatic tumor-bearing mice is shown in Table 2. The control metastatic tumor-bearing mice survived only up to 32 days. Administration of nomilin increased the life span of tumor-bearing animals (Figure 1).
Effect of Nomilin on Lung Tumor Colony Formation and Survival Rate a

B16F-10 melanoma cells (1 × 106) were injected into each animal via the lateral tail vein. Nomilin was administered intraperitoneally for 10 days. Control animals (n = 8) were treated with vehicle paraffin oil. Animals were killed on the 21st day and lung tumor nodules counted. For the survival study, death caused by tumor burden was recorded, and the life span was calculated.
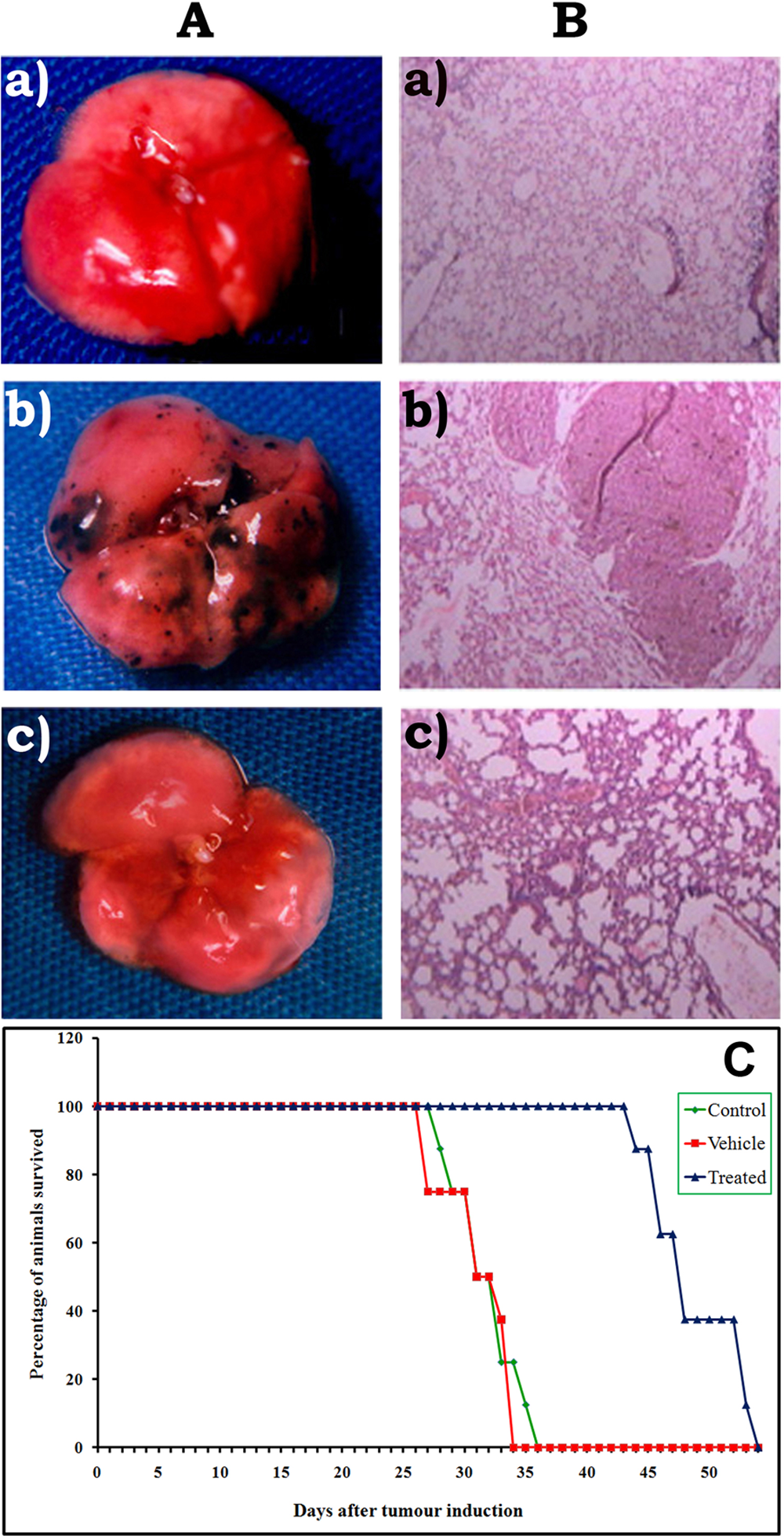
Lungs of metastatic tumor–bearing mice. A. Lung tumor nodule formation: (a) normal, (b) control with tumor nodules, (c) nomilin treated. B. Histopathological analysis: (a) normal, (b) control, (c) nomilin treated. C. Survival plot
Lung Biochemical Parameters
The effect of nomilin on lung collagen hydroxyproline, hexosamine, and uronic acid contents in metastatic tumor-bearing animals is shown in Table 3. Both tumor controls and vehicle-treated animals showed a drastic increase in the level of lung collagen hydroxyproline, hexosamine, and uronic acid content, which was significantly (P < .05) reduced in animals treated with nomilin. Differences between tumor control and vehicle-treated groups were not statistically significant.
Effect of Nomilin on Lung Collagen Hydroxyproline, Hexosamine, and Uronic Acid Levels of Metastatic Tumor-Bearing Animals a

B16F-10 melanoma cells (1 × 106) were injected into each animal via the lateral tail vein. Nomilin was administered intraperitoneally for 10 days. Control animals (n = 8) were treated with vehicle paraffin oil. Animals were killed on the 21st day; lungs were dissected out, and the levels of lung OH-proline, hexosamine, and uronic acid were determined. Values are mean ± standard deviation.
Means without a common superscript differ (P < .05).
Serum Biochemical Parameters
The effect of nomilin on serum sialic acid and serum γGT levels of metastatic tumor–bearing animals is shown in Table 4. Serum sialic acid and serum γGT levels were elevated in the tumor controls and in vehicle-treated animals, but these were significantly (P < .05) lowered by the administration of nomilin. There was no statistically significant difference between tumor control and vehicle-treated groups.
Effect of Nomilin on Serum Sialic Acid and Serum γGT Levels of Metastatic Tumor–Bearing Animals a

B16F-10 melanoma cells (1 × 106) were injected into each animal via the lateral tail vein. Nomilin was administered intraperitoneally for 10 days. Control animals (n = 8) were treated with vehicle paraffin oil. Animals were killed on the 21st day; blood was collected by heart puncture and serum separated, and the levels of serum sialic acid and serum γ-glutamyl transferase (γGT) were determined. Values are mean ± standard deviation.
Means without a common superscript differ (P < .05).
Cell Viability Assay
Cytotoxicity of nomilin toward B16F-10 melanoma cells in vitro is shown in Table 5. Concentrations of 5 to 25 µg/mL of nomilin were found to be nontoxic to B16F-10 melanoma cells, and these concentrations were used for further in vitro experiments.
Cell Viability Assay a

B16-F10 melanoma cells (5000 cells/well) were plated in a 96-well titer plate with different concentrations of nomilin and incubated for 48 hours. The percentage of viable cells was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT) assay. Values are means of triplicate.
Collagen Matrix Invasion Assay
The invasiveness of metastatic B16F-10 melanoma cells through the collagen matrix is very high. Very large numbers of cells were found in the lower surface of the polycarbonate membrane, but administration of nomilin produced an inhibition in the invasion of the collagen matrix by the tumor cells in a dose-dependent manner. At a concentration of 25 µg/mL, nomilin significantly inhibited the invasion of B16F-10 melanoma cells by 59.3% (P < .05), whereas at 10 µg/mL and 5 µg/mL, the percentage of inhibition of invasion was found to be 44.9% (P < .05) and 26.8% (P < .05), respectively (Figure 2).

The effect of nomilin on collagen matrix invasion: B16F-10 melanoma cells (1 × 105 cells /150 µL Dulbecco’s modified Eagle’s medium [DMEM]) were seeded into collagen type-I-coated polycarbonate membrane on the upper compartment of the Boyden chamber. The lower compartment was filled with DMEM. Cells were incubated in the presence of nomilin for 10 hours at 370°C. The membrane was removed, fixed, and stained, and the cells that had migrated in the test and control groups were counted. Experiments were performed in triplicate, and the results are expressed as percentage inhibition of invasion. Values are mean ± standard deviation
Gelatin Zymographic Analysis
Zymographic analysis of the trypsin-activated B16F-10 melanoma cell-conditioned medium showed digested clear areas at 92 kDa and 72 kDa. Gels loaded with tumor cell culture supernatant without trypsin activation did not show any clear areas, indicating the inactive form of the enzyme collagenase. Trypsin-activated tumor cell culture supernatant loaded gels, after incubation with 10 mM EDTA, did not show clear areas, which indicates that EDTA inhibited enzyme activation and that the enzyme is metalloproteinase. The conditioned media of nomilin-pretreated cells, which were then activated by trypsin, did not show any clear band, indicating the inhibition of expression or activity of gelatinases by the treatment at concentrations of 25 and 10 µg/mL. The conditioned media of 5 µg/mL nomilin-pretreated cells did not show any inhibition of expression or activity of gelatinases by the treatment (Figure 3).

Effect of nomilin on the activation of matrix metalloproteinases: A. conditioned medium without trypsin activation. B. Trypsin activation + EDTA (ethylenediaminetetraacetic acid). C. Trypsin activation alone. D. Trypsin activation + nomilin (25 µg/mL). E. Trypsin activation + nomilin (10 µg/mL). F. Trypsin activation + nomilin (5 µg/mL).
Effect of Nomilin on Inducing Apoptosis of B16F-10 Cells
Apoptotic cells were characterized by cell shrinkage, DNA fragmentation, membrane blebbing, and presence of apoptotic bodies. B16F-10 cells treated with different doses of nomilin (25, 10, and 5 µg/mL) showed the presence of apoptotic bodies and membrane blebbing in a dose-dependent manner (supplementary Figure 2A). The maximum effect was on the cells treated with 25 µg/mL nomilin. The dose-dependent enhancement of DNA fragmentation in nomilin-treated cells as compared with the untreated controls also supports the above observation (supplementary Figure 2B available online).
TUNEL Assay
TUNEL assay also confirmed the presence of apoptotic bodies by staining free 3’-OH termini enzymatically labeled with modified nucleotides, which supports the above observation (supplementary Figure 2C). These new DNA ends that are generated on DNA fragmentation are typically localized in morphologically identifiable nuclei and apoptotic bodies. In contrast, the normal B16F-10 cells (control), which have relatively insignificant numbers of DNA 3’-OH ends, did not stain in the above experiment. Nomilin treatment yielded a significant (P < .05) induction of apoptosis in B16F-10 cells, and the number of apoptotic cells was found to be 44% (supplementary Figure 2D).
Cell Cycle Analysis
Figure 4 shows the effect of nomilin on the B16F-10 cell cycle. In untreated controls, 64.76% of the cells were in the G1-phase, 13.38% of the cells were in the G2/M-phase, and 14.81% of cells were in the S-phase. The percentage of cells with fragmented DNA (sub-G1) was only 2.32%. Treatment with 25 µg/mL nomilin increased the percentage of cells with fragmented DNA (sub-G1) to 36.02%, but the number of cells in G1, S, and G2/M phases decreased considerably (40.2%, 12.9%, and 7.08%, respectively, for nomilin).
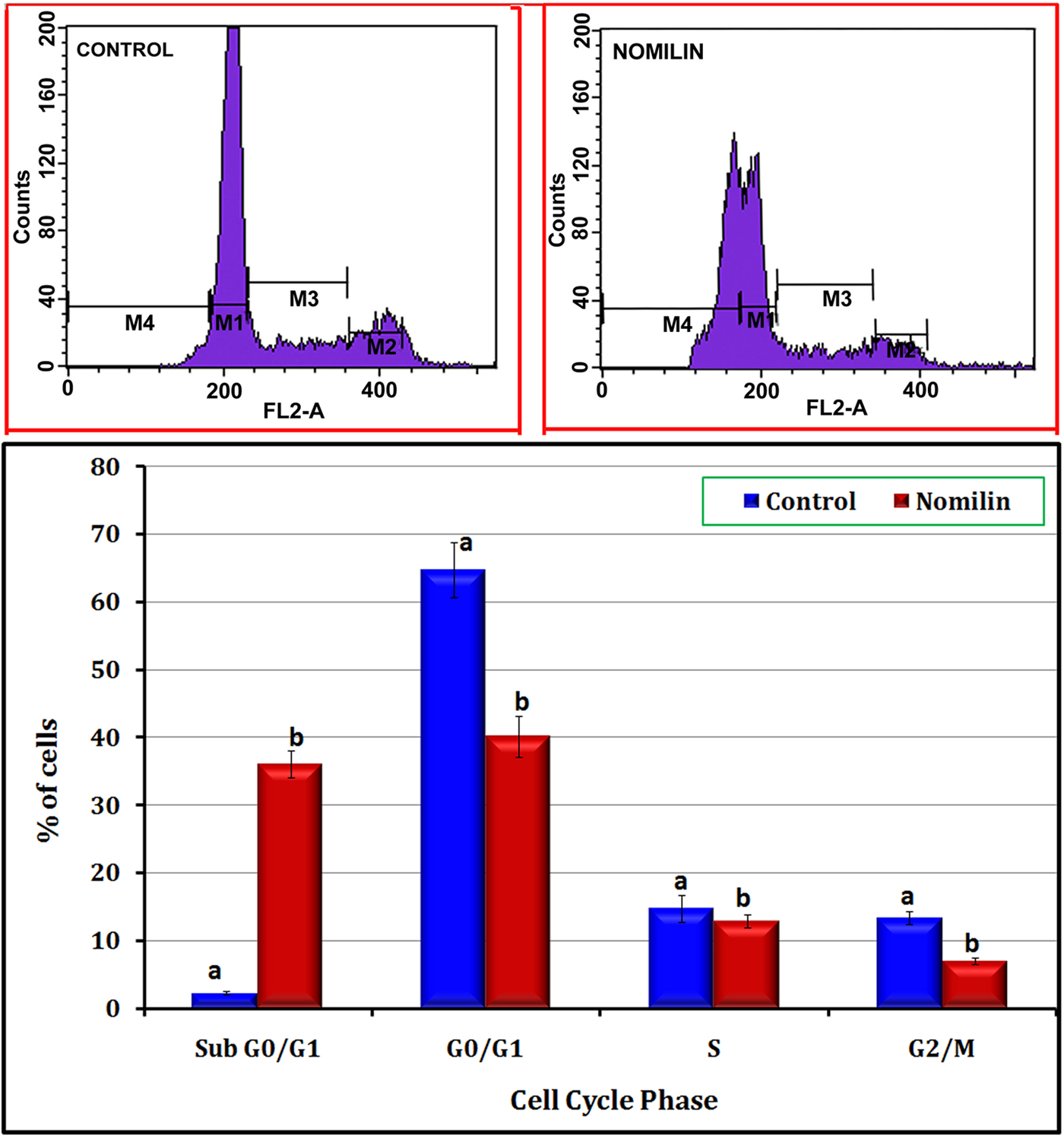
Effect of nomilin on cell cycle progression: B16F-10 melanoma cells were treated with nomilin (25 µg/mL) for 10 hours and analyzed for propidium iodide–stained DNA content by flow cytometry (FACS Calibur, Becton Dickinson Immunocytometry Systems, San Jose, CA). Values indicate the percentage of the cell population at the phase of the cell cycle. M1 = G1 (diploid), M2 = G2/M (tetraploid), M3 = S (synthetic phase), M4 = sub-G1 phase. The population of cells in the sub-G1 phase consists of cellular fragments resulting from apoptosis. Values are mean ± standard deviation
Effect of Nomilin on the Expression of p53, Caspase-9, Caspase-3, Caspase-8, Bcl-2, Bid, Bax, p21, p27, and Cyclin-D1 Genes Using RT-PCR
Agarose gel electrophoresis of the amplified samples shows (Figure 5) that the expression of p53, caspase-9, caspase-3, Bax, p21, and p27 were upregulated in nomilin (25 µg/mL)-treated cells compared with the nontreated control cells; caspase-8 did not show any expression (data not shown) in either nontreated controls or nomilin-treated cells, whereas bcl-2 and Cyclin-D1 genes showed a diminished expression in nomilin-treated B16F-10 cells.

Effect of nomilin on the expression of p53, Bax, caspase-3, caspase-9, bcl-2, p21, p27, Cyclin-D1, GM-CSF, TNF-α, IL-1β, and IL-6 genes and the housekeeping gene GAPDH in B16F-10 melanoma cells: B16F-10 cells (5 × 104) were treated with or without nomilin (25 µg/mL) for 4 hours, and cDNA was synthesized and amplified with an appropriate primer for p53, p53, Bax, caspase-3, caspase-9, bcl-2, p21, p27, Cyclin-D1, GM-CSF, TNF-α, IL-1β, IL-6, and GAPDH
Effect of Nomilin on GM-CSF and Proinflammatory Cytokine Production and Gene Expression
The effect of nomilin on GM-CSF and proinflammatory cytokine gene expression is shown in Figure 5. Expression of GM-CSF and the proinflammatory cytokines TNF-α, IL-1β, and IL-6 genes was downregulated in B16F-10 melanoma cells by treatment with nomilin. Nomilin could significantly (P < .05) inhibit the production of TNF-α, IL-1β, IL-6, and GM-CSF by B16F-10 melanoma cells in culture (Table 6).
Effect of Nomilin on the Release of TNF-α, IL-1β, IL-6, and GM-CSF by B16F-10 Melanoma Cellsa

Abbreviations: TNF, tumor necrosis factor; IL, interleukin; GM-CSF, granulocyte macrophage colony-stimulating factor.
B16F-10 cells (5 × 103 cells) were cultured in the presence of nomilin (25 µg/mL) for 48 hours, and concentration in the culture supernatant was estimated. Values are expressed as mean ± standard deviation (mean of triplicate).
Means without a common superscript differ (P < .05).
Effect of Nomilin on NF-κB, c-fos, ATF-2, and CREB-1
The effect of nomilin on the activation of transcription factors is shown in Figure 6. Nomilin could inhibit the activation and nuclear translocation of NF-κB subunits, p65 (52.3%), p50 (62.9%), c-Rel (65.8%), and also other transcription factors such as c-fos (71.6%), ATF-2 (73.4%), and CREB-1 (62.5%).

Effect of nomilin on the translocation of transcription factors: B16F-10 cells were grown in a 25 cm2 culture flask and treated with nomilin (25 µg/mL) for 2 hours at 37°C in 5% CO2. Cells were washed with phosphate buffered saline and incubated with TNF-α (10 pg/mL) for 30 minutes at 37°C in 5% CO2. Nuclear extracts were prepared separately and subjected to transfactor assay. Values are mean ± standard deviation (mean of triplicate)
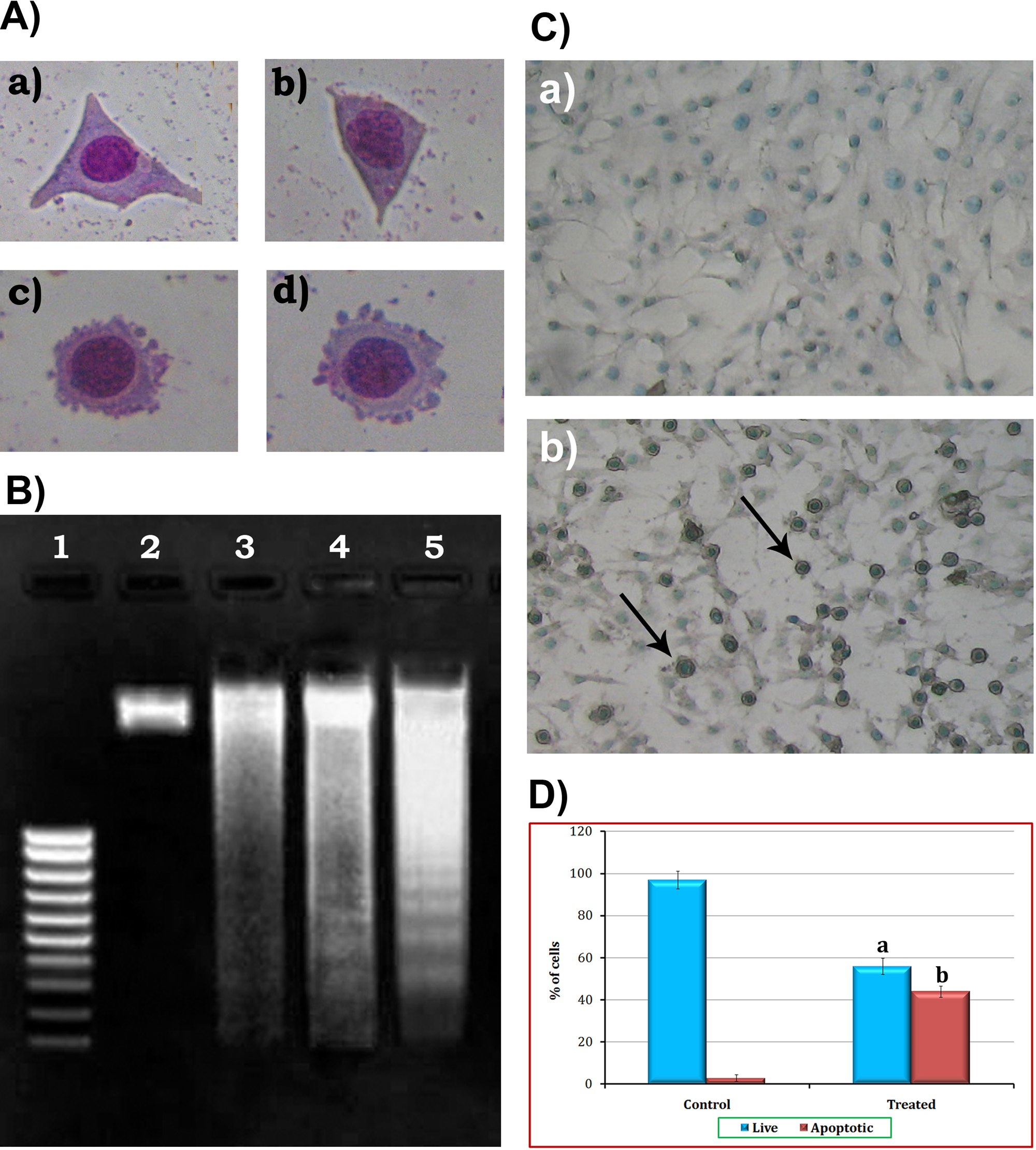
Effect of nomilin on apoptosis of B16F-10 cells: A. Morphology of B16F-10 melanoma cells: (a) untreated control after 48 hours of incubation, (b) treated with nomilin (5 µg/mL) after 48 hours of incubation, (c) treated with nomilin (10 µg/mL) after 48 hours of incubation, (d) treated with nomilin (25 µg/mL) after 48 hours of incubation. B. DNA fragmentation: lane 1, molecular weight marker; lane 2, DNA from untreated control cells; lane 3, DNA from nomilin-treated (5 µg/mL) cells; lane 4, DNA from nomilin-treated (10 µg/mL) cells; lane 5, DNA from nomilin-treated (25 µg/mL) cells. C. TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labeling) assay: (a) untreated control after 48 hours of incubation; (b) treated with nomilin (25 µg/mL) after 48 hours of incubation. D. Quantification of apoptotic cells. Values are mean ± standard deviation (mean of triplicate)
Discussion
Uncontrolled cell growth and resistance to apoptosis are major defects in cancer. The development of approaches that induce apoptotic machinery within cancer cells could be effective against their proliferation and invasive potential. 18 In the present study, we analyzed the effect and mechanism of action of nomilin on the inhibition of lung metastasis induced by B16F-10 melanoma cells. Invasion and metastasis are the most insidious and life-threatening aspects of cancer. Metastasis of cancer cells is a threat to available cancer therapy and is a major reason for treatment failure. Medicines available presently are cytoreductive drugs, which are nonspecific.
B16F-10 melanoma cells are highly metastatic and form colonies of tumor nodules in the lungs when administered through the tail vein, which in turn promote lung fibrosis and collagen deposition. The characteristic of this defect is an increased accumulation of extracellular matrix proteins in the alveolar walls, specifically collagen, resulting in reduction in pulmonary function. The index of lung fibrosis was evaluated by estimating the lung collagen hydroxyproline content. Nomilin treatment was effective in reducing lung fibrosis, which was confirmed by the lowered levels of lung collagen hydroxyproline content.
Sialic acid, a family of acetylated derivatives of neuraminic acid, occurs as the terminal component of carbohydrate chains of glycolipids and glycoproteins. Increased levels of β(1-6) branched N-linked oligosaccharides have been observed in many metastatic tumor cells, 19 and progression from a tumorigenic to metastatic phenotype in both rodent and human cancers has been associated with corresponding upregulation of sialic acid content. 20 Because uronic acid and hexosamines are the basic structures of sialic acids, 21 the index of tumor cell proliferation was also evaluated by estimating hexosamine and uronic acid contents. The lungs of the metastatic tumor–bearing animals showed higher levels of uronic acid and hexosamine. Administration of nomilin significantly (P < .05) reduced the elevated levels of uronic acid and hexosamine. Total sialic acid level in serum has been recognized as a valuable nonspecific monitor of tumor burden in melanomas. 22 The increased sialic acid levels in the control metastatic tumor–bearing animals were significantly (P < .05) reduced by the treatment with nomilin. Serum γGT is also a marker for neoplastic proliferation.23,24 Highly elevated levels of γGT are observed in patients with either primary or secondary neoplasms. The increased serum γGT level was reduced by treatment with nomilin. All these results correlated with the histopathological analysis and increase in life span of treated animals. The above experimental evidence shows that nomilin can inhibit the metastatic growth of B16F-10 melanoma cells in a mouse model.
Matrix metalloproteinases (MMPs) are a family of zinc-dependent endoproteinases that are capable of degrading almost all the components of the extracellular matrix and thereby upregulating invasion and metastasis.25,26 Among the MMPs reported earlier, MMP-2 and MMP-9 are key enzymes for degrading type IV collagen, which is a major component of the basement membrane.27,28 The result of zymographic analysis indicates that administration of nomilin inhibited the activation of MMPs. It also inhibited the invasion of B16F-10 melanoma cells through the collagen matrix. There is also strong evidence for the direct inhibition of activation of MMPs by nomilin.
To study the mechanism of action of antimetastatic activity, we have analyzed the ability of nomilin to suppress cell cycle progression and induce apoptosis in B16F-10 melanoma cells. Defective apoptosis is a major causative factor in the development and progression of cancer. 29 Cancer cells acquire resistance to apoptosis by overexpression of antiapoptotic proteins or downregulation or mutation of proapoptotic proteins. In the present study, morphological analysis of B16F-10 cells treated with nontoxic concentrations of nomilin showed the presence of membrane blebbing, cytoplasmic shrinkage, and nuclear condensation. A DNA ladder pattern in nomilin-treated cells also supports the above result.
Apoptosis is regulated by antiapoptotic and proapoptotic effectors, which involve a large number of biomolecules. The tumor suppressor gene p53 has come to be known as a master guardian of the genome and a member of the DNA damage-response pathway. p53 is able to induce growth arrest or apoptosis in DNA-damaged cells. Induction of G1 arrest depends on sequence-specific DNA binding and transcriptional activation of p53 target genes such as p21. p21 and p27 are CDK (cyclin-dependent kinases) inhibitors, shown to arrest the cell cycle and inhibit the growth of cancer cells.30,31 The members of the Bcl-2 family play an important role in the induction of apoptosis and are considered a target for anticancer therapy. Bcl-2 is an upstream effector molecule in the apoptotic pathway and is identified as a potent suppressor of apoptosis; its downregulation causes tumor regression. On the other hand, predominance of Bax, a proapoptotic protein, over Bcl-2 promotes apoptosis.32,33 Caspase-3, an essential member of the caspase family, can eventually induce cell apoptosis and be activated by upstream caspases, including caspase-9 and caspase-8. In this study, administration of nomilin could induce apoptosis by upregulating p53, Bax, caspase-9, caspase-3, p21, and p27 and also downregulating Bcl-2 gene expression.
Inhibition of deregulated cell cycle progression in cancer cells is an effective strategy to halt tumor growth. 34 Our results further demonstrate that treatment of B16F-10 cells with nomilin increases the peak of the sub-G1 phase (cells with fragmented DNA) of cell cycle progression, indicating that one of the mechanisms by which nomilin inhibits the proliferation and invasion of B16F-10 cells is the induction of apoptosis. Cyclin D1 serves as a key sensor and integrator of extracellular signals of cells in G0 and early G1 phases, mediating its function through binding with Cdk4/6. Aberrant cyclin D1 expression is a critical event in cancer development. 35 Treatment of nomilin downregulated the overexpression of cyclin D1 in B16F-10 cells.
The NF-κB family of transcription factors (Rel [c-Rel], Rel A [p65], Rel B, NF-κB-1 [p105/p50], NF-κB-2 [p100/p52]) is involved mainly in stress-induced immune and inflammatory responses. Chronic inflammation by constitutively active NF-κB has been shown to contribute to the development of many cancers. NF-κB can be activated by a variety of stimuli, including IL-1 (IL-1) and TNF-α, and among its multiple effects, it also induces antiapoptotic proteins and blocks caspase activation.36,37 Therefore, NF-κB was generally considered to act predominantly as a mediator of cell survival. In the present study, treatment with nomilin was found to inhibit the activation and nuclear translocation of transcription factors such as NF-κB p50, NF-κB p65, NF-κBc-Rel, c-Fos, ATF-2, and CREB in B16F-10 melanoma cells.
Cytokines and growth factors are critical regulators of immune responses, inflammatory reactions, cell proliferation, angiogenesis, and apoptosis; they play an important role in chronic diseases such as cancer.38-40 TNF, a proinflammatory cytokine that plays a major role in modulating cytokine cascades, upregulates the expression of several other cytokines such as GM-CSF, IL-1 (IL-1β), and IL-6. 41 Nomilin also inhibited the production and expression of proinflammatory cytokines such as IL-1β, TNF-α, IL-6, and GM-CSF in B16-F10 cells.
In summary, our study demonstrates that nomilin inhibits metastasis via induction of apoptosis in B16F-10 melanoma cells. Nomilin downregulated Bcl-2 and cyclin-D1 expression and upregulated p53, Bax, caspase-9, caspase-3, p21, and p27 gene expression in B16F-10 cells. Proinflammatory cytokine production and gene expression were found to be downregulated in nomilin-treated cells. The study also shows that nomilin could inhibit the activation and nuclear translocation of antiapoptotic transcription factors such as NF-κB, CREB, and ATF-2 in B16F-10 cells.
Footnotes
Acknowledgements
The authors sincerely thank Dr Ramadasan Kuttan, Research Director, Amala Cancer Research Centre, for his valuable suggestions.
The authors alone are responsible for the content and writing of the article.
The author(s) declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) received no financial support for the research and/or authorship of this article.