
Abstract
Objective:
Lysine-specific demethylase 2A (KDM2A), a specific H3K36me1/2 demethylase, has been reported to be closely associated with several types of cancer. In this study, we aimed to investigate the expression and function of KDM2A in colorectal adenocarcinoma.
Methods:
A total of 215 colorectal adenocarcinoma specimens were collected, and then subjected to immunohistochemistry assay to evaluate the expression levels of KDM2A, cyclin D1 and other proteins in colorectal adenocarcinoma tissues. Real-time polymerase chain reaction, Western blot, and other molecular biology methods were used to explore the role of KDM2A in colorectal adenocarcinoma cells.
Results:
In this study, we report that the expression level of KDM2A is high in colorectal adenocarcinoma tissues, and this high expression promotes the proliferation and colony formation of colorectal adenocarcinoma cells, as demonstrated by KDM2A knockdown experiments. In addition, the expression of KDM2A is closely associated with cyclin D1 expression in colorectal adenocarcinoma tissues and cell lines.
Conclusions:
Our study reveals a novel role for high-expressed KDM2A in colorectal adenocarcinoma cell growth, and that the expression of KDM2A is associated with that of cyclin D1 in colorectal adenocarcinoma.
Introduction
Colorectal adenocarcinoma represents a global public health threat, with over one million new cancer cases and over half a million cancer deaths occurring worldwide every year.1,2 The molecular mechanisms underlying the tumorigenesis and progression of colorectal adenocarcinoma remain largely unclear despite advances in clinical research and molecular biology over the last several decades. In addition, chemotherapy resistance has become a serious problem during colorectal adenocarcinoma treatment, and a number of patients show low response rates to current therapies. 2 Therefore, it is essential to elucidate the underlying molecular mechanisms of therapy resistance and to identify new biomarkers and therapeutic targets for colorectal adenocarcinoma diagnosis and treatment.
An overwhelming number of studies have proven that histone methylation is closely associated with cancer. Histone methylation is an important histone modification, and it plays critical roles in different biological processes, such as gene transcription and DNA damage response. 3 In general, methylation at H3K9, H3K27, and H4K20 correlates with transcriptional repression, while methylation at H3K4, H3K36, and H3K79 is linked to transcriptional activation.4-6 Histone methylation is tightly regulated to maintain a relatively stable state in normal cells, but it is dysregulated in cancer cells, resulting in transcriptional activation of oncogenes and repression of tumor suppressor genes, as well as genomic instability. 7
Histone methylation is reversible, which is mainly mediated by histone demethylases. Lysine-specific demethylase 2A (KDM2A) is a histone demethylase, which is capable of removing H3K36me1/2 from chromatin. 8 It has been demonstrated that KDM2A is involved in the development and progression of some types of cancer. For example, KDM2A was reported to be frequently overexpressed in non-small cell lung cancer (NSCLC) and required for the proliferation and invasion of NSCLC cells in vitro and in vivo. 9 In addition, low levels of KDM2A were found in prostate carcinomas, and KDM2A downregulation led to transcriptional activation of centromeric satellite repeats and unstable heterochromatin. 10 In our previous study, KDM2A was phosphorylated by protein kinase ATM in response to DNA double-strand breaks, and this phosphorylation abrogated KDM2A’s chromatin-binding capacity. Consequently, H3K36 dimethylation near DNA damage sites was significantly increased and served as a platform to recruit the MRE11 complex to DNA damage sites, indicating the critical role of KDM2A in DNA damage response. 11 However, the role of KDM2A in cancer development and progression, as well as the underlying molecular mechanisms of these events, is still largely unclear.
In this study, we present evidence that KDM2A is highly expressed in colorectal adenocarcinoma tissues and that KDM2A knockdown inhibits the proliferation and colony formation of colorectal adenocarcinoma cells. In addition, we found that the expression level of KDM2A is positively associated with that of cyclin D1 in colorectal adenocarcinoma. Overall, this study is the first to demonstrate that KDM2A is highly expressed in colorectal adenocarcinoma and is critical for colorectal adenocarcinoma cell growth, which may be because of its role in regulating cyclin D1 expression.
Materials and methods
Patients
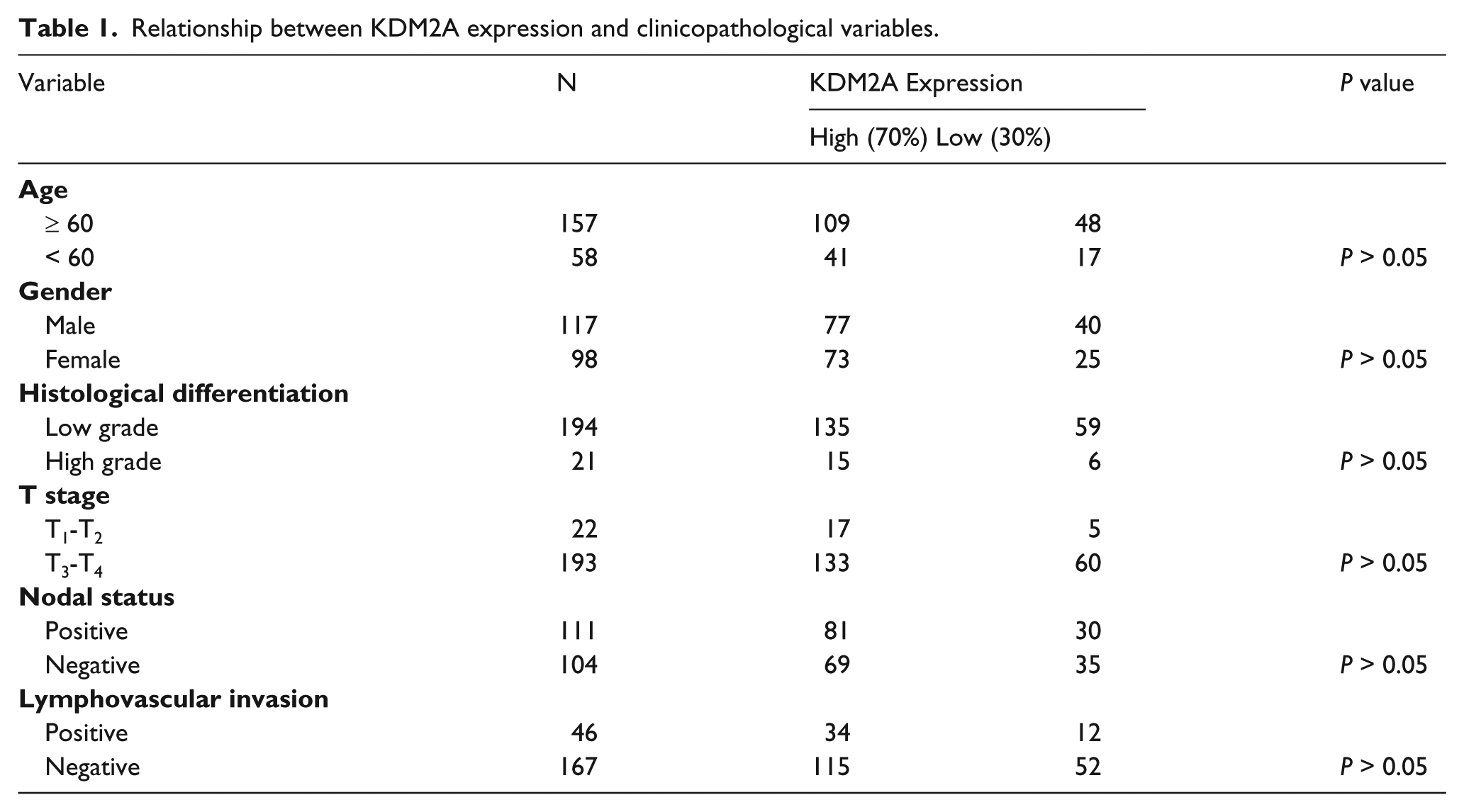
A total of 215 patients with colorectal adenocarcinoma treated at Peking University Cancer Hospital from 2004 to 2009 were included in this study. All 215 included patients underwent curative surgery. The patient demographic and clinicopathological characteristics are summarized in Table 1 and listed in Supplemental Table 1. This was a double-blind study, and approved by the Ethics Committee of our hospital, and all patients provided informed consent to agree to donate their tumor tissues for research.
Relationship between KDM2A expression and clinicopathological variables.
Cell lines and antibodies
HCT116 and LoVo cell lines were purchased from the American Type Culture Collection and grown in McCoy’s 5A medium or DMEM medium supplemented with 10% fetal bovine serum (Biological Industries) in a 37°C incubator with a humidified, 5% CO2 atmosphere. The antibodies used were anti-KDM2A (ABclonal Technology and Bethyl), anti-cyclin D1 (Abcam), anti-β-actin (ZSGB-BIO), anti-p27 (Abcam), anti-p53 (Invitrogen), anti-CD44 (Abcam) and anti-p170 (Sigma-Aldrich).
Immunohistochemistry
All specimens were immunostained for KDM2A, p53, p27, CD44, p170 and cyclin D1. Sections 4 μm in thickness were cut from the formalin-fixed, paraffin-embedded tumor tissues and mounted on cationic slides. After dewaxing and rehydration, sections for immunostaining were subjected to heat antigen retrieval in a citrate buffer solution (pH 6.0) and heated for three min (103 kPa, 120°C). After cooling, 0.5% hydrogen peroxide was used to block the activation of endogenous peroxidase, and sections were then rinsed with phosphate-buffered saline (PBS) five times. Next, the sections were incubated at 37°C with primary antibody for 1 h. After rinsing in PBS, sections were incubated with a biotinylated secondary antibody for 30 min at 37°C and with avidin-biotin prediluted biotinylated complex for 10 min at room temperature. Antigenic sites were revealed by adding diaminobenzidine solution. Finally, the sections were washed with distilled water, counterstained lightly with hematoxylin, then dehydrated and mounted. The results were evaluated by a senior pathologist, who was blinded to the patient clinical data. The immunostaining intensity of KDM2A, p53, p27, CD44, p170, and cyclin D1 was classified as previously reported. 12 The patients were categorized as having high expression of a marker if >50% of cells had strong staining; otherwise, they were considered to have low expression.
RNA interference
The sequences of the small interfering RNA (siRNA) were as follows: negative control (NC), which was a non-specific siRNA, UUCUCCGAACGUGUCACGU; KDM2A siRNA1, AACAAGGAGAGUGUGGUGUUU, KDM2A siRNA2, GAACCCGAAGAAGAAAGGAUUCG. All siRNA oligonucleotides were purchased from Shanghai GenePharma (Shanghai, China). The siRNAs were transfected into cells with Lipofectamine 2000 (Life Technologies-Invitrogen) and incubated for 48 hours according to the manufacturer’s instructions.
RNA extraction and real-time polymerase chain reaction
The cells transfected with NC or KDM2A siRNA were collected, and RNA was extracted from each using TRIzol (Invitrogen) according to the manufacturer’s instructions. RNA was reverse-transcribed to cDNA by a Quantscript RT Kit (Tian-gen). Then, the cDNA was amplified by a SYBR Green qPCR Kit (Tian-gen) and an ABI7500 instrument (Life Technologies). The primers used for amplifying KDM2A and GAPDH were as follows: KDM2A-F, ATGGCTCGTCAATAGGCT; KDM2A-R, GGACTGATCTGTAAGGTGGC; GAPDH-F, AACGGATTTGGTCGTATTGGG; GAPDH-R, CCTGGAAGATGGTGATGGGATT.
Colony formation assay
The colony formation assay protocol has been previously described. 11 Briefly, cells were transfected with either NC or KDM2A siRNA. Forty-eight hours later, cells were trypsinized and counted, and an equal number was plated into six-well plates. After several days, methanol fixation and staining with methylene blue was performed to identify visible colonies.
Cell proliferation assay
The cell proliferation assay has been previously described in another study, 13 and it is comparable to a traditional MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide) assay. Briefly, LoVo and HCT116 cells were transfected with either NC or KDM2A siRNA. Then, the cell confluences were determined by a Genetix CloneSelect Imager every 24 hours. Each experiment was performed three times, and growth curves were generated using Microsoft Excel.
Western blot
Total protein was extracted from colorectal adenocarcinoma tissues and cells with lysis buffer containing 50 mM Tris-HCl, 250 mM NaCl, 5 mM EDTA, 50 mM NaF, 0.1% NP40, and 1% protease inhibitor cocktail. Equal amounts of protein were size fractionated by SDS-PAGE and then incubated with primary antibodies and secondary antibodies, sequentially.
Statistical analysis
For all statistical tests, P<0.05 by a chi-squared test, Fisher’s exact test, or Student’s t-test, as appropriate, was considered statistically significant.
Results
KDM2A was highly expressed in colorectal adenocarcinoma tissues
As KDM2A was dysregulated in NSCLC and prostate carcinomas, we wanted to determine whether abnormal expression of KDM2A occurred in colorectal adenocarcinoma. At first, 215 patients were included to assess KDM2A expression in colorectal adenocarcinoma tissues. Immunohistochemistry (IHC) analysis indicated that KDM2A was highly expressed in 70% of cancer tissues (Figure 1(a), Table 1), and a representative IHC staining pattern indicating high KDM2A expression is shown in Figure 1(b). No significant association between KDM2A expression and clinicopathological variables was observed (Table 1), which may be because the number of enrolled patients is small.
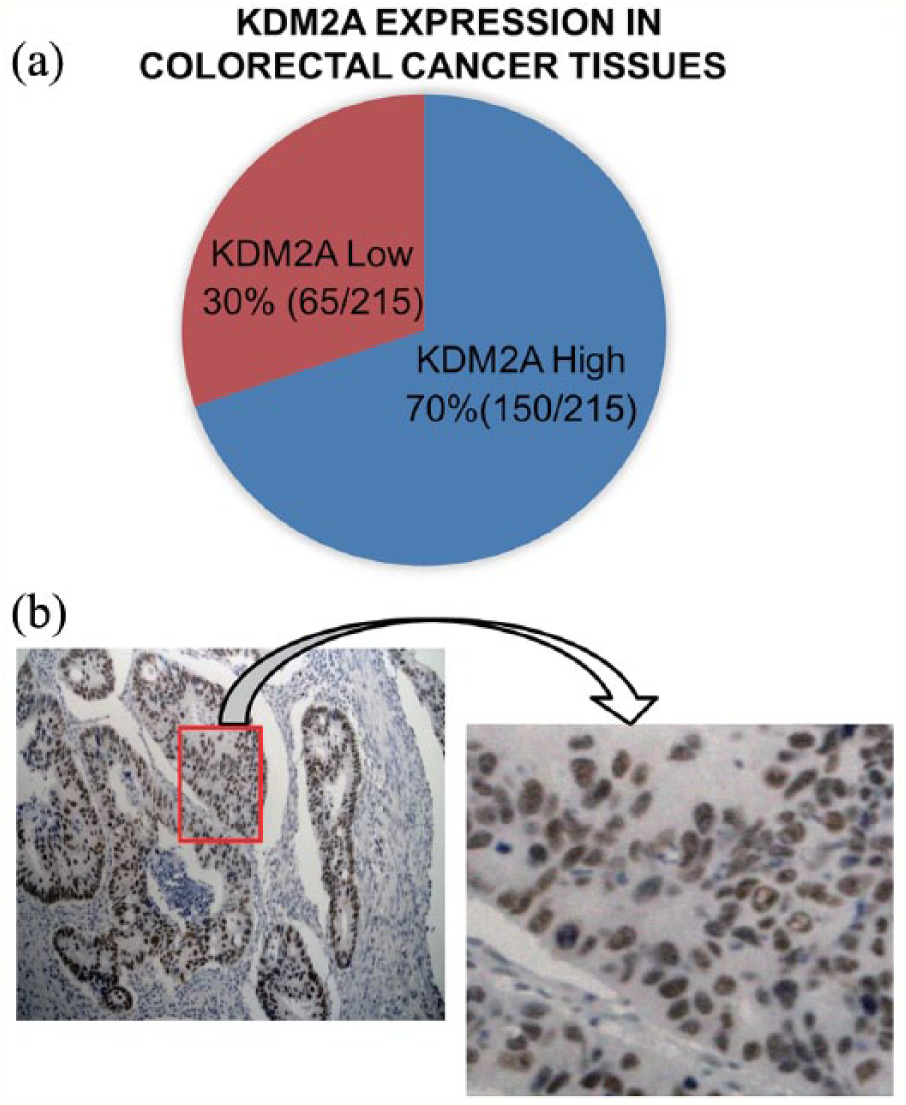
High expression of KDM2A in colorectal adenocarcinoma tissues. (a) The pie diagram showing the expression level of KDM2A in 215 colorectal adenocarcinoma tissues. (b) Left, a representative IHC staining pattern of KDM2A in primary colorectal adenocarcinoma; right, magnification of the square in the left image.
KDM2A knockdown inhibits the proliferative and colony formation abilities of colorectal adenocarcinoma cells
As KDM2A was highly expressed in colorectal adenocarcinoma, it was hypothesized that KDM2A plays a critical role in colorectal adenocarcinoma development and progression. To investigate the function of KDM2A, RNA interference was used to knock down KDM2A in colorectal adenocarcinoma cells. As shown in Figure 2 and Supplemental Figure 1, the mRNA and protein levels of KDM2A were markedly downregulated in HCT116 or LoVo cells transfected with KDM2A siRNA compared with control cells. The cell confluency was determined every day after transfection with NC or KDM2A siRNA2, and we found that the proliferative ability of both HCT116 and LoVo cells was inhibited upon KDM2A knockdown (Figure 3(a) and (b)). To further investigate the influence of KDM2A knockdown on the colony formation ability of colorectal adenocarcinoma cells, colony formation assays were performed. As shown in Figure 3(c) and (d), the number of colonies formed by KDM2A knockdown HCT116 or LoVo cells was less than that by control cells, indicating that KDM2A knockdown inhibited the colony formation ability of colorectal adenocarcinoma cells.
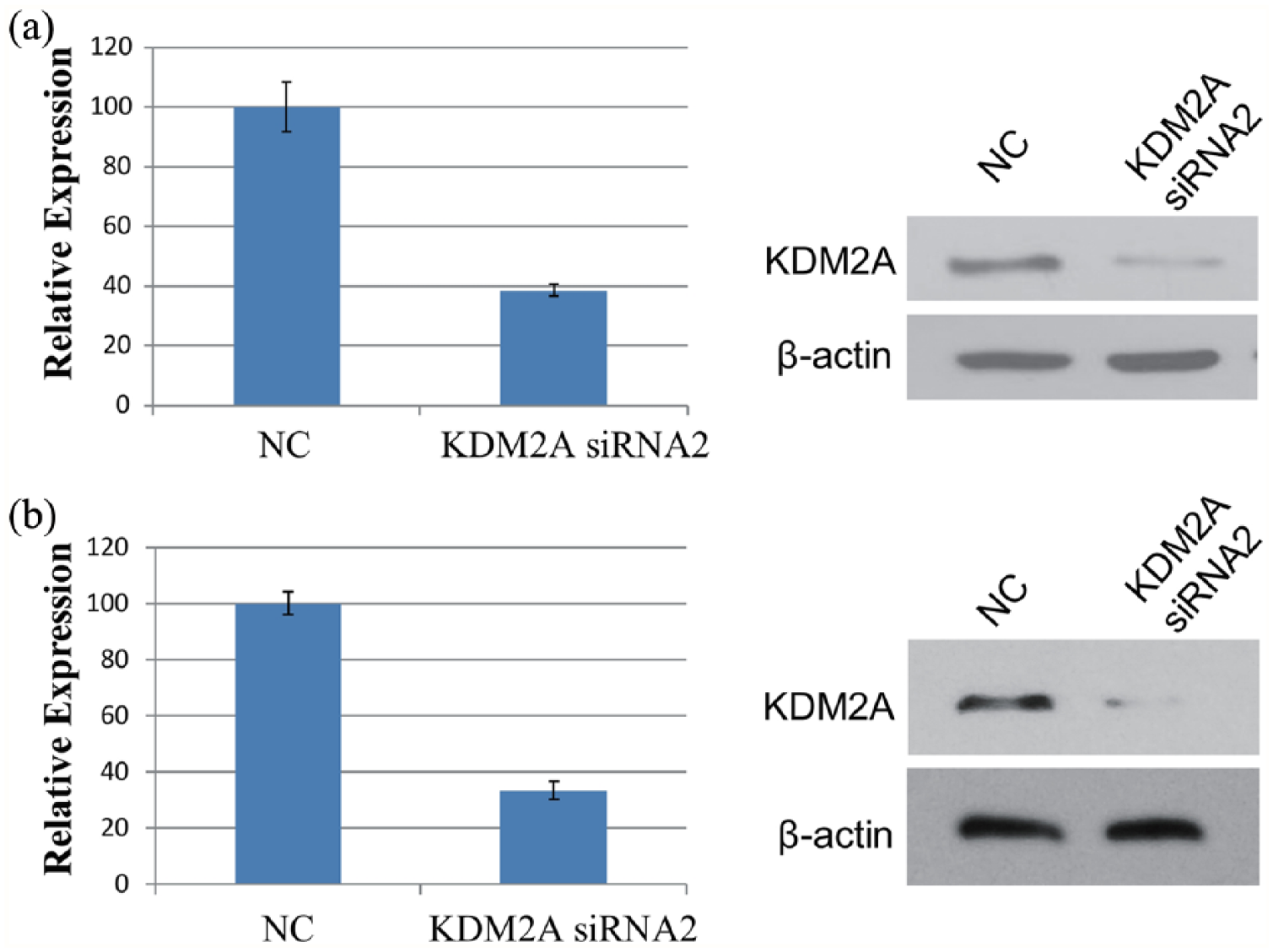
KDM2A knockdown in HCT116 and LoVo cells. (a) HCT116 cells were transfected with NC or KDM2A siRNA2. At 48 hours after transfection, the expression of KDM2A mRNA was determined by real-time PCR (left), and the protein level of KDM2A was detected by western blot (right). (b) LoVo cells were transfected with NC or KDM2A siRNA2. At 48 hours after transfection, the expression of KDM2A mRNA was determined by real-time PCR (left), and the protein level of KDM2A was detected by Western blot (right).
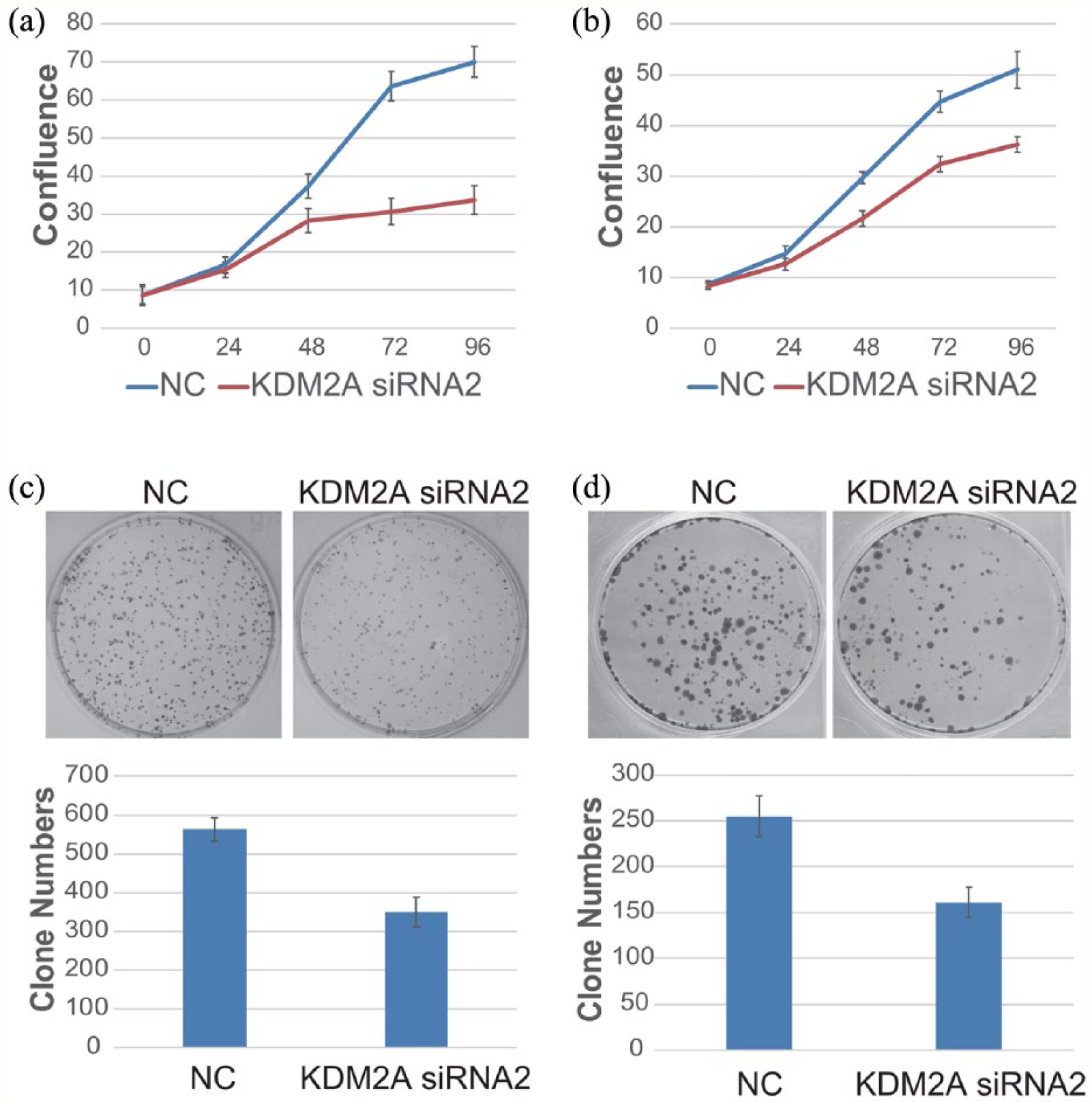
Inhibition of the proliferative and colony formation abilities of colorectal adenocarcinoma cells by KDM2A knockdown. (a) and (b) HCT116 cells (a), as well as LoVo cells (b), were transfected with NC or KDM2A siRNA2. Cell confluency was detected by a Genetix CloneSelect Imager every 24 hours. (c) HCT116 cells were transfected with NC or KDM2A siRNA2. Then, cells were trypsinized and equally plated into six-well plates. About 10 days later, the cell colonies were stained and counted. Data are shown as the means ± SD from three independent experiments. (d) LoVo cells were transfected, trypsinized and plated as in C. Approximately 14 days later, the cell colonies were stained and counted. Data are shown as the means ± SD from three independent experiments. Student’s t-test (two-tailed): NC versus KDM2A siRNA2 (HCT116), P<0.05; NC versus KDM2A siRNA2 (LoVo), P<0.05.
KDM2A expression is associated with cyclin D1 expression in colorectal adenocarcinoma tissues
Subsequently, we determined the molecular mechanism by which KDM2A affected the proliferative and colony formation abilities of colorectal adenocarcinoma cells. As protein tumor suppressor p53, 14 cyclin-dependent kinase inhibitor 1B (p27), 15 epidermal growth factor receptor (p170), 16 cyclin D1, 17 and CD44 18 are all critical regulators of cell growth in colorectal adenocarcinoma, we first determined the expression levels of these proteins by IHC assays in 215 colorectal adenocarcinoma tissues (Supplemental Table 1). Then, we investigated the association of KDM2A with each of these tumor growth-related proteins separately. As shown in Figure 4, there was no significant association of KDM2A expression with expression of p53, p27, p170, or CD44 in colorectal adenocarcinoma tissues. Interestingly, we found that KDM2A expression was positively associated with cyclin D1 expression in vivo (Figure 4(d)). In addition, although KDM2A was highly expressed in 70% (150/215) of colorectal adenocarcinoma tissues, there were still 30% (65/215) cancer tissues with low-expressed KDM2A (Table 1). We found that cyclin D1 was low expressed in 72.3% (47/65) cancer tissues with low-expressed KDM2A, and it was further verified by IHC double staining of both KDM2A and cyclin D1 (Supplemental Figure 2).
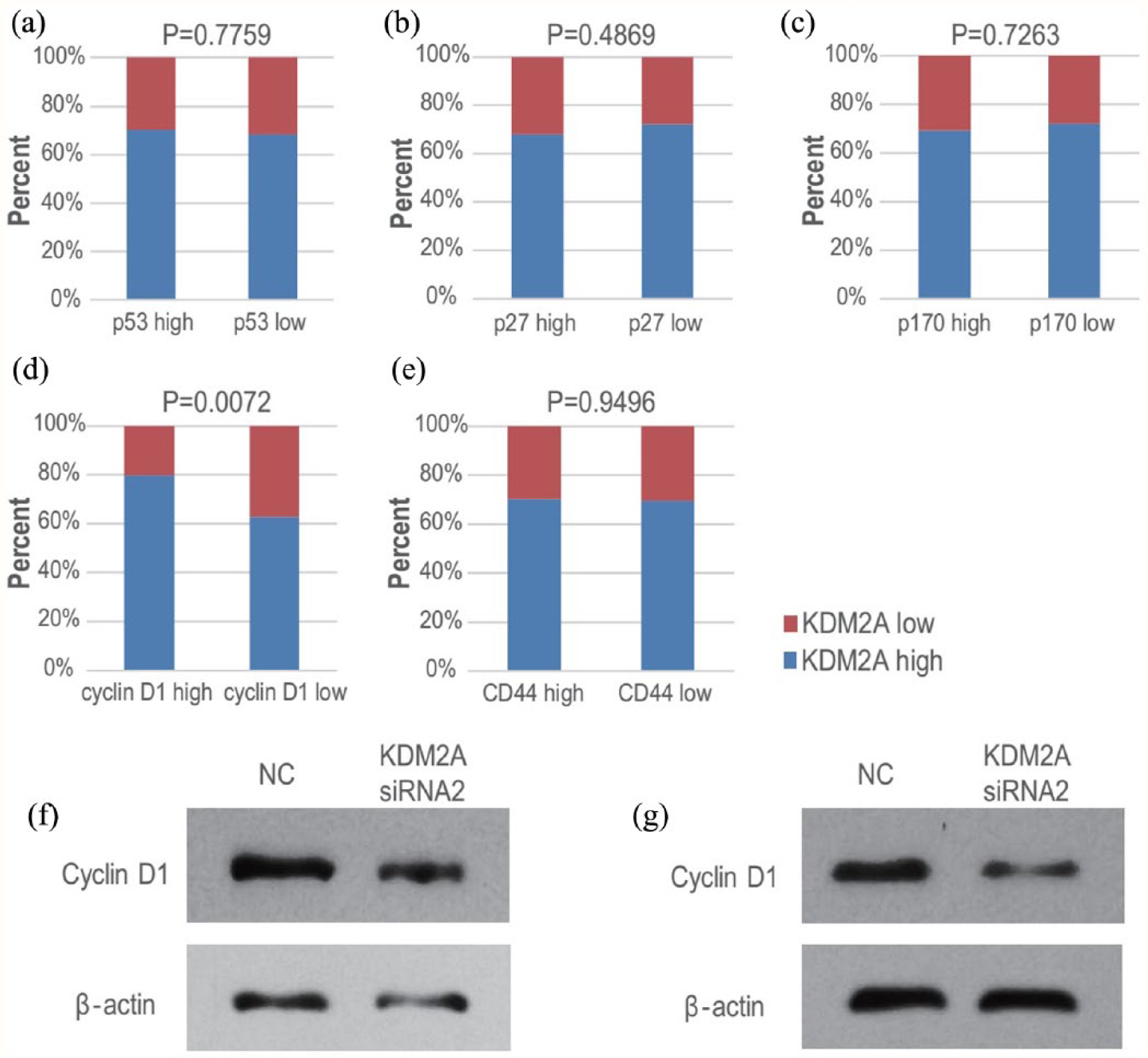
The expression of KDM2A was associated with that of cyclin D1 in colorectal adenocarcinoma tissues and cell lines. (a) to (e) The expression of p53, p27, p170, cyclin D1, and CD44 was determined by IHC in 215 colorectal adenocarcinoma tissues, and then the association analysis between KDM2A and p53 (a), p27 (b), p170 (c), cyclin D1 (d) or CD44 (e) was performed. (f) to (g) HCT116 cells (f), as well as LoVo cells (g), were transfected with NC or KDM2A siRNA2 separately. Then the cell extracts were subjected to Western blot.
As KDM2A expression positively correlated with cyclin D1 expression in colorectal adenocarcinoma tissues, and KDM2A is a regulator of gene expression as a histone demethylase, we next investigated whether KDM2A regulated cyclin D1 expression in colorectal adenocarcinoma cells. KDM2A was knockdown in HCT116 and LoVo cells, and the protein levels of cyclin D1 were detected. As shown in Figure 4(f) to (g), cyclin D1 protein levels were both significantly downregulated upon KDM2A knockdown in HCT116 and LoVo cells, suggesting that KDM2A regulates cyclin D1 expression and is associated with cyclin D1 in colorectal adenocarcinoma cells.
Discussion
In this study, KDM2A was identified as a novel modulator of tumor growth in colorectal adenocarcinoma. KDM2A was highly expressed in colorectal adenocarcinoma tissues, and the proliferative and colony formation abilities of colorectal adenocarcinoma cells were inhibited upon KDM2A knockdown. In addition, we found that KDM2A expression was positively associated with cyclin D1 expression in colorectal adenocarcinoma tissues and cell lines.
Recently, it was reported that histone methylation has a critical role in tumor development and progression. For example, H3K27 methyltransferase enhancer of zeste homolog 2 (EZH2) is overexpressed in hormone-refractory, metastatic prostate cancer, and prostate cancers that express higher EZH2 show a poorer prognosis. 19 Another study showed that EZH2 overexpression in prostate cancer cells induces transcriptional repression of a specific cohort of genes, including ADRB2 (adrenergic receptor, beta-2), a critical mediator of beta-adrenergic signaling. 20 Previous studies have also demonstrated that H3K36 methylation plays an important role in cancer. Defects in several H3K36 methyltransferase genes, such as nuclear receptor SET domain-containing 1 (NSD1), NSD2, NSD3, and SET domain-containing 2 (SETD2), have been implicated in multiple cancer types.21-26 In addition, KDM2A and lysine-specific demethylase 2B (KDM2B), the H3K36 demethylases, were also reported to be involved in cancer initiation, maintenance, or progression.9,27-29 Therefore, aberrant H3K36 methylation, as well as dysregulated H3K36 methyltransferases and demethylases, are critical for tumor development and progression. Our study contributes evidence that KDM2A is critical for the proliferation of colorectal adenocarcinoma cells.
KDM2A is often dysregulated in cancer, but the role of KDM2A in different cancer types varies. In some types of cancer, KDM2A acts as a tumor suppressor gene to inhibit cancer progression. For example, it has been reported that KDM2A associates with centromeres and represses the transcription of small non-coding RNAs that are encoded by the clusters of satellite repeats at the centromere, and that decreased KDM2A in prostate carcinomas compromises the heterochromatic state and induces genomic instability. 10 However, KDM2A seems to be an oncogene in some other cancer types. For example, KDM2A was reported to be increased in gastric cancer tissues and to facilitate gastric cancer cell growth and migration through the downregulation of programmed cell death 4 (PDCD4), a known tumor suppressor in the progression of gastric cancer. 29 In addition, KDM2A promotes stemness and angiogenesis in breast cancer by upregulating two pro-angiogenic factors, Jagged 1 and platelet-derived growth factor subunit A (PDGFA). 30 Similarly, this study reports that KDM2A may serve as an oncogene to promote colorectal adenocarcinoma cell growth.
This study has several limitations. First, the patients included were only from one hospital in Beijing, China, and the number was small. It is necessary to conduct a multi-center study with much greater number of enrolled patients to further evaluate the value of KDM2A in colorectal adenocarcinoma diagnosis and prognosis. In addition, the molecular mechanism by which KDM2A regulated colorectal adenocarcinoma cell growth is not clear yet.
In summary, our data indicate that KDM2A might be a new regulator of colorectal adenocarcinoma. To our knowledge, this is the first study to show a role for high-expressed KDM2A in colorectal adenocarcinoma cell growth. This study may provide a useful strategy by which to diagnose and cure colorectal adenocarcinoma by targeting KDM2A.
Supplemental Material
Supplement_Fig1 – Supplemental material for Lysine-specific demethylase 2A expression is associated with cell growth and cyclin D1 expression in colorectal adenocarcinoma
Supplemental material, Supplement_Fig1 for Lysine-specific demethylase 2A expression is associated with cell growth and cyclin D1 expression in colorectal adenocarcinoma by Lin-Lin Cao, Changzheng Du, Hangqi Liu, Lin Pei, Li Qin, Mei Jia and Hui Wang in The International Journal of Biological Markers
Supplemental Material
Supplement_Fig2 – Supplemental material for Lysine-specific demethylase 2A expression is associated with cell growth and cyclin D1 expression in colorectal adenocarcinoma
Supplemental material, Supplement_Fig2 for Lysine-specific demethylase 2A expression is associated with cell growth and cyclin D1 expression in colorectal adenocarcinoma by Lin-Lin Cao, Changzheng Du, Hangqi Liu, Lin Pei, Li Qin, Mei Jia and Hui Wang in The International Journal of Biological Markers
Supplemental Material
Supplement_Tab1 – Supplemental material for Lysine-specific demethylase 2A expression is associated with cell growth and cyclin D1 expression in colorectal adenocarcinoma
Supplemental material, Supplement_Tab1 for Lysine-specific demethylase 2A expression is associated with cell growth and cyclin D1 expression in colorectal adenocarcinoma by Lin-Lin Cao, Changzheng Du, Hangqi Liu, Lin Pei, Li Qin, Mei Jia and Hui Wang in The International Journal of Biological Markers
Footnotes
Author contributions
Lin-Lin Cao and Changzheng Du contributed equally to this study.
Declaration of conflicting interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Beijing Natural Science Foundation grant 7164305 and National Natural Science Foundation of China grant 81702788.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.