
Abstract
In this study, we quantified intergenerational, demographic variability of Moina macrocopa subjected to cadmium stress. Exposure of M macrocopa to cadmium (0.2, 0.3, and 0.4 mg/L as CdCl2) through 4 consecutive generations revealed changes in demographic responses not only in survivorship variables but also in reproductive parameters. Long-term demographic responses varied differently, depending on the demographic trait and the concentration of heavy metal in the medium. With the exception of generation time, all life history traits were significantly and adversely influenced due to increase in Cd concentrations. The average life span of M macrocopa varied up to 40% depending on Cd level and the generation of exposure. The highest gross reproductive rates were recorded in controls, while the lowest (∼30% less) were recorded at the highest Cd level. Survival-weighted net reproductive rates were reduced by nearly 50% due to Cd toxicity. The rate of population increase per day of M macrocopa was also significantly affected (∼25%) by Cd as compared to controls. This cladoceran showed a dose–response to Cd toxicity with a significance in both magnitude and frequency of offspring production.
Introduction
Most aquatic ecosystems become contaminated with heavy metals due to their release from domestic, industrial, and other anthropogenic activities. This creates stressful conditions that restrict normal life responses, for example, development and reproduction of aquatic organisms, including plankton. 1 Mercury and cadmium are the 2 heavy metals commonly found in water bodies around the world. In Mexico, Cd is more abundant than Hg. 2 This heavy metal is a ubiquitous environmental contaminant and highly toxic to zooplankton. 3 It has no known biological role and induces toxicity and stress symptoms. 4,5 Cd ions elicit a diverse range of cellular responses that modulate life history characteristics of aquatic organisms. 6,7 Cd is also a potent calcium antagonist disrupting calcium uptake, inducing oxidative stress, and affecting lipid peroxidation. The result of Cd toxicity is severe damage to many systems, membranes, and tissues. 5
For zooplankton species, the persistence depends on life history strategies that allow them to survive through unfavorable and stressful conditions. 8 Ability to survive stress is determinant in an organism’s fitness. This capacity depends, in part, on the genotypes of the species, but abiotic factors also influence the outcome. 9 In addition to the environmental conditions currently experienced by an organism, conditions experienced by previous generations can shape this outcome. Intraspecific variations may be studied within and across populations or generations. 10 The latter approach offers a research advantage because organisms exposed to stress can exert strong maternal influence on the life history traits of their offspring. 11,12 Thus, these transgenerational effects are of fundamental importance for species to survive in stressful environments. 13
Most demographic studies have been conducted within a single generation (horizontal transfer effects) in ecological communities. 14 Essentially, these studies have shown that the concentrations of heavy metals or other pollutants decrease the body size, life span, somatic and population growth rates, and delay the onset of maturity. 15 However, these studies have considered only a few taxa and then extrapolated to the future consequences based on an organism’s current responses. 16 -18 This approach ignores potential negative effects of toxicant exposure from parental to successive generations. 19,20 Research focusing on the effect of chronic exposure to Cd on demographic processes across generations is scarce and needs further investigation. 21 However, the available data are limited, suggesting that the standard cladoceran reproduction test be extended for multigenerational experiments to cover effects on progeny fitness. 22,23
Considering that sublethal toxicity often correlates with a reduction in many life table responses including reproduction-related parameters, it would be important to know how cadmium levels modulate life history traits of M macrocopa, through consecutive generations exposed to this metal. Thus, the aim of this study was to quantify the intra- and intergenerational demographic variability of this cladoceran. Consequently, our aim was to evaluate across generations the vertical transfer of effects driven by sublethal cadmium stress on the demography and fecundity of M macrocopa. To test the toxic effects and stress resistance, we hypothesized that low to moderate concentrations of Cd in the range normally found in most aquatic ecosystems increase the life span and stress resistance of M. macrocopa due to transgenerational effects.
Material and Methods
Isolation and Mass Culture of Cladocerans
Moina macrocopa was originally isolated from a permanent water body in Mexico City, (Mexico). Mass culture of clonal population of M macrocopa was established under laboratory conditions starting with a single parthenogenetic female. The cladocerans in mass cultures were fed single-celled green alga Chlorella vulgaris (strain CL V-3 registered in the CICESE, Ensenada, Mexico) at a density of about 1 × 10 6 cells/mL, and the medium was changed every alternate day. Cladocerans were cultured on moderately hard water (Environmental Protection Agency [EPA] medium), which consisted of 96 mg NaHCO3, 60 mg CaSO4, 60 mg MgSO4, and 4 mg KCl dissolved in 1 L of distilled water. 24 Chlorella was mass cultured in 2 L transparent bottles using Bold’s basal medium. 25 Log-phase algae was harvested, centrifuged at 3000 rpm, rinsed, resuspended in a small volume (5 mL) of distilled water, and stored in a refrigerator until use. The density of the concentrated alga was estimated using a hemocytometer. The desired algal density was obtained through dilution using EPA medium.
From the mass cladoceran culture, we carefully separated gravid females that were grown in isolation to obtain sufficient virgin neonates (<12 hours old), which were used in the experiments. The test conditions that favored optimal survival and reproduction of Moina were photoperiod 16 to 8 hours light:dark cycle, at 25°C ± 2°C, and pH at 7.2.
Multigeneration Experimental Setup
The sublethal cadmium levels were chosen based on previous reports in the literature. 7 Using analytical grade CdCl2, we prepared a nominal stock solution at a concentration of 50 mg/L. From the stock solution, the desired concentrations (0.2, 0.3, and 0.4 mg/L) were prepared using EPA medium. The test design consisted of 3 sublethal cadmium levels plus controls (0.0 mg/L of cadmium) and for 4 consecutive generations (F0, F1, F2, and F3) of M macrocopa. The neonates of F0 generation came from the mass cultures, while the neonates of F1, F2, and F3 generations were obtained from the second broods of test population treatments in controls or exposed to respective Cd concentrations.
Thus, we conducted the life table experiments for all 4 consecutive generations under continuous exposure to sublethal Cd concentrations. For any given generation, the life table tests were initiated with a cohort of 20 neonates (<12 hours old) present in transparent jars containing 50 mL of EPA medium, 0.75 × 106 cells/mL of C vulgaris, and the chosen cadmium levels plus controls. For each treatment, we set up 4 replicate cohorts. Thus, for each generation, we used 16 test jars (3 cadmium levels × 4 replicates + 4 controls). Following initiation of experiment, daily we recorded the number of surviving individuals from each cohort and the number of offspring born, if any. Surviving individuals of the original cohort were transferred to fresh test jars containing appropriate treatment, and offspring were discarded daily. However, the offspring produced during the second and third broods (and rarely the fourth) under a given treatment were used as the starter population for the next-generation life table experiments. The experiments were terminated when every individual in each cohort of all treatments had died.
For each generation, based on the survivorship and reproduction data collected, we derived life expectancy at birth, gross reproductive rate, net reproductive rate, generation time, and the rate of population increase (r) using the following formulae. 26
lx = Proportion of surviving to start of age x
mx = Offspring produced per female at age x
Life expectancy at the start of age x:
where Tx = cumulative number of individuals for further live from age x
nx = number of living individuals at the initiation of age x (days)
Gross reproductive rate
Net reproductive rate
Generation time:
For the rate of population increase, we iteratively resolved the Euler-Lotka equation:

To test whether the differences among the treatments were significant, we used 2-way analysis of variance and post hoc (Tukey) tests after satisfying the conditions of normality of data and homogeneity of variance. 27
Results
Exposure of M macrocopa to cadmium levels through the 4 consecutive generations revealed changes in demographic responses not only in survivorship variables but also in reproductive parameters. At any given generation, increase in the Cd concentration resulted in decreased survivorship. This effect was more severe with increasing generations. For example, at F0 generation, the age-specific survivorship curves were nearly rectangular, even at a Cd concentration of 0.4 mg/L. On the other hand, survivorship curves showed steep mortality after fifth day at 0.4 mg/L of Cd level at F1 and F2 generations. In the absence of Cd, survivorships had nearly the same pattern of mortality except for F3 where the population of Moina had slightly higher mortality in the initial days (Figure 1).

Age-specific survivorship (lx) of Moina macrocopa exposed to different concentrations of cadmium through different generations. Shown are the mean ± standard error based on 4 replicates (cohorts).
The age-specific fecundity curves across the generations and treatments with and without Cd showed continuous reproduction until death of the population. However, there was a strong reduction in the magnitude of offspring production in treatments exposed to Cd. At any given Cd concentration, reduction in offspring was stronger with increasing generations. Similarly, at any given generation, increasing Cd had adverse effects on the cladoceran reproduction. With increase in Cd level and generation, there was also a delay in the day at first reproduction, especially in low and medium heavy metal levels. For example, in controls, M macrocopa began to reproduce within 2 days, while in Cd treatments from F2 and F3 generations, the animals needed nearly double this duration for the age at first reproduction (Figure 2).
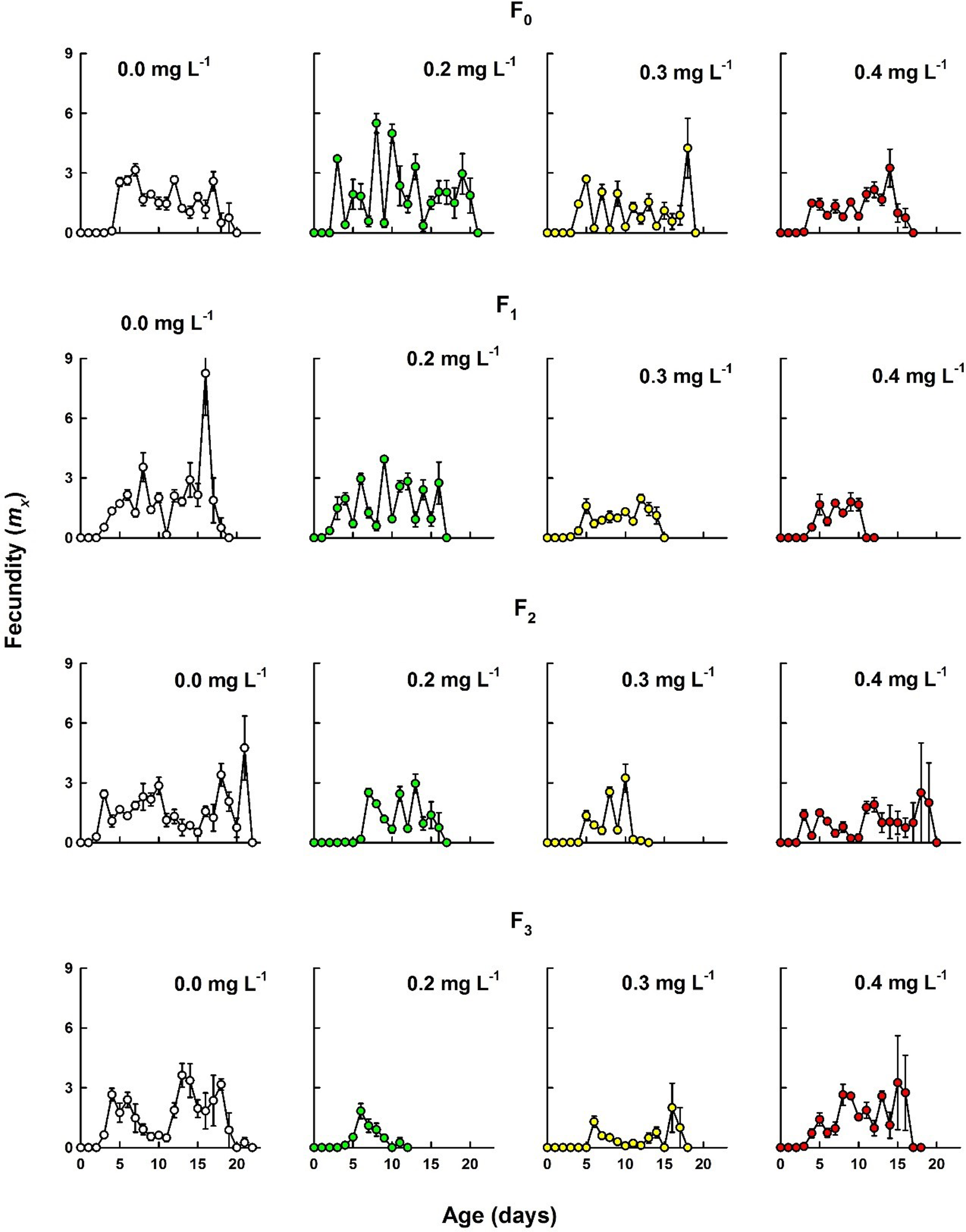
Age-specific fecundity (mx) of Moina macrocopa exposed to different concentrations of cadmium through different generations. Shown are the mean ± standard error based on 4 replicates (cohorts).
Long-term demographic responses of M macrocopa exposed to sublethal cadmium concentrations varied, depending to the demographic trait and the heavy metal concentration in the medium (Table 1). In general, with the exception of generation time, all life history metrics were significantly and adversely influenced due to increase in the Cd concentrations. Average life span of M macrocopa varied from 8 to 14 days depending on Cd level and the generation of exposure. High gross reproductive rates (>30 offspring/female/life span) were recorded in controls, while in the highest Cd level these were reduced to nearly one-third. Survival-weighted net reproductive rates were reduced ca 50% due to Cd toxicity. Rate of population increase per day of M macrocopa was also significantly (P < .05, Tukey test) affected (about 25%) by Cd as compared to controls.
Intergenerational (F0,…, F3) Variations in the Life History Parameters of Moina macrocopa Exposed to Different Concentrations of Cadmium.a
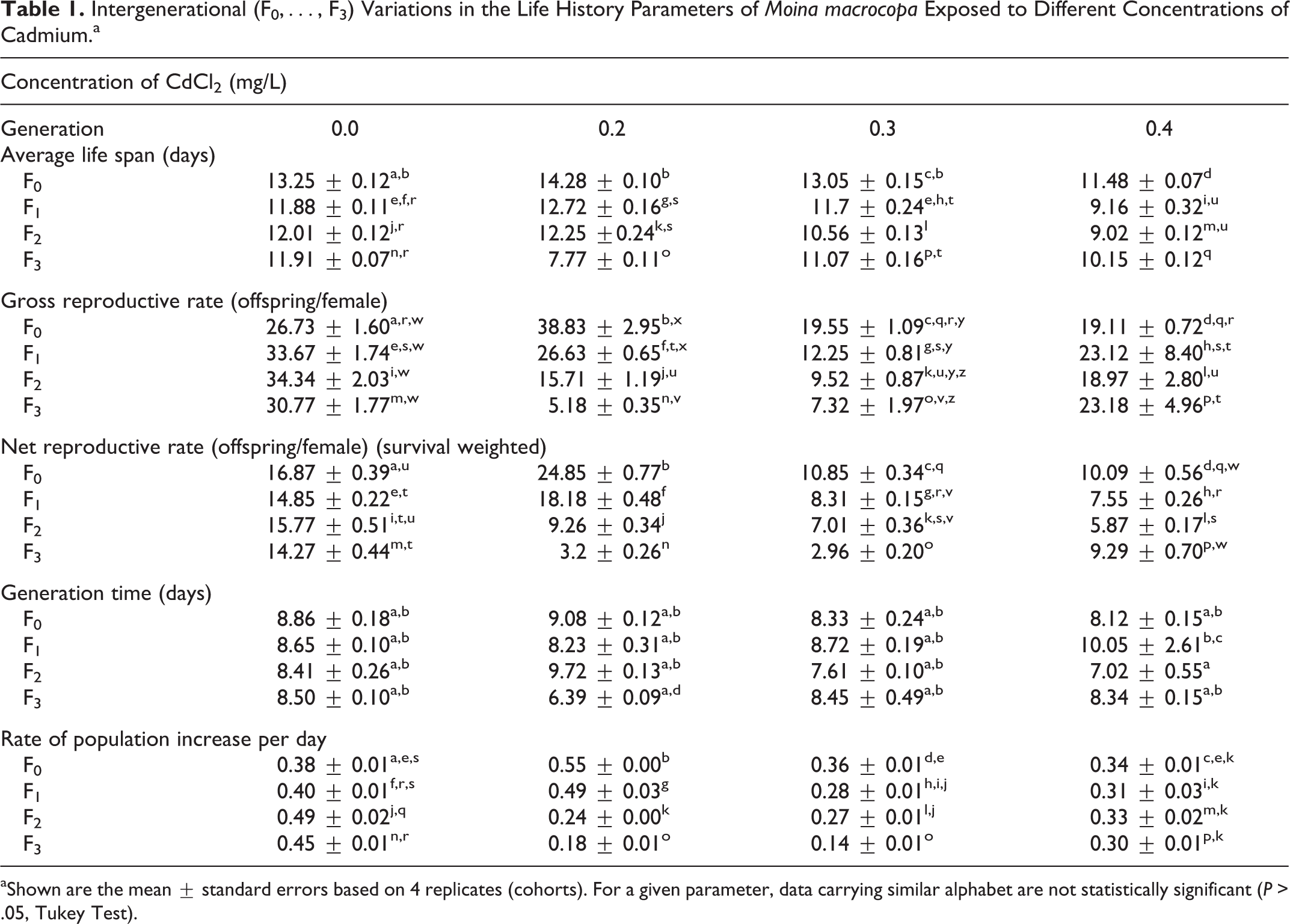
aShown are the mean ± standard errors based on 4 replicates (cohorts). For a given parameter, data carrying similar alphabet are not statistically significant (P > .05, Tukey Test).
Discussion
Heavy metals such as Cd adversely affect life history variables of cladocerans. Cadmium at concentrations as low as 0.2 mg/L reduce both life span and reproduction of zooplankton species. Gama-Flores et al 28 have exposed rotifers to Cd at concentrations of 0.05 to 0.2 mg/L and quantified the survival and reproductive rates. They found that at the highest Cd level tested, survival was reduced by 10% and reproductive output was decreased by ∼20% as compared to controls. In our study, increase in Cd level had deleterious effects across generations on most of the life history variables of Moina. Generally, the demographic studies on cladocerans subjected to toxicant stress are conducted in 1 generation, and rarely multigenerational approach is attempted. 29 On the other hand, it is well known that many toxic substances including heavy metals accumulate in offspring, and this may influence survival and reproduction of subsequent generations. 20 Some studies on the intergenerational toxicity reveal almost similar effects across generations. 30,31 Because single-generation studies are more common than the intergenerational, this has biased the literature, thus giving the impression that intragenerational studies alone are sufficient to derive adequate information from which safe concentrations of heavy metal may be determined. Our intergenerational study of life history traits of M macrocopa exposed to Cd revealed interesting responses; some variables such as average life span, gross and net reproductive rates were strongly reduced through generations, while others such as generation time and rate of population increase showed no clear trends. In the long term, potential negative transfer effects of toxicants across generations to offspring indicate an array of stress symptoms in planktonic species. 32 -34
In our study, M macrocopa also experienced stimulatory effects at low cadmium stress but only in few life history variables of the F0 generation. For example, M macrocopa in F0 exposed to Cd at 0.2 mg/L showed significantly higher gross and net reproductive rates, and the rate of population increase as compared to controls or other Cd levels. This enhanced reproductive output under low stress conditions is considered as hormesis. 35 In hormesis, the stress response systems in organisms are started at low concentrations of toxicants which results in a stimulation of metabolism and hence overcompensation of offspring production. 36 Another explanation of this behavior is in terms of trade-offs between various biological endpoints (different traits), in which there could be no net gain to the species in question. 37,38 In other words, enhanced reproductive rates due to low toxicant stress could have the negative consequences involving some other life history traits such as reduced body size which was not determined in this work, although such trends have been documented in literature. 39 An adaptive response that may shape the life history traits of offspring is to rely on the maternal influence, in addition to the environmental conditions currently experienced by an organism. 11,12 In this sense, we found that Cd had cumulative negative effects on the F1 progeny of M macrocopa in nearly all life traits, being more intense for those related to reproductive output.
Multigenerational approach is useful for understanding the role of adaption to test conditions. Indeed, a few studies involving multigeneration approach have shown reduced tolerance to sublethal metal toxicity after long-term exposure. For example, it has been documented a higher MeHg retention by the F1 generation of Daphnia magna than that by the parental generation (F0), which led to lower reproductive output in daphniids. 30 This is also evident in our study where up to F4 generation, there was a continuous reduction in offspring production due to Cd at concentrations as low as 0.2 mg/L. Like many other aquatic organisms, cladocerans also possess a range of detoxification mechanisms with metabolic costs which could induce increased energy consumption and with associated adverse effects on reproduction and growth. 40,41 Although we have not quantified algal consumption rates of M macrocopa, cumulative, chronic toxic effects are also possibly due to feeding depression. This may have caused lower toxicity threshold, as pointed out by Zhao and Wang, 42 and consequential parental nutritional deficiency in the offspring that can contribute to transgenerational toxicity. 30,32 In rotifers grown under stressful conditions, it has been shown that maternal investment enhanced life span of the next generation through enhanced production of antioxidant enzymes. 43 Moina macrocopa in this study may have behaved in a similar way but only in a few cases. For example, the average life span of M macrocopa subjected to Cd at 0.2 mg/L was significantly higher than controls for both F0 and F1 generations. Although we did not observe any recovery patterns in the reproductive output as a result of continuous exposure to Cd, we did detect enhanced gross and net reproductive rates, as well as the rate of population increase for at least first 2 generations. This may be in part explained by the observations of Gustafsson et al 33 and Jiang et al 44 who showed increased tolerance of organisms to toxicants over long-term exposure through several generations.
Our study highlighted that with Cd toxicity over 4 consecutive generations, M macrocopa continued to decline in its reproductive output. This suggests that demographic evaluations of cladocerans in cadmium-contaminated environments conducted for a single generation do not guarantee the long-term survival of zooplankton species.
Conclusion
Exposure of M macrocopa to cadmium stress levels over 4 consecutive generations revealed a demographic process of mostly reduced responses of this species. Fecundity in this cladoceran species showed a clear and significant reduction not only in magnitude but also in frequency. Progenitors’ exposure to Cd stress appeared to be the basic feature for reduced offspring production in subsequent generations. Potential tolerance and recovery trends were evident in only a few life history variables. Generation time appeared to be largely unaffected due to Cd stress across the tested generations.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors (JLGF, SSSS, and SN) thank CONACyT for financial support (SNI 40405, 18723, and 20520, respectively).