
Abstract
Background. Poststroke, the ipsilesional upper limb shows slight but substantial and long-term motor deficits. Objective. To define brain activation patterns during a gross motor flexion/extension task of the ipsilesional elbow early poststroke before and after rehabilitation, in relation to the corresponding kinematic characteristics at each time point. Method. Simultaneous analysis of kinematic features (amplitude, frequency, smoothness, and trajectory of movement) and of corresponding functional magnetic resonance imaging activations (block-design). A total of 21 persons with subacute initial severe stroke (Fugl-Meyer score <30/66) participated twice: within the first 2 months poststroke (V0) and after 6 weeks of rehabilitation (V1). Results at both time points were compared with activation patterns and kinematics of 13 healthy controls. Results. Compared with controls (a) movements of the ipsilesional upper-limb poststroke were smaller (V0 + V1) and less smooth (V0 + V1) and (b) participants poststroke showed additional recruitment of the contralesional middle temporal gyrus (V0) and rolandic opercularis involved in movement visualization (V0 + V1), whereas they lacked activation of the supramarginal gyrus (V0 + V1). Over time, participants poststroke showed an extended activation of the contralesional sensorimotor cortex at V0. Conclusion. Movements of the ipsilesional upper limb within an initially severe stroke group were not only atypical in motor outcome, but seemed to be controlled differently. Together the observed changes pointed toward an overall disturbance of the bihemispheric motor network poststroke, marked by (a) a possible despecialization of the nondamaged hemisphere and (b) the employment of alternative control strategies to ensure optimal task execution.
Keywords
Introduction
Daily, people poststroke use their “unaffected” arm up to 6 times more than their affected or paretic arm. 1 However, a growing body of research has established that the “unaffected side” after unilateral stroke is actually also affected. 2 In the first month poststroke, proximal arm muscles show substantial weakness, with clumsy and slow performance. 3 Then, 3 to 4 months poststroke, although accuracy approaches normal levels,4-6 the movement remains less smooth 5 . Even chronically post-stroke, the amplitude of the peak acceleration during pointing is decreased, albeit its timing being comparable to that of controls. 7
Several hypotheses have been put forward to explain the deficit of the ipsilesional upper limb poststroke, like the disruption of ipsilesional descending pathways of the corticospinal tract, 8 cognitive disorders affecting attention, perception, and control of action, 9 and interhemispheric disbalance by transcallosal inhibition and excitation processes.10,11 Although the relative contribution of these processes is still under debate, agreement exists about its holistic impact on the bihemispheric network functioning. It underpins recent neuromodulation rehabilitation techniques like transcranial direct current stimulation, 12 and it explains contralesional recruitments during paretic upper-limb movements. 13 Surprisingly, it remains unclear how the general disturbance affects brain activations during ipsilesional upper-limb movements.
Here, we aim to investigate brain activation patterns during gross motor movement of the ipsilesional upper limb early poststroke in persons with an initial severe motor deficit in relation to kinematic movement characteristics before and after rehabilitation. In order to allow for later comparisons with paretic motor control, an elbow flexion/extension task was evaluated rather than traditional fine motor finger-tapping or wrist movement paradigms, as recovery post-stroke generally evolves from proximal to distal. 14 The usability of the task has been previously validated with healthy controls. 15
Based on the holistic point of view, we expect to observe an increased contralesional hemispheric activation following ipsilesional elbow movement. This may result from the disturbed inter-hemispheric balance and possible disrupted ipsilesional descending pathways of the corticospinal tract that together increase the load on the contralesional hemisphere. This effect is expected to be strongest immediately poststroke, when the control is disturbed not only by the lesion itself but also by secondary effects like edema and disturbed metabolic processes. 16 In addition, if cognitive disorders are of great importance, we expect to observe changes in motor planning regions in the frontal cortex. 17
Methods
Participants
Twenty-one poststroke patients (S: 15 males, age 59 ± 13.2 years) with a first-ever unilateral ischemic stroke and an initial severe motor deficit and 13 healthy controls (C: 8 males, age 44 ± 14.5 years) with no history of neurological, psychiatric, or orthopedic disease participated in this study. All poststroke participants suffered occlusion of the middle cerebral artery (MCA). The study protocol received ethic approval. Procedures were compliant with the declaration of Helsinki. The study was registered as clinical trial (NCT01554449) and was part of a larger protocol, focusing on rehabilitation of the upper limb early poststroke (MARGAUT, 2010-A00596-33). Participants were recruited at the University Hospital of Montpellier and Nimes. All gave written consent before inclusion. A flowchart is presented in Figure 1. Exclusion criteria were (a) age <18 years, (b) occurrence of other neurological or psychiatric diseases, (c) Fugl-Meyer score 18 >30/66, (d) severe aphasia (Boston diagnostic aphasia examination scale 19 <3/5, (e) strong neglect (bell test score 20 >8 missed bells), (f) contraindications to magnetic resonance imaging (MRI), (g) bihemispheric stroke, and (h) hemorrhagic stroke. Handedness was evaluated using the Edinburgh Handedness Inventory. 21 Exclusion criteria were applied during the first week poststroke. The Fugl-Meyer score was evaluated a second time at study onset.
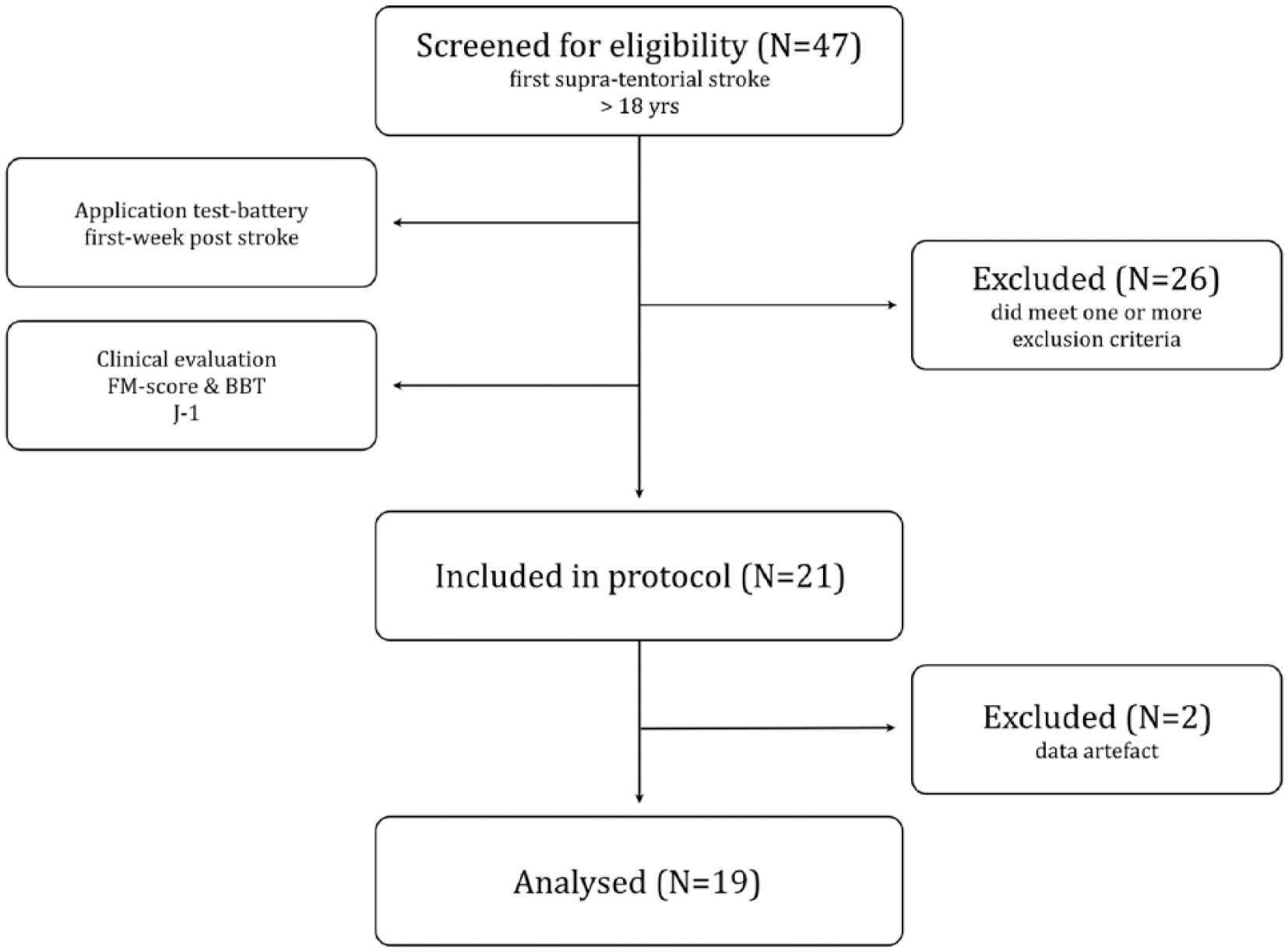
Flowchart of poststroke participants’ inclusion.
Clinical Evaluation
Clinical scores provide an overall image of paresis severity participant-by-participant. The Fugl-Meyer Assessment (FM) score quantifies motor function of the paretic upper limb, based on a sensori-motor assessment with a maximum score of 66 points. 18 Upper-limb dexterity was assessed with the Box and Block Test (BBT). The BBT required moving as many 4 × 4 cm blocks, block by block, from one box to another within 1 minute. The normative score for the age group 60 to 64 years is to transfer 70 to 75 blocks. 22 Correct execution of the BBT requires elbow flexion/extension.
The FM score evolution demonstrated that none of the participants showed full motor recovery of the paretic upper limb. Patients’ paretic limb dexterity was highly compromised: at study onset, only 2 participants were able to transfer some blocks. After 6 weeks of rehabilitation, BBT scores of the paretic upper limb (BBT-p) improved for 13 patients (Table 1). The majority of the participants also showed improvement of the ipsilesional upper-limb (BBT-il). FM scores of the ipsilesional upper limb were not administered.
Poststroke Participant’s Characteristics. a
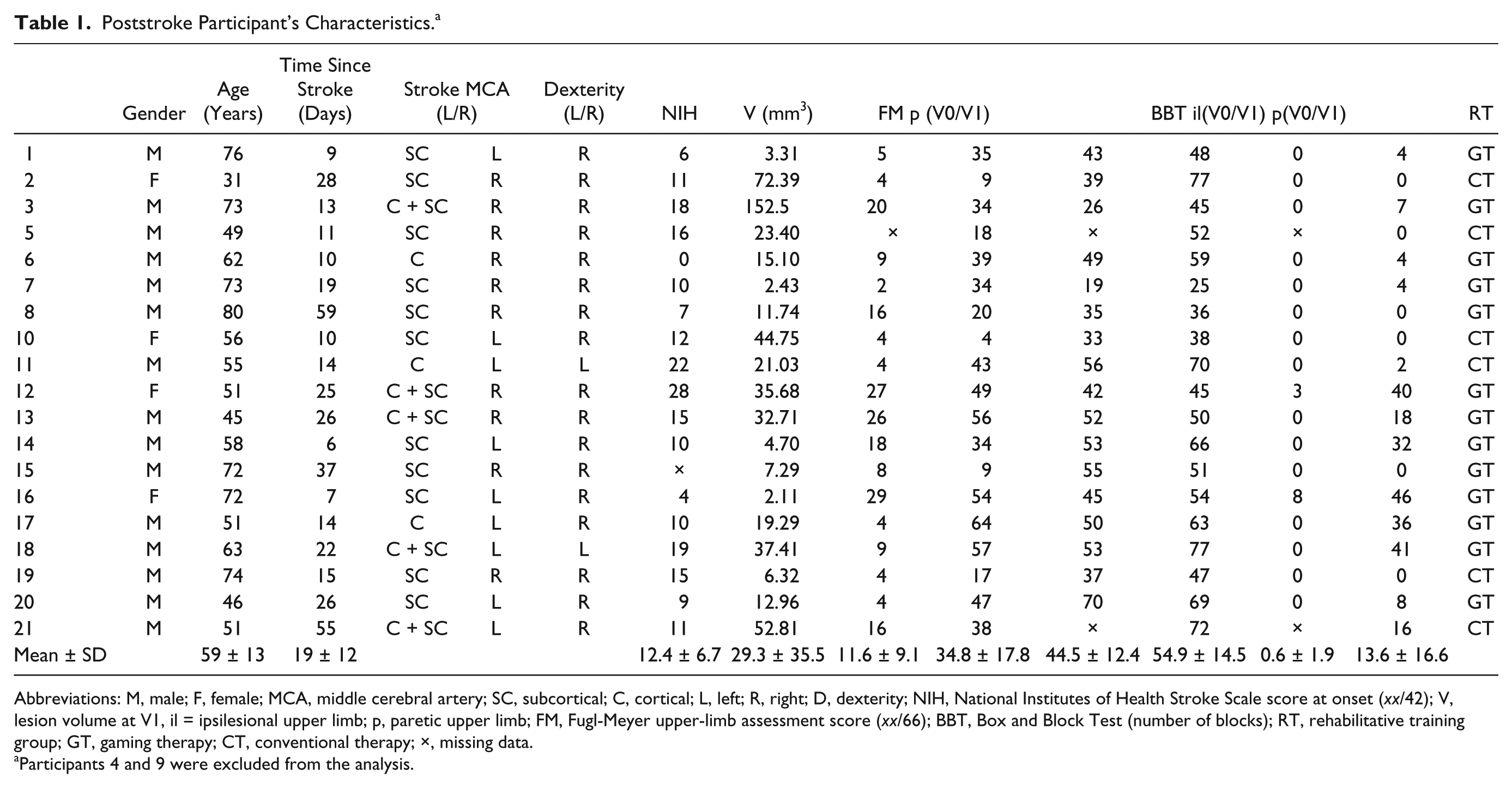
Abbreviations: M, male; F, female; MCA, middle cerebral artery; SC, subcortical; C, cortical; L, left; R, right; D, dexterity; NIH, National Institutes of Health Stroke Scale score at onset (xx/42); V, lesion volume at V1, il = ipsilesional upper limb; p, paretic upper limb; FM, Fugl-Meyer upper-limb assessment score (xx/66); BBT, Box and Block Test (number of blocks); RT, rehabilitative training group; GT, gaming therapy; CT, conventional therapy; ×, missing data.
Participants 4 and 9 were excluded from the analysis.
Protocol
The study consisted of 2 separate functional MRI sessions: V0 (study onset <2 months poststroke) and V1 (V0 + 6 weeks). In between, participants received standard rehabilitation with additionally 5 times a week 45 minutes of conventional or therapeutic game-based rehabilitation. Rehabilitative training was executed with and focused on the paretic upper limb. Experimental setup can be found in Figure 2.
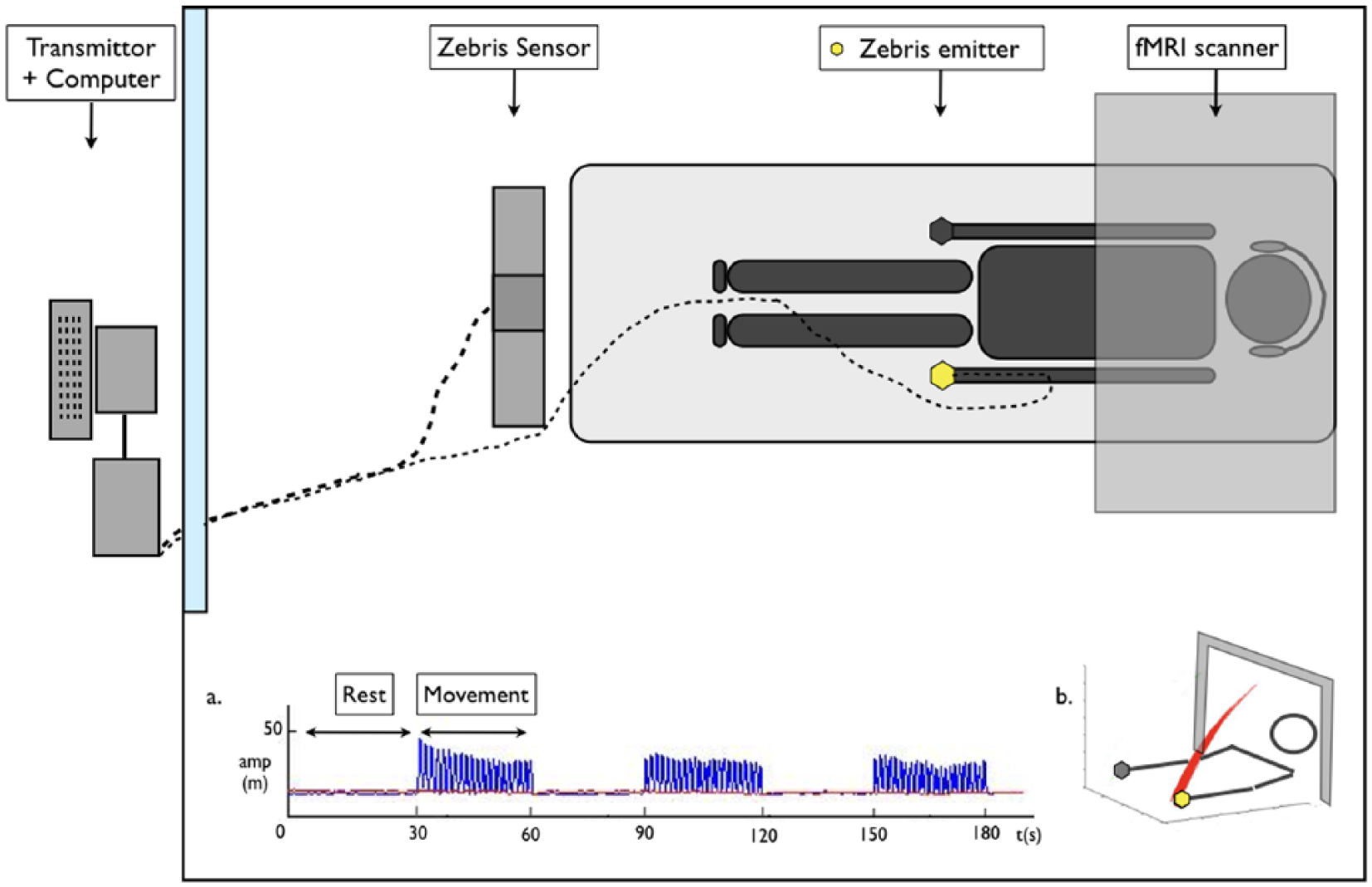
Experimental setup with (a) the movement acquisition in time following a block-design and (b) exemplary healthy movement in 3-dimensional space. Amp (m) = amplitude in meters, t (s) = time in seconds. Adapted from van Dokkum et al (2017). 15
During each functional magnetic resonance imaging (fMRI) session, participants were asked to perform a continuous elbow flexion/extension movement with the ipsilesional upper limb in a blocked design: 30 seconds of rest followed by 30 seconds of activity, repeated 3 times within 1 acquisition (Figure 2a). During rest, the arms were extended alongside the body. The fingers were flexed without forcing. During movement, only the elbow flexed and extended rhythmically at a self-selected comfortable pace in the vertical plane (for a typical movement representation see Figure 2b). The maximum flexion amplitude was constrained by the MRI tube. Participants were free to select their comfortable movement amplitude within this material induced boundary. Moving at the self-selected preferred frequency with comfortable amplitude is thought to equal task-induced cost levels between participants.23,24
Before each fMRI session, protocol proceedings were explained. The required movement was practiced at least twice in room setting and once before acquisition onset in the MRI. Participants were instructed to relax, to not move anything but their lower arm and to try to execute the movement in the vertical plane. They were told to move at a comfortable pace and amplitude without entering their hand in the MRI tube, and were instructed to avoid head movements. The protocol was first validated with the healthy control group 15 and subsequently evaluated in a pilot-study, including 6 people chronically poststroke, with various levels of severity.
To limit the task dependent increased risk of head movement, the participant’s head was fixed with supportive foam blocks. Participants exceeding 2 mm of pitch, roll, or jaw displacement were excluded from further analysis. To limit interference with eye movements and stimulate proprioceptive control, eyes were closed throughout the protocol. Controls executed the movement task with their dominant side. All were right-handed.
Material
fMRI acquisitions were performed with a 1.5 whole-body MRI system (Magneton Avanto, Siemens, Erlangen, Germany), equipped with a standard 12-channel receive-only head coil. The acquisition consisted of 60 BOLD (blood oxygen level–dependent changes) echo-planar images (EPI), using a block-design in which 10 “resting” volumes (both arms stretched alongside the body) were alternated with 10 “active” (continuous flexion/extension of the elbow) volumes (TR/TE = 3560/50 ms, field of view/slice gap = 230/0.75 mm, 36 axial slices extending from the vertex to lower parts of the cerebellum, voxel size 3.28 × 3.28 × 3.75mm3, flip angle = 90°). A rapid auditory stimulus (500 ms) signaled block onset. A 3DT1 MPRAGE (TR/TE/TI 2100/4.1/1100 ms, 15° flip angle, aligned with the corpus callosum, voxel size 0.98 × 0.98 × 1.0mm3, 176 transversal slices) was obtained.
Movement characteristics were registered with a 1.5-T MRI compatible 3D ultrasound motion capture system (sample frequency: 20 Hz) (Zebris, CMS-20, Medical GmbH, Isny, Germany). Data were low-pass filtered (5 Hz) with a zero-lag Butterworth filter. The ultrasound emitter was placed on the head of the third proximal phalange. The Zebris receiver was placed in front of the fMRI tube (Figure 2). Pilot evaluations confirmed the absence of interference between the Zebris and the 1.5-T MRI.
fMRI Data Analysis
Data were spatially preprocessed using SPM8 (Statistical Parametric Mapping toolbox, http://www.fil.ion.ucl.ac.uk/spm/) with MATLAB R2012a, The MathWorks. The EPI were reoriented to the anterior commissure, slice-time corrected, realigned to the first volume, co-registered, normalized to 3DT1 template (provided by Montreal Neurological Institute [MNI]), and smoothed with a 8-mm FWHM (full-width at half maximum) Gaussian filter. Potential normalization distortions were evaluated using (a) an individual based gray-matter mask and (b) a DARTEL (diffeomorphic anatomical registration through exponentiated lie algebra) individualized template. After consulting the neuroradiologist, the former method was selected showing lowest incoherence levels. These standard analysis procedures have previously been published by our group. 15
The brain activations related to the motor task were evaluated using a general linear model analysis. Realignment parameters were included. A voxel-by-voxel based evaluation was used to calculate statistical t-maps. The T-threshold was set at P < .0005. Clusters >10 voxels that exceeded the threshold were considered activated. A whole-brain multiple comparison (family-wise error [FWE]), P < .05, was applied at cluster level to define relevant clusters.
First, we performed separate analyses for left- and right-sided strokes. The effects of lesion side, dominance, and paretic upper limb rehabilitation (traditional vs game) were evaluated using independent T tests (P < .05, uncorrected). Group analysis was subsequently performed, whereby the images of participants with a left-sided stroke were 180° flipped around the mid-axis so that all lesions appeared virtually at the right side (Figure 3). All fMRI analyses were corrected for subtle possible age effects. In addition, the main effect of age was evaluated using a full factorial design, including all participants and both acquisitions times poststroke.
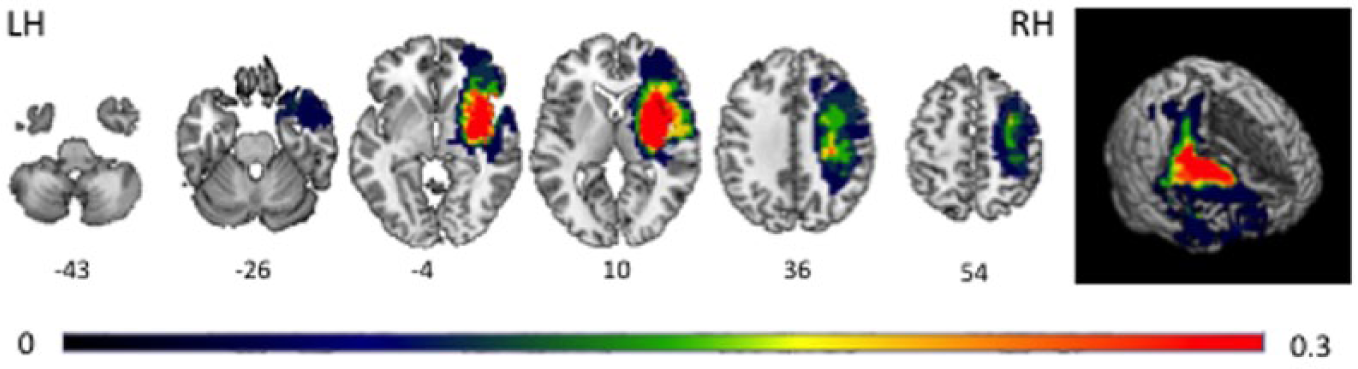
Probability map of stroke localization of all poststroke participants.
Clusters were localized using local maxima with the automated anatomic labeling toolbox (AAL). 25 Within large clusters multiple local maxima were identified.
Kinematic Data Analysis
Variable selection was based on previous research demonstrating that movement organization in healthy and poststroke populations can be captured by the movement velocity, the smoothness and directness of the trajectory.26,27 Simple rhythmical movement is typically presented by a smooth and bell-shaped velocity profile that is slightly variable over cycles (Figure 2a and b). 28 This natural variability is caused by biological noise, inherent to our system and asking for continuous correction processes. 29 People poststroke exhibit increased noise levels, challenging motor control and slowing down correctional movements. 30 Although the intended movement was uniaxial in the vertical plane, especially people poststroke diverted from this trajectory with internal medial rotation of the shoulder. Based on the protocol validated with healthy people, 15 we used a principal component analysis of the x, y, z time series to extract the principal oscillation. It was parsed in subcycles of 1 flexion and extension movement using the maxima between 2 velocity peaks, and used to analyze the amplitude (AMP) and the frequency (FREQ) of movement. The amplitude of each subcycle was calculated by taking the absolute distance between the lowest and the highest point. AMP was defined as the mean amplitude of all subcycles. Next, we extracted participants’ preferred frequency (FREQ) by dividing the total number of subcycles by the time of movement.
Then, based on the complete time-series, we calculated the number of velocity peaks (NVP) and the structure of the trajectory in space (nTL) to gain insight in the motor control strategy used. Movements post-stroke tend to be highly segmented, while becoming smoother over recovery.5,27,31 To quantify smoothness, the simplest way and least sensitive to outliers, is to count the number of velocity peaks: the more peaks, the less smooth the movement. Recall that participants moved at their preferred frequency (variable number of cycles) with comfortable amplitude (variable absolute distance). Thus, to compare NVP values between a patient making large slow movements and a patient making fast small movements, the NVP was corrected for the signal’s corresponding FREQ and AMP. Consequently, the NVP value does not reflect a real number of peaks, but the number of peaks relative to the movement characteristics.
The trajectories’ structure was defined as the normalized movement trajectory length (nTL). Normalized by its standard deviation, the nTL is a dimensionless variable that reflects the amplitude of the deviation from the optimal trajectory rather than the actual trajectory length. 32 The higher the nTL value, the less straight was the hand path trajectory.
The kinematic variables were mathematically independent. Differences in kinematics were analyzed using repeated measures ANOVA, with GROUP (C/S) as between factor and TIME (V0/V1) as within factor. A nested model was used, as controls were tested once and poststroke participants twice. Effect size was estimated with the partial squared eta (
Results
Participants
Three participants were excluded: 2 participants poststroke because of exceeding head movements and 1 control participant because of fMRI artifacts. There were no effects of (a) side of lesion, (b) therapeutic intervention of the paretic upper limb, (c) hemispheric dominance, and (d) age on brain activations and on kinematics.
Functional Network Activation
The principal sensorimotor network controlling the continuous flexion/extension movement of the ipsilesional upper limb consisted of the contralateral primary motor cortex (M1), the primary sensory cortex (S1), the supplementary motor area (SMA), the ipsilateral cerebellum and the vermis. This pattern was comparable with that of the controls (see Table 2, Figure 4).
Overview of BOLD Activations of Movements of the Less Affected Upper Limb for Poststroke patients, Compared With the Dominant-Sided Movements for the Control Group. a
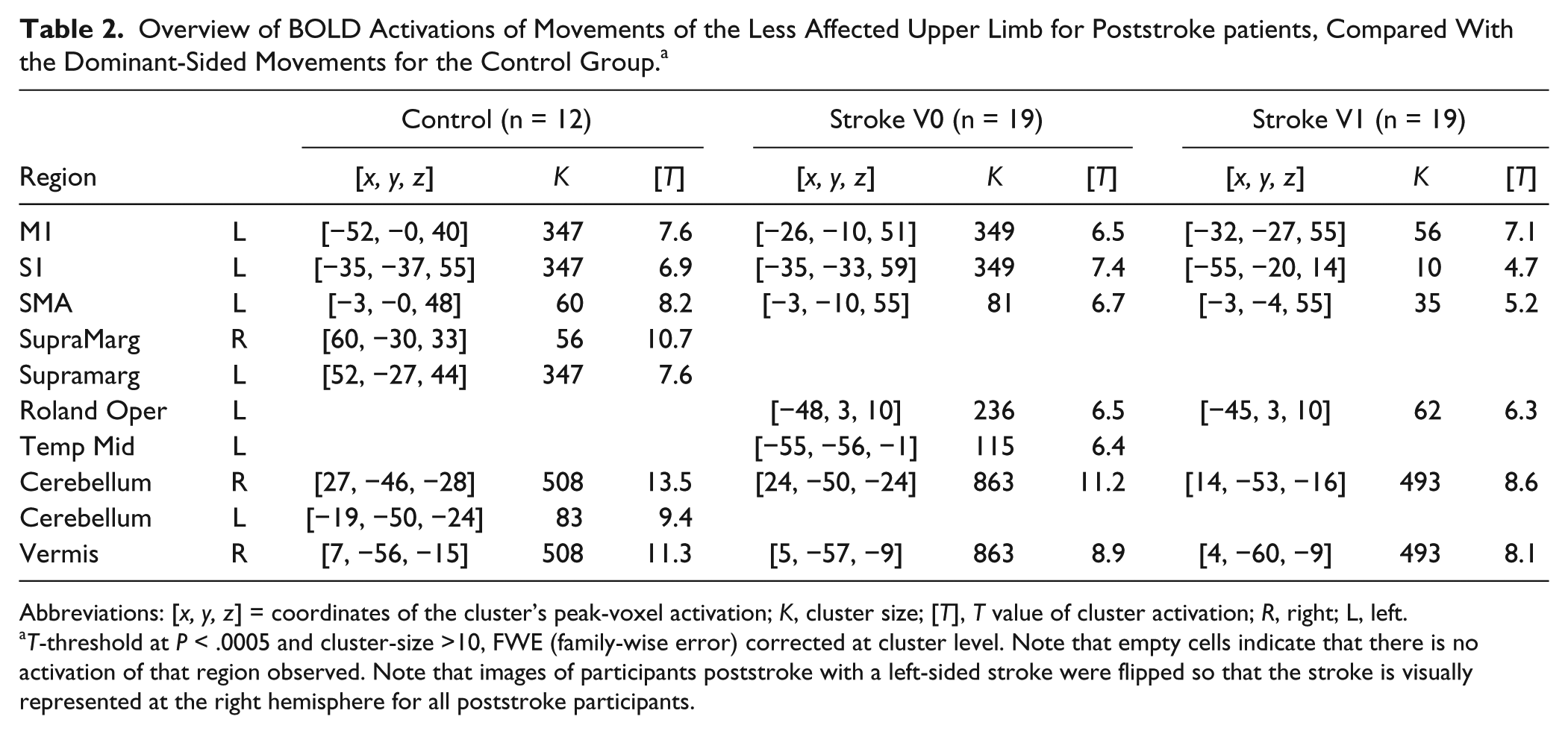
Abbreviations: [x, y, z] = coordinates of the cluster’s peak-voxel activation; K, cluster size; [T], T value of cluster activation; R, right; L, left.
T-threshold at P < .0005 and cluster-size >10, FWE (family-wise error) corrected at cluster level. Note that empty cells indicate that there is no activation of that region observed. Note that images of participants poststroke with a left-sided stroke were flipped so that the stroke is visually represented at the right hemisphere for all poststroke participants.
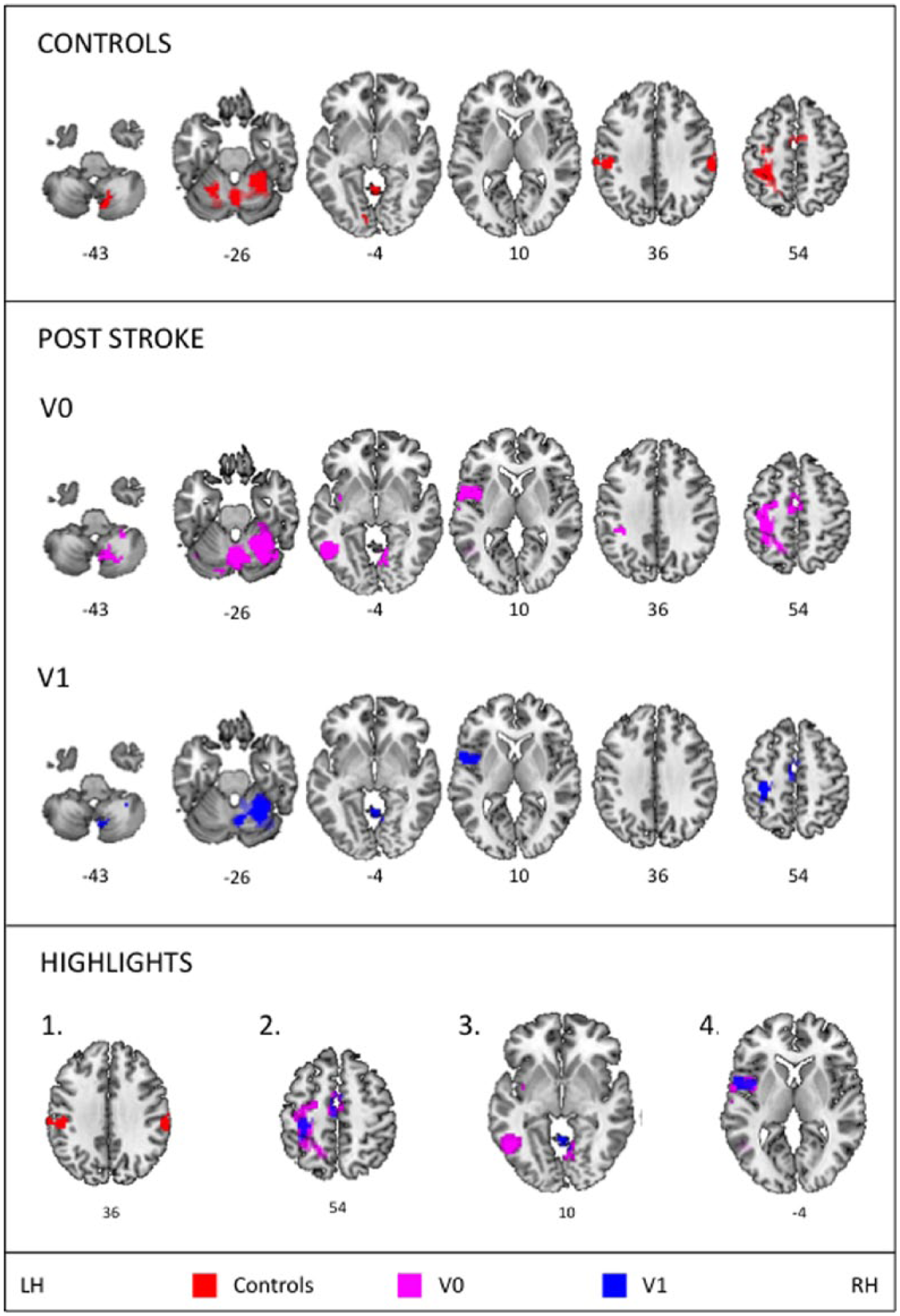
Group activations for the Controls and the Post-stroke group (V0, V1), including slice numbers, P < .0005, cluster-size >10, FWE (family-wise error) corrected (P < .05) at cluster level. Highlighted main results present: (1) a bilateral supramarginal activation in controls, (2) decreased activation from V0 to V1 poststroke, (3) activation of the middle temporal area at V0, and (4) activation of the rolandic operculum at V0 and V1. LH, left hemisphere; RH, right hemisphere.
In contrast: (1) controls showed a bilateral activation of the supramarginal gyrus, absent in poststroke participants; (2) the extent of the primary sensorimotor activation decreased from V0 to V1 poststroke; (3) the contralateral middle temporal area was activated at V0 poststroke; and (4) the contralateral rolandic opercularis was activated at V0 and V1 poststroke (see Figure 4 highlights).
Movement Kinematics
No effects of age, side of lesion, hand dominance, and lesion volume on kinematics were found, thus they were removed as covariates from the analysis. Significant GROUP effects were found for: AMP (F(1, 25) = 6.46, P = .018,
Clinical Scales
All clinical scores improved (F-M: +23.2 points, t = −2.278, P = .038; BBT-p: +13 blocks, t = −3.197, P = .006; BBT-il: +10.4 blocks, t = −3.783, P = .002). A significant correlation existed between F-M and BBT-p performances at V0 (r = 0.612, P = .01, standard error SE = 0.120, confidence interval CI [0.424-0.869]) and V1 (r = 0.692, P = .003, SE = 0.096, CI [0.477-0.858]). These clinical scores were unrelated to the BBT-il score. At V0, only the AMP was correlated to the BBT-il (r = 0.608, P = .016, SE = 0.152, CI [0.273-0.876]). This correlation disappeared when controlling for hand dominance and/or side of lesion, indicating that the relation between the initial amplitude and BBT-il performance was influenced by whether the ipsilesional upper limb was the dominant or nondominant side. None of the other kinematic variables were correlated to the clinical scores at V0 or V1.
Discussion
Movement deficits of the ipsilesional upper limb poststroke have been reported frequently2-7 and are confirmed by this study for gross motor movements in people with an initial severe motor deficit of the paretic upper limb. Although their ipsilesional upper limb moves with speed and trajectory comparable to controls, it does so with smaller amplitude and decreased smoothness. This kinematic profile was congruent with the one observed by Metrot and colleagues, 5 and it was independent of lesion volume and side, hand dominance, and clinical scores of both upper limbs.
Buma et al 33 demonstrated an association between the additional recruitment of secondary motor areas early poststroke and decreased smoothness during a finger flexion/extension task of the paretic upper limb. They suggested that the additional recruitment of sensorimotor areas poststroke reflects adaptive motor learning strategies rather than restitution of motor function. Focusing on brain activations associated with ipsilesional upper limb elbow movement, we found that (a) participants poststroke show no activation of the bilateral supramarginal region, (b) at V0 there was an extended activation of the contralateral sensorimotor network, (c) at V0 there was an additional recruitment of the contralateral middle temporal area, and (4) at V0 as well as V1 there was an additional recruitment of the contralateral rolandic opercularis.
These results underpin not only that the kinematic outcome of ipsilesional upper limb movement is atypical but also that the movement is controlled differently. As expected, extended activations were mainly observed at V0, while it returned to levels comparable to controls following presumably spontaneous and training induced recovery processes. The lack of frontal activations, indicate that participants understood the relative simple task and that cognitive deficits did not strongly influence its performance. The extended and novel activations do support the notion of an overall disturbance of the bihemispheric sensorimotor network poststroke. The fact that kinematics is unrelated to clinical scores and lesion characteristics might indicate that the altered behavior results from adaptive motor control strategies. In the following, we will first address in greater detail the possible underlying reasons for the extended sensorimotor activations. Second, we will analyze the additional recruitment of the temporal and rolandic areas, as well as the lacking supramarginal activation. And, third, the clinical impact of findings will be discussed.
Extended Sensorimotor Activations: Despecialization of the Contralesional Hemisphere
Both hemispheres play distinctive roles in motor control. The dynamic dominance model distinguishes between specialization in multijoint dynamics control (left hemisphere) and steady-state position control (right hemisphere). 34 In an extensive review, Serrien et al 35 emphasize that it is “through an active interplay of neural processing, mainly via callosal projections, that the transfer of information for sensorimotor integration, intention, decision making and response preparation is supported.” That is, a complex dynamic interplay between hemispheres, task, and performer related determinants, so that movement organization is reflected by a distributed engagement of multiple neural regions. 35 The simple product of functional specialization and functional integration is that both hemispheres are required to perform an optimally controlled movement, in such a way that when one hemisphere is damaged the movement and/or the network enabling movement is compromised.
In healthy people, extended sensorimotor activations are associated with motor learning over months of practice. 36 Even experts in complex motor tasks, like musicians, show such extended activations when confronted with a novel complex task. 37 Poststroke we observed extended activations at V0, which disappeared after six weeks of rehabilitation. Early poststroke, the interhemispheric network is challenged not only by the lesion itself but also by secondary effects like oedema and disturbed metabolic processes. 16 At this time, the contralesional hemisphere is recruited to contribute to the control of the paretic upper limb. 38 A temporary despecialization might be required to enable the nondamaged hemisphere to control both limbs at the same time. Well-learned automatic processes will then be controlled in a more conscious manner, resulting in activation patterns resembling control strategies during motor learning,13,37 that is, with an increased emphasis on feedback, resulting in less smooth movements.
We thus hypothesized that the extended activation pattern during ipsilesional upper-limb movement suggests a form of hemispheric despecialization that is required to enable the highly solicited nondamaged hemisphere to control both the paretic and ipsilesional upper limb. Because of the despecialization, the relative simple well-known flexion/extension movement needed to be “re-learned” at V0. Further research into motor (re)learning strategies post-stroke is required to verify this hypothesis.
Additional Recruitment: The Role of Visualization
The control group showed a bilateral activation of the supramarginal gyrus. Especially the right supramarginal gyrus is known to be involved in proprioception in both healthy controls and poststroke, whereby decreased activations are linked to decreased use of proprioception. 39 The absence of supramarginal activations in our participants poststroke might indicate that they did not rely on proprioceptive information. As a consequence, the recruitment of the additional temporal and rolandic opercularis regions poststroke might reflect an adaptive control strategy to enable task execution in the absence of visual and proprioceptive information.
The temporal cortex is, amongst others, recognized as one of the retinotopic network’s higher order regions. 40 Within this network, the middle temporal area is specifically related to spatial orientation processes serving movement. 41 It contributes to establishing the allocentric environment, that is, the location of objects relative to one and another. Damage to this area has produced difficulties in recalling visual stimuli with limited conscious visual perception. 42
The rolandic opercularis is associated with internal representations of movement 43 and is involved in both visual and kinesthetic motor imagery. 44 Motor imagery exhibits a supportive function during movement preparation and training. 45 A recent pilot study demonstrated how, poststroke, simultaneously imagining a functional movement (pushing a door handle) that incorporates the executed abstract movement (supination of the forearm) facilitated motor network activity during planning and execution. 46 Interestingly, this also induced activations of the rolandic opercularis.
Putting these pieces together, the subsequent hypothesis was deduced: when lacking reliable visual and proprioceptive information during ipsilesional movement post-stroke, visualizing the environment (middle temporal regions) and imagining the movement in this environment (rolandic opercularis) might represent an active adaptation strategy to ensure task execution. Over recovery, the visualization of the environment might have become less necessary, presumably related to improved proprioception. 47
Clinical Implications
The kinematic profile suggested a stronger feedback-based control of the ipsilesional upper limb, with brain activation patterns that resemble patterns of motor learning. We also demonstrated an additional role for visualization during task execution with closed eyes. The changes in motor control and kinematic outcome seemed independent of stroke characteristics, but rather related to despecialization of the highly solicited nondamaged hemisphere. How can these findings be translated into clinical practice? As the contralesional hemisphere seemed to despecialize to enable control of both upper-limbs at the initial phase of recovery, performing bimanual tasks during this phase could be an interesting way to train both limbs and optimize the efficiency of unihemispheric control. However, this should not come at the cost of unilateral training of the paretic limb, which requires a minimal amount of exercise to favor maximal recovery 48 and is also favorable to restore interhemispheric disbalance. 10 Bimanual training might therefore be especially interesting for those poststroke patients that seem unlikely to regain full functional activity of the lesioned hemisphere and are stronger dependent on direct corticospinal projections for paretic upper-limb movements. Because of limited sample size and with heterogeneous recovery profiles, we were unable to verify this hypothesis. Extended patient profiling with larger patient populations is required to allow for valid patient group categorizations in order to develop therapeutic intervention programs that are adapted to the individual patient.
The efficacy of motor imagery in poststroke rehabilitation has been frequently addressed because of the intervention’s simplicity and being neither physically exhausting nor harmful. Nor does it require existing motor control. 49 In addition to physical training it has proven its benefit in various domains including medicine, psychology and music. 50 Under the umbrella of mental practice it is recognized as an important training tool in sport since 1983. 51 Also, computer brain interfaces for motor learning support the potential of motor imagery based learning. 52 The impact of motor imagery or task visualization during rehabilitation has gained increasing attention over the past decade. Based on our results, it could be argued that visualization of a motor task during its execution might be an adaptive strategy that is intuitively applied to support movement execution. Together with the promising results of the aforementioned pilot study, 46 this argues to integrate visualization and mental imagery in a more structural manner during the execution of rehabilitative exercises.
Limitations
The results are based on a cohort with lesions following a blockage of the deep MCA. The MCA is implicated in more than two thirds of all infarcts with a 33% involvement of the deep territories. 53 Thus, although the results are based on a common type of stroke, results should be interpreted carefully. In addition, all participants showed severe to very severe motor deficits at study onset, but showed a larger variability ranging from very severe (3 participants) to mild deficit (2 participants) after 6 weeks of rehabilitation, limiting the generalization of findings.
Conclusions
The combination of movement kinematics with brain activations of the corresponding movement in relation to stroke and clinical scores allowed us to demonstrate that movements of the ipsilesional upper limb (a) showed atypical kinematics and (b) were differently controlled. This seemed to result from a general disturbance of the bihemispheric (motor) network following stroke, rather than the stroke itself, as no link was found between these changes and stroke characteristics or clinical scores. First, the extended activation of the primary sensorimotor network is thought to reflect a despecialization of the nondamaged hemisphere early poststroke, requiring a well-known task to be relearned. Second, the additional recruitment of the middle temporal area and rolandic opercularis is thought to reflect adaptive control strategies in the absence of reliable proprioception: visualization to ensure task execution that is controlled in a feedback-based manner.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by the French “Ministère de la Santé” (MARGAUT: 2010-A00596-33) and the ‘Agence Nationale de la Recherche’ (NUMEV: ANR-10-LABX-20).