
Abstract
Purpose: To analyze the long-term effects of exposure to low-dose γ-irradiation on survival and life span in directly irradiated Daphnia magna and their offspring. Materials and methods: One-day-old Daphnia magna were exposed to 10, 100 and 1000 mGy of acute γ-rays. The life-span of irradiated and control animals, as well as their non-exposed progeny was analyzed. Results: Irradiation at dose of 100 and 1000 mGy resulted in a significant decrease in the life span of irradiated Daphnia. The data presented here also show that the survival of non-exposed first-generation offspring of irradiated parents is significantly compromised, thus implying the presence of transgenerational effects. Conclusion: The results of our study show that low-dose irradiation significantly reduces the life-span of directly exposed Daphnia and their first-generation offspring. The observed compromised viability is most likely attributed to non-targeted effects of ionizing radiation.
1. INTRODUCTION
The effects of low-dose exposure to ionizing radiation on life span of animals and humans still remain poorly understood. Given that quite often such analysis is extremely time-consuming and requires profiling a large number of exposed animals, the introduction of new and highly sensitive experimental models should provide further insights into this important issue.
The target theory can only provide some explanation of the effects of exposure to ionizing radiation on life span. This theory, independently developed in 1947 by Timofeeff-Ressovsky and Zimmer, and Lea (Timofeeff-Ressovsky and Zimmer 1947; Lea 1947), assumes that mutation induction mainly occurs in the directly exposed cells at the damaged sites. However, a number of recent studies have clearly demonstrated the existence of non-targeted effects of ionising radiation, including radiation induced genomic instability, detected among the non-exposed progeny of irradiated cells/organisms (Morgan 2003a, 2003b; Dubrova 2003), and bystander effect, whereby an elevated mutation rate is detected in the non-exposed cells, which receive signals from the irradiated neighbours (Morgan 2003a, 2003b; Lorimore et al. 2003). The contribution of the non-targeted mechanisms to the survival of irradiated organisms still remains poorly understood.
The results of a number of previous studies show that the effects of low-dose irradiation can properly be explored either in vitro (Morgan 2003a; O'Dowd et al. 2006) or, alternatively, in vivo by using some invertebrate experimental models, such as earthworms, crustaceans and insects (Hertel-Aas et al. 2007; Gilbin et al. 2008; Nakamori et al. 2009). It should be stressed that the results of these studies are often in line with those obtained in mice and rats, as well as the results of large epidemiological studies (Dauer et al. 2010). As far as the effects of low-dose exposure are concerned, it has been shown that some of them clearly challenge the existing paradigm in radiation biology, which is solely based on the target theory. It has been suggested that these non-targeted effects of low-dose irradiation could provide a plausible explanation for some experimental observations, including, for example, the unusual high efficiency of low-dose exposure, radiation-induced genomic instability and bystander effects (Geras'kin et al. 2007; Barber et al. 2009; Mothersill and Seymour 2010).
Over recent years the work in our research group has been focused on the analysis of non-targeted effects of ionizing radiation. We have studied the effects of irradiation on cell motility in unicellular paramecium and found that exposure to even very low doses of γ-rays can substantially decrease the mobility of directly exposed organisms as well as their non-exposed progeny (Sarapultseva 2008; Sarapultseva and Bychkovskaya 2010a). For example, according to our results the alterations in cell mobility manifest in the progeny of irradiated cells up to 15 generations following the initial insult.
We have previously published the results of a study aimed to analyze the effects of acute exposure to γ-rays on mortality of the directly exposed Daphnia magna (Sarapultseva and Malina 2009). Similarly to the paramecium data, the results of our study showed that the mortality in the first-generation non-exposed progeny of parents irradiated to doses of 0.1, 1 and 20 Gy significantly exceeds that in controls (Sarapultseva and Bychkovskaya 2010b). Most importantly, our results clearly showed that the effects observed among the offspring are independent from the dose of parental exposure, thus providing clear evidence for the non-targeted mechanisms underlying this phenomenon, including the bystander effect. Moreover we obtained that the life span of Daphnia, exposed to the dose of 0.1 Gy was significantly compromised (Bychkovskaya and Sarapultseva 2011). To further investigate the non-targeted effects of ionizing radiation on survival, we have designed a study aimed to analyze the life span of Daphnia, exposed to wide range of low doses of γ-rays from 10 to 1000 mGy and their non-irradiated progeny.
2. MATERIALS AND METHODS
The life span of Daphnia is ∼10–11 weeks and is characterized by relatively short periods of embryonic development (3–4 days) and gestation (5–8 days). Daphnia magna reproduces by parthenogenesis, which allows quick and convenient establishment of genetically identical strains.
The strain of D. magna Straus used in our experiments was originally collected in the pond of the Moscow Zoo and successfully reared for 5 years in our laboratory. Daphnia were kept at 20 – 2° C in aerated carbon filtered tap-water (pH 7.7) renewed every week and fed with algae suspension (Chlorella vulgaris) obtained from the Department of Hydrobiology Moscow State University (density of 600–1000 million cells per ml). One-day-old Daphnia were given 10, 100 and 1000 mGy of acute γ-rays delivered at 17–100 cGy min−-1 (60Co source, Lutch Irradiator, Latenegro, Latvia).
Forty irradiated or control Daphnia were transferred to individual 150 ml glass vials (Simax, Sklarny Kavalier, Czech Republic). To each vial, containing 5 Daphnia in 100 ml aerated carbon filtered tap-water, 0.1 ml algal suspension was added daily. Irradiated and control Daphnia were checked daily for survival over the period of 2 months. All dead and neonates were removed. To analyze the transgenerational effects of parental exposure, forty one-day-old neonates were randomly taken from irradiated and control groups and transferred to glass vials (5 Daphnia per vial). The survival of their first-generation parthenogenetic progeny was measured over the period of one month.
The mean and variance of life span for controls, directly exposed Daphnia and their first-generation offspring were estimated using the Kaplan Meier approach (Rothman and Greenland 1998). Mean life span was estimated as:

where S k (survival rate) and S M are the probability of survival for the period of t k (k = 1 day) and for entire period of observation, respectively. The survival curves were compared using the log-rank test (Hosmer et al. 2008).
3. RESULTS
3.1. The effects of irradiation on the life span of directly exposed Daphnia
The survival curves for irradiated and control Daphnia are shown on Figure 1. Using the Kaplan-Meier statistical technique, we estimated the mean survival rates for controls and irradiated Daphnia (Table 1). According to our estimates, acute exposure to 100 and 1000 mGy of γ-rays significantly shortened the life span of exposed Daphnia (20–30% reduction), whereas the survival rate of those exposed to 10 mGy did not significantly differ from controls. It should be noted that following exposure to 100 mGy the mean life span did not significantly differ for that in the group of Daphnia exposed to the higher dose of 1000 mGy (Z=1.07; P=0.1407). We therefore conclude that low-dose acute irradiation significantly affects their life span. We also note that for the interval of doses between 100 and 1000 mGy the effects of irradiation reach a plateau.
The effects of irradiation on the lifespan of Daphnia
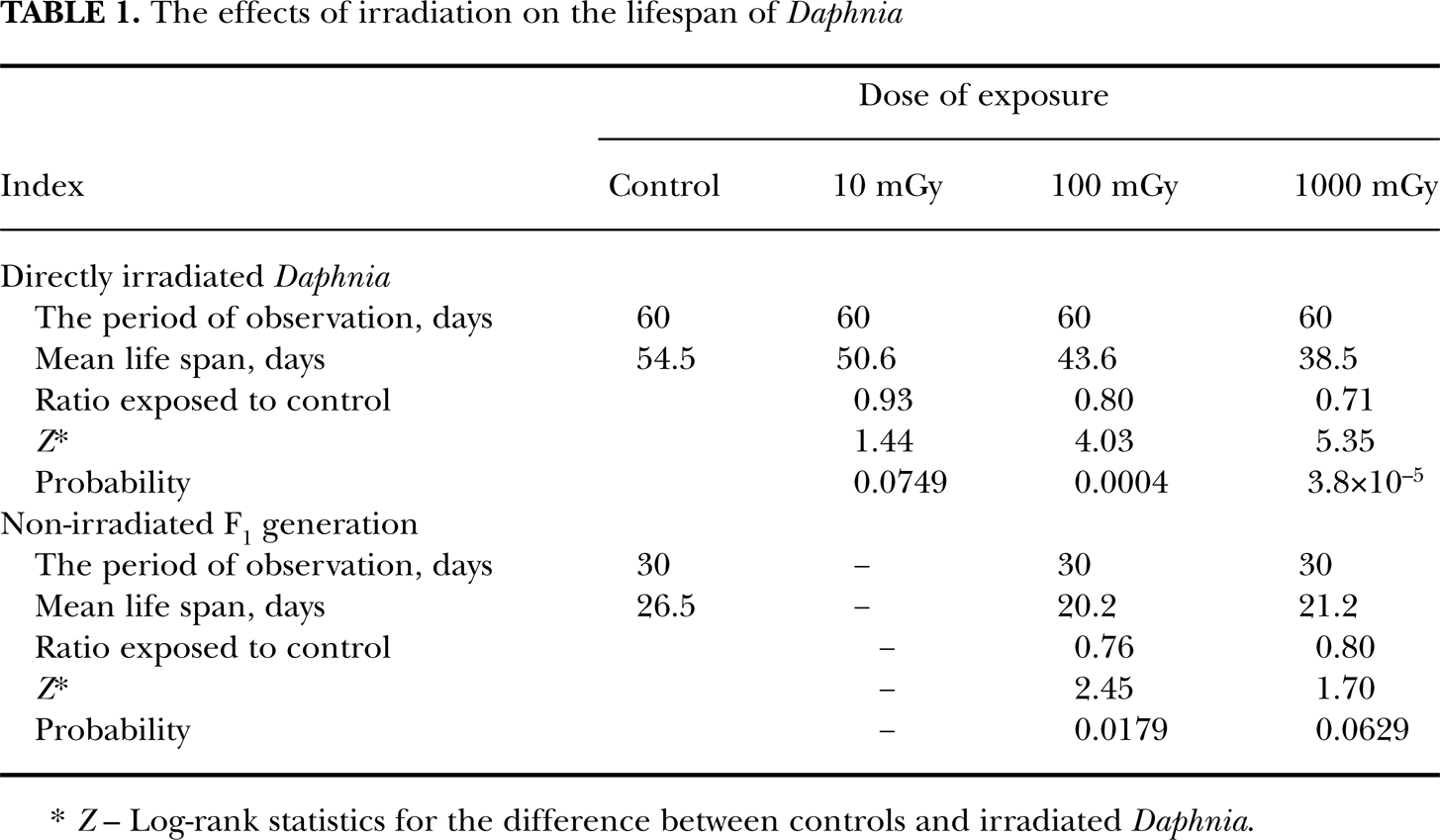
Z– Log-rank statistics for the difference between controls and irradiated Daphnia.

Survival curves for controls and directly irradiated Daphnia. Mean values for a single experiment (N = 40 for each control and exposed replica) are shown.
3.2. The transgenerational effects of parental irradiation
The survival curves for the first-generation offspring of irradiated and control parents are shown on Figure 2. Using the same statistical approach, we compared the mean survival for the three groups (Table 1). The life span of non-exposed first-generation progeny was significantly affected by the history of paternal irradiation. According to our data, in the offspring of irradiated parents mortality significantly exceeded that in controls. The magnitude of decrease in the mean life span (∼20%) was similar in the offspring of parents exposed to 100 and 1000 mGy of acute γ-rays (Z=0.34; P=0.3660).

Survival curves for the first-generation offspring of irradiated and control parents. Mean values for a single experiment (N = 40 for each control and exposed replica) are shown.
4. DISCUSSION
This study was designed to establish the long-term effects of low-dose acute γ-irradiation on the life span of directly exposed Daphnia and their first-generation offspring. The analysis of survival rates has revealed that: (i) the survival of Daphnia exposed to 100 and 1000 mGy of γ-rays was significantly compromised; (ii) the life span of non-exposed first-generation offspring of irradiated parents was similarly affected; (iii) the magnitude of effects of irradiation on survival of directly exposed Daphnia and their offspring did not increase within the range of doses from 100 to 1000 mGy.
The data presented here are in line with the results of a number of recent studies aimed to analyze the effects of low-dose irradiation on directly exposed animals and plants. It has been reported that exposure to 100 mGy of γ-rays can significantly compromise the in vitro survival of rainbow trout spleenocytes and also alter the activity of enzymes involved in oxidative phosphorylation (O'Dowd et al. 2009). Geras'kin and co-workers found that doses of exposure as low as 50 mGy can cause cytogenetic damage in barley cells (Geras'kin et al. 2007). Significant increases in the yield of chromosome aberrations were also found in human lymphocytes following exposure to the doses of 40–50 mGy from low-Linear Energy Transfer (low-LET) sources (Pohl-Ruling et al. 1983; Lloyd et al. 1992).
Taken together, these data indicate that exposure to dose as low as 100 mGy can result in a number of somatic alterations, including DNA damage and mutation induction.
The data on the plateau of radiation induced mortality in Daphnia exposed to 100 and 1000 mGy are in agreement with the results of our previous studies on aquatic protozoa ciliates Spirostomum, showing dose-independent effects of irradiation on a number of physiological traits (Sarapultseva 2008, Sarapultseva and Bychkovskaya 2010a). The results of previous work aimed to analyze the effects of irradiation on Daphnia survival also suggest the presence of a plateau for a range of doses from 100 mGy to 20 Gy (Sarapultseva and Malina 2009, Sarapultseva and Bychkovskaya 2010b). It should be noted that Geras'kin and co-workers also reported that in irradiated barley the yield of chromosome aberrations reached a plateau within the range of doses from 50 to 500 mGy (Geras'kin et al. 2007). The same phenomenon was found in a number of studies on the effects of low-LET irradiation on different species (Dauer et al. 2010).
The results of our study show significant decreases in the life span of irradiated Daphnia not only to the dose of 100 mGy (Bychkovskaya and Sarapultseva 2011), but also to the dose of 1000 mGy and the absent of effect to the dose of 10 mGy. The same effects of low dose of radiation were recently reported by Oghiso and co-workers, who analyzed the effects of 400 mGy of γ-radiation on the life span of exposed mice (Oghiso et al. 2008).
Moreover the data presented here also show that the survival of non-exposed first-generation offspring of irradiated parents is significantly compromised, thus implying the presence of transgenerational effects. It should be noted that, to date, the transgenerational effects of parental irradiation have mostly been analyzed in mammals (reviewed by Barber and Dubrova 2006) and therefore our results provide important evidence for the presence of these effects in other species. It should be noted that the mechanisms underlying transgenerational effects remain poorly understood. It would appear that the compromised survival of the first-generation offspring of irradiated Daphnia may be explained by the detrimental effects of deleterious mutations induced in the germline of irradiated parents. If this is the case, then their frequency in the offspring should correlate with the parental dose of exposure, as exposure to ionizing radiation results in a linear increase in the yield of mutations induced in the parental generation and passed to their parthenogenic progeny. Although the pattern of mutation induction in Daphnia remains poorly understood, it would appear highly probable that exposure to the two quite distinct doses of 100 and 1000 mGy should result in accumulation of a different number of radiation-induced mutation affecting the viability. However, according to the results of our study the mean life span of the offspring of parents exposed to 100 and 1000 mGy of γ-rays does not substantially differ. Therefore, similarly to the above-mentioned mammalian data (Barber and Dubrova 2006), the observed transgenerational effects of parental irradiation in Daphnia may be attributed to epigenetic changes occurring in the germline of exposed parents and their manifestation in the offspring. It should be noted that our results are in line with a number of recent data suggesting that the epigenetic inheritance may play an important in evolution (Jablonka and Raz 2009; Dauer et al. 2010; Vandegehuchte et al. 2010).
The results of our study also provide further support for the hypothesis of non-targeted effects of ionizing radiation. Indeed, the unusually high effectiveness of low-dose irradiation, the lack of dose response and the presence of transgenerational effects are the well-established hallmarks of non-targeted effects (UNSCEAR 2008). As already mentioned, similar data were also reported in our previous studies on the survival of protozoa and daphnia (Sarapultseva and Bychkovskaya 2010a, 2010b).
In conclusion, our current and previous (Sarapultseva and Malina 2009; Sarapultseva and Bychkovskaya 2010b) findings show D. magna provides a useful and very sensitive experimental model for the analysis of long-term effects of exposure to ionizing radiation.
Footnotes
ACKNOWLEDGEMENT
The authors would like to thank Professor Irena B. Bychkovskaya for discussion of experimental data. We also thank J. Kulish for expert care of Daphnia, J. Malina and T. Kolesnikova for participation in experiments and A. Brovin for dose calculation and dose control.
DECLARATION OF INTEREST
The work was supported by the Federal Target Program ‘Scientific and scientific-pedagogical personnel of innovative Russia’ (2009 – 2013), project N P968.