
Abstract
Walking rehabilitation is one of the primary goals in stroke survivors because of its great potential for recovery and its functional relevance in daily living activities. Although 70% to 80% of people in the chronic poststroke phases are able to walk, impairment of gait often persists, involving speed, endurance, and stability. Walking involves several brain regions, such as the sensorimotor cortex, supplementary motor area, cerebellum, and brainstem, which are approachable by the application of noninvasive brain stimulation (NIBS). NIBS techniques, such as repetitive transcranial magnetic stimulation and transcranial direct current stimulation, have been reported to modulate neural activity beyond the period of stimulation, facilitating neuroplasticity. NIBS methods have been largely applied for improving paretic hand motor function and stroke-associated cognitive deficits. Recent studies suggest a possible effectiveness of these techniques also in the recovery of poststroke gait disturbance. This article is a selective review about functional investigations addressing the mechanisms of lower-limb motor system reorganization after stroke and the application of NIBS for neurorehabilitation.
Introduction
Stroke is the leading cause of severe adult disability worldwide. 1 Several neurological functions can be affected by stroke, but the most common is the motor system. Less than 50% of stroke survivors recover completely despite intensive acute care and rehabilitation training. Considering gait impairment, after stroke, about 57% to 63% of individuals cannot walk independently at symptom onset and 22% to 50% after rehabilitation.2-8 It is well known that the human brain continues to adjust throughout life. Neuroplasticity is particularly active in the first months after a brain lesion, 9 but even years after stroke, the capacity to reorganize in response to interventions influencing motor recovery is still retained. 10 Several efforts have focused on the development of new restorative therapies. In particular, noninvasive brain stimulation (NIBS) techniques, such as transcranial magnetic stimulation (TMS) and transcranial direct current stimulation (tDCS), have been recognized as a promising intervention for treating poststroke motor deficits. This review summarizes the literature on the physiological control of normal walking, the reorganization of the motor system after stroke, and the application of NIBS to improve lower-limb motor function and walking in stroke patients.
Neural Control of Lower-Limb Motor Function and Normal Walking
Walking results from a dynamic interaction between central program and feedback mechanisms, occurring at spinal and supraspinal levels. The sensorimotor cortex, cerebellum, basal ganglia, brainstem, and the central pattern generator (CPG) are, indeed, all neural structures involved in gait.11-14 The CPG defines neuronal networks at the spinal cord level that generate intrinsic patterns of rhythmic movements in mammals.12,13,15 In humans, the actual role of CPG in generating gait is still debated. Converging works underlined the importance of peripheral sensory information 16 and motor cortical descending inputs in shaping CPG function. 17 The evolution of the corticospinal tract (CST) in mammals is associated with the acquisition of motor skills. Several studies show that people affected by stroke could walk even after extensive damage of the lateral CST,18-20 although the upper-limb motor function remains severely impaired.20,21 However, CST is considered essential for skilled gait by modulating the walking pattern in response to environmental influences.19-22 Fine control of walking involves several brain regions, including the sensorimotor cortex, supplementary motor area (SMA), cerebellum, and brainstem. 23
Several studies suggest that upper-limb brain activity is mainly lateralized over the contralateral sensorimotor regions during unilateral motor task in healthy individuals, with differences pointing to a less-lateralized activation related to aging or complexity of the motor task.24-26 However, a transfer of findings from arm to leg seems unjustified. In particular, a bihemispheric control of foot movements in healthy individuals has been hypothesized. 27 Few studies have investigated brain activation during lower-extremity movements. A single-photon-emission computerized tomography (SPECT) study in healthy individuals showed that walking is associated with activation of the primary motor cortex (M1) bilaterally, the SMA, and somatosensory cortex (S1), compared with contralateral M1 focus during isolated arm movements. 28 Near infrared spectroscopy (NIRS) monitors regional relative changes of hemoglobin concentration to measure cortical activation, utilizing the tight coupling between neural activity and regional cerebral blood flow. 29 The technique may help in visualizing brain activation during gross movements like locomotion, with certain limitations concerning the low spatial resolution and the possible head motion artifacts. NIRS has been applied for comparing the distribution of activation during locomotion, isolated arm swing, and isolated foot dorsiflexion. Walking, as well as isolated foot flexion, was associated with activation of M1 lower-limb representation and SMA bilaterally, whereas isolated arm swing predominantly involves the lateral motor cortices of the contralateral hemisphere. 30 Consistently, functional magnetic resonance imaging (fMRI) studies have compared the pattern of brain activation associated with isolated upper- and lower-limb joint movements. Finger or elbow movements show a stronger lateralized pattern of activation in comparison with lower-limb joints, implying a different functional specialization.22,31 Evaluating the differences between movement of different joints within the lower limb, a proximal to distal increase in lateralization was observed. 31 Moreover, the activation of SMA proper was one of the main features characterizing lower- with respect to upper-limb movement.22,31 Increased activity in the cerebellum and precuneus associated with lower-limb movement has been observed with aging. Recruitment of these areas has been interpreted as potentially compensatory for other age-related changes in the sensorimotor networks responsible for control of limb function. 32
Also, TMS techniques have been used to study lower-limb motor function and gait mechanisms as well. Most studies aimed at assessing the contribution of M1 and the CST in the control of lower-limb muscles (ie, distal) during human gait. The size of motor-evoked potentials (MEPs) in ankle dorsiflexor muscles to TMS over M1 is increased in amplitude only during phases of the gait cycle where a particular muscle is active. For example, MEPs in the soleus are present during stance and absent during swing.33-35 This observation suggests that dorsiflexion of the ankle in walking requires the participation of the brain. However, changes in the size of the TMS-induced MEPs could reflect modifications at cortical and subcortical levels as well. TMS of the motor cortex activates, indeed, cells not only with monosynaptic connections to the motoneurons but also pathways with polysynaptic connections.36,37 Petersen et al 38 tried to evaluate the effective contribution of the motor cortex, determining whether the increased cortical excitability directly relates to the activation of spinal motoneurons during human walking. Similar to what is observed for the upper limb, 39 the authors hypothesized that for the leg muscles also, the activation of inhibitory mechanisms within the cortex (ie, intracortical inhibition) should reduce cortical excitability and, consequently, the cortical motor output. Therefore, they evaluated the suppression of electromyography (EMG) activity in tibialis anterior and soleus muscles by a subthreshold TMS stimulus (below threshold for a MEP) delivered over the leg motor cortex during walking. They found that low-intensity TMS effectively produces a suppression of ongoing EMG activity during walking and demonstrated that motoneuronal activity during walking can be suppressed by activation of intracortical inhibitory circuits. 38 Moreover, evaluating the corticospinal control of gait, TMS studies demonstrate that a task induces modulation of MEPs in leg muscles. MEP facilitation seems to be greater during precision stepping tasks and selective for lower-leg or upper-leg muscles.34,40 Unlike for the upper limb, the somatotopy of the legs is located deep, adjacent to the midline; therefore, for technical limitations, the transcallosal connections between M1 of the 2 legs cannot be studied by TMS. However, Goodwill et al 41 observed an increase in strength of the left leg following unilateral training of the right lower limb. The cross-transfer of strength between the 2 legs was associated with an increase in excitability to TMS of the untrained leg motor cortex, implying changes in functional interhemispheric connectivity between the 2 motor cortices.
Effects of Ischemic Injuries on M1 or the CST
In the case that M1 or the CST were affected by a stroke event, depending on the biological individual age and on the localization and size of the ischemic lesion, the clinical outcome may vary from complete inability (plegia) to impairment (paresis) of motor performance. In comparison with hand function, walking function has a greater possibility of recovery because it is less dependent on the CST than hand function.19,20,42 In addition, walking requires a smaller degree of motor function. A person affected by stroke can walk when motor function is recovered to the proximal joint (hip and knee) at least to the degree of being against gravity; in contrast, hand function requires a larger degree of recovery to distal joints. 43 Despite that, the majority of studies on this topic have reported that 70% to 80% of stroke survivors are able to walk at the chronic stage2-6 even if walking speed and endurance often persist impaired. 44 Indeed, as previously mentioned, corticospinal connectivity plays a crucial role in lower-limb fine motor control. In particular, it is well known that corticospinal projections to motor neurons of the tibialis anterior are more pronounced than those of any other leg muscles. 33 In agreement, impaired gait performance after stroke is significantly associated with decreased dorsiflexor strength of the affected side. Deficits of gait coordination might persist in the chronic stages, indicating an impaired interaction among different functional connected networks involved in walking. The coordination deficits include the following: altered temporal and spatial coordination between the head, trunk, and pelvis45,46; impaired pelvic, knee, and ankle control during stance; and impaired excursions and timing during the swing phase. 47 Poor interlimb coordination is reflected by asymmetries in propulsive forces between the paretic and nonparetic limbs, step lengths and widths, and stance and swing phase durations. 48
Lower-Limb Motor System Reorganization After Stroke
At least part of the recovery process undoubtedly involves resolution of pathophysiological events associated with cortical injury; thus, mechanisms that mediate early improvements may include dynamic changes within the lesion and at the interface with the surrounding tissue, such as resolution of edema, vascular recanalization, and cellular remodeling. However, motor recovery after cortical injury occurs in large part through behavioral compensation, rather than true recovery or restitution of “normal” motor strategies. It has also been suggested that other cortical structures, either adjacent to or remote from the damaged area, can take over the function of the damaged area. This theory is known as vicariation of function. 49 Most of the recovery of function after stroke may represent actual relearning of motor skills with the injured brain; thus, the areas important in poststroke recovery are essentially those involved in healthy brain motor skill acquisition. These regions include M1, the premotor cortex, and the SMA. Finally, callosal fibers connect both M1 cortices partly directly but largely via the premotor cortex. 50 Therefore, structures that may help restore motor activity can be subsumed into 3 groups: intact perilesional areas of the ipsilesional M1; the ipsilesional and contralesional ancillary motor systems, next to structures responsible for executive control; and the contralesional motor system. 50 However, plastic processes of recovery have been mainly collected for the upper limb.51-54
The pathophysiological reorganization of leg motor outputs after stroke is still unclear. A study performed with NIRS technique in poststroke individuals showed that during walking, similarly to the upper limb, the cortical activation patterns of the motor, premotor, and supplementary lower-limb motor cortex are greater for the unaffected hemisphere than for the affected hemisphere. 55 Improvements of gait parameters of the paretic lower limb seem to be associated with a reduction in the interhemispheric asymmetry of the primary sensorimotor cortical activations in favor of the affected hemisphere. 56 The latter data suggest that the concept of interhemispheric competition, proposed for homologues of upper-limb motor areas, might be applied even in the case of lower-limb poststroke recovery. As described by Ward and Cohen 57 and more recently stated by Nowak et al, 58 the hypothesis of interhemispheric competition proposes that poststroke motor deficits are a result of reduced output from the damaged hemisphere, not only because of the infarct itself but also because of excessive transcallosal inhibition from the intact hemisphere. However, considering the differences in use patterns of upper (mainly unilateral, eg, reaching) and lower extremities (mainly bilateral, eg, walking) and the differences in brain activation associated with healthy arm and leg movement, 22 a transfer of findings from arm to leg seems unjustified. Moreover, as previously reported, the NIRS technique has the same limitations concerning the low spatial resolution and possible head motion artifacts. Enzinger et al, 59 observed in 2008, in a group of chronic subcortical stroke patients during ankle movements, an extent of fMRI activation comparable with that in controls in the SMAs and sensorimotor cortex of the unlesioned hemisphere associated with worse function of the paretic leg. The same group investigated the functional reorganization after 4 weeks of treadmill training in chronic poststroke individuals. Greater walking endurance after training correlated with increased brain activity in the primary sensorimotor cortex bilaterally, the cingulate motor areas and the caudate nuclei bilaterally, and in the thalamus of the affected hemisphere. 60 An fMRI study suggests that the neural control mechanism of walking might vary according to lesion localization at the cortical, subcortical, and brainstem levels, respectively. Similar activation patterns were observed in those with subcortical lesions and controls with recruitment of the contralateral primary motor cortex, SMA, and bilateral somatosensory area. Cortical recruitment was reduced in brainstem and cortical stroke. Better walking was associated with lesser contralateral sensorimotor cortex activation in the brainstem but stronger recruitment of ipsilateral sensorimotor and bilateral somatosensory cortices in patients with subcortical and cortical stroke, respectively. 61 In a combined TMS and MRI study, Jayaram et al 62 evaluated whether a variation in walking impairment in stroke survivors was associated with a variation in the structural or functional integrity of the CST. They used TMS to directly assess functional connectivity from the motor cortex of each hemisphere to both lower limbs (vastus lateralis bilaterally) in chronic stroke patients with persistent paresis of their lower limb. EMG measures were used to calculate both ipsilateral and contralateral recruitment curves for each lower limb. The slope of these recruitment curves was used to examine the strength of functional connectivity from the motor cortex in each hemisphere to the lower limbs in chronic stroke patients and to calculate a ratio between ipsilateral and contralateral outputs, referred to as the functional connectivity ratio. The structural integrity of the CST was determined using diffusion tensor MRI to measure the asymmetry in fractional anisotropy (FA) of the internal capsule. The functional connectivity ratio for the paretic leg correlated with walking impairment, such that greater relative ipsilateral connectivity was associated with slower walking speeds. Asymmetrical FA values, reflecting reduced structural integrity of the lesioned CST, were associated with greater walking impairment. Flexor carpi radialis (FCR) and FA asymmetry were strongly and positively correlated with each other. Therefore, individuals with relatively greater ipsilateral connectivity between the contralesional motor cortex and the paretic lower limb had more structural damage of the ipsilesional CST. 62
Taken all together, these data suggest that the reorganization of the contralesional motor system is potentially adaptive for lower-limb poststroke recovery, probably depending on lesion localization and the residual integrity of the ipsilesional CST.
Noninvasive Neuromodulation
Repetitive TMS (rTMS) and Theta Burst Stimulation (TBS)
rTMS refers to regularly repeated single TMS pulses delivered in trains at specific frequencies. In healthy individuals, low-frequency rTMS (≤1 Hz) usually results in cortical excitability reduction, 63 whereas with higher frequencies (>5 Hz) cortical excitability is usually increased. 64 Besides the well-known frequency-dependent modulation of cortical excitability, the rTMS aftereffect seems to depend on different factors, such as intensity of stimulation,65,66 number of pulses,67,68 or duration of stimulation. 69 TBS consists of brief bursts of theta frequency low-intensity stimuli. TBS delivered in a continuous pattern produces a decrease in motor cortex excitability, whereas an intermittent paradigm produces opposite effects. 70 The biological effect of both rTMS and TBS is also related to type and orientation of coil, besides distance between the coil and the brain. In particular, the geometric shape of the coil used for stimulation results in variations of focality and depth of magnetic field penetration. The figure-of-8 coil (butterfly coil) consists of 2 adjacent circular coils, placed side by side, each traversed by an opposite current. The induced tissue current is, at its maximum, directed under the central segment where the 2 windings meet, giving a more accurately defined area of stimulation. The main advantage of this coil shape is the focal stimulation of superficial cortical regions (about 2 cm from the scalp). The fundamental limitation of this tool is the dramatic drop of the intensity of the electric field deeper in the brain as a function of the distance. Therefore, to stimulate deep-brain regions such as the lower-limb motor cortex, a very high intensity would be needed. The double-cone coil was reported to have the ability to stimulate the leg motor area, which is 3 to 4 cm in depth. 71 This coil is made of 2 large, adjacent circular wings at an angle of 95° and induces a stronger, but less focal, electric field compared with a figure-of-8 coil. 72 The Hesed coil (H-coil) is a more recent device developed to effectively stimulate deep-brain regions without increasing the electric field intensity in the superficial cortical regions. 73 The basic concept is to obtain summation of the electric field in depth, minimizing the current components that only cause accumulation of surface charge. Roth et al 73 compared the ability of the H-coil and figure-8 coil to stimulate both the upper- and lower-limb motor cortical regions. They found lower resting and active motor thresholds, particularly for the abductor hallucis brevis but also for the abductor pollicis brevis with the H-coil, suggesting a more efficient activation of deeper motor cortical regions using the H-coil than the figure-of-8 coil. 74 The safety of H-coils has also been assessed at relatively high intensities (120% of the motor threshold). 75 The H-coils designed have been preferentially tested to evaluate their efficacy in cognitive and mood alterations in psychiatric and neurological diseases.76-78 Moreover, using a H-coil designed to target the lower-limb motor cortex, analgesic effects have been obtained in patients with painful diabetic neuropathy. 79
Others NIBS Techniques
The so-called tDCS consists of the application of weak electrical currents through the scalp. This technique utilizes 2 surface electrodes (anode and cathode), whose placement is fundamental for the outcome, by determining the direction of current flow. The montage with anode on the brain region of interest (anodal tDCS) has an excitatory effect (and cathodal tDCS the opposite), possibly through mechanisms involving neuronal membrane depolarization or hyperpolarization, respectively.80,81
Paired associative stimulation (PAS) consists of low-frequency repetitive peripheral nerve stimulation combined with timed TMS over the contralateral motor cortex. Depending on the interstimulus interval (ISI) between the peripheral nerve stimulation and the TMS pulse, PAS can increase or decrease the corticospinal excitability. To achieve PAS-induced facilitation of corticospinal pathways, the TMS must be delivered at an ISI equivalent to the estimated MEP latency plus 5 ms. To achieve PAS-induced inhibition, the TMS must be delivered with an ISI 10 ms shorter than the estimated MEP latency. 82
Neurorehabilitation and NIBS
NIBS techniques can induce behavioral effects by facilitating or inhibiting the cortical excitability depending on the stimulation protocol applied, as previously described. The rationale for the application of NIBS for the rehabilitation of neurological deficits is mainly related to 2 observations:
NIBS-induced brain plasticity: different NIBS protocols are able to change and modulate activity beyond the stimulation period. The mechanisms underlying the aftereffect of NIBS are not completely understood, and it is likely that a number of interacting factors can act together. One hypothesis is that the duration of the effect implicates changes in synaptic plasticity throughout mechanisms of long-term potentiation and depression.
83
However, experiments performed in healthy rat brains show that NIBS has the ability to mediate neural plasticity by enhancing the expressions of neurotransmitters and neurotrophins, such as glutamate, N-methyl-
The influence on activity of brain regions distant from the site of stimulation 92 : NIBS can modulate regions functionally connected to the target throughout corticocortical pathways. The facilitation or inhibition of one motor cortex can, indeed, produce opposite effects on the contralateral hemisphere,93,94 possibly by modulating the excitability of interhemispheric inhibitory circuits. 95 It has also been observed that the modulation of the premotor cortex or SMA can affect the excitability of the primary motor cortex.96,97 Moreover, NIBS also produces adaptations at the spinal level throughout corticospinal pathways. For instance, inhibitory rTMS (1 Hz) over the left M1 representation of the FCR muscle significantly increases the amplitude of the FCR H-reflex, which is an electrophysiological measure of the stretch reflex. 98 Low-frequency rTMS might be able to facilitate the monosynaptic spinal cord reflex by inhibiting the corticospinal projections that modulate segmental spinal excitability. In contrast, the facilitation of the motor cortex by enhancing CST excitability reduces the amplitude of the H-reflex. 99 Both 5-Hz rTMS and intermittent TBS over the leg motor area seem to induce a long-lasting depression of the stretch reflex when applied during the course of several days in individuals affected by multiple sclerosis.100,101 This NIBS application may be of great interest in rehabilitation for spasticity.
On the other hand, the application of NIBS in neurorehabilitation is limited by the intraindividual and interindividual variability of response. Studies performed on a large number of participants indicate that the same stimulation protocol may induce opposite after-effects (inhibition/facilitation) or produce no effects at all. It has also been observed that variation in response to NIBS is influenced by the recruitment of different interneuronal networks, particularly with the efficiency of late I-wave recruitment.102-104 Because of the cross-sectional experimental design of these studies, it is not possible to distinguish whether the variability observed is mainly a result of variation between individuals or session-to-session variation within each person. 102 Some of the possible involved factors are, indeed, intrinsic—that is, gender, age, and genetic (BDNF polymorphism Val66Met), whereas others such as time of the day of the stimulation session, concurrent aerobic exercise, pharmacological intake, and attention can be manipulated. 105 Many of the factors involved are, however, still unknown. Moreover, the amount and distribution of current reaching different brain regions varies as a function of individual physiology and anatomy. Skull thickness, distribution of cerebrospinal fluid, subcutaneous fat, and the topography of the cortical surface (ie, the pattern of sulci and gyri in the stimulated area) have a radical effect on current flow and add to the uncertainty about the destination and strength of transcranially delivered stimulation. 106 The issue becomes trickier if we consider that the stroke-related functional and anatomical changes of the motor system may increase the variability in response to NIBS. More specifically, current flow in patients might differ. For instance, the extent of the brain area affected by stroke, lesion localization at the cortical or subcortical level, integrity of the stimulation target, the stroke stage (acute, subcute, or chronic), and the state of intracortical inhibitory and excitatory circuits are some of the factors influencing the response to NIBS.107-109 As proposed for the upper motor system, 110 for gait disorders also, modeling studies would be important before applying NIBS to predict the outcome and identify the best stimulation protocol. Because current flow and the mechanisms underlying the effects of each NIBS technique on motor cortical function are very likely to differ, 111 we can hypothesize that different patients may benefit from different NIBS techniques and different stimulation protocols. Certainly, individualized mathematical models for predicting current flow offer a way to estimate current distributions and optimize current delivery to target areas. 106 However, to date, the relative lack of understanding of the NIBS mechanism of action makes behavioral effects difficult to predict.
Data from several studies suggest, indeed, a complex influence of transcranial stimulation on excitation/inhibition. 112 For example, the distinction between anodal and cathodal tDCS on the basis of the neuronal membrane depolarization or hyperpolarization, respectively, produced constitutes a simplification. The neuron cell membrane seems to act as a resistor when superficial compartments are hyperpolarized; then, deep compartments in the same cell will be depolarized and vice versa. 113 Variations in both electric field direction and relative cell morphologies will, therefore, produce variations in polarization profiles. 114 Moreover, modeling and MRI studies indicate that anodal polarization may also affect excitability in inhibitory interneurons.115,116 The behavioral consequences of NIBS could, therefore, be strictly related to the emergent properties of the stimulated networks, also considering the fact that an increase in excitability does not necessarily mean an increase in processing efficacy and vice versa. 112
NIBS and Lower-Limb Motor Recovery After Stroke
Although the ability to walk is impaired in more than 80% of poststroke patients, most efforts have focused on the recovery of the paretic upper extremity than on the recovery of lower-limb function. This might be mainly a result of functional and anatomical limitations. Concerning the latter, lower-limb cortical motor areas are located close to the midline into the depth of the medial longitudinal fissure. These areas are less easily approached with NIBS techniques, especially when dealing with distal muscles. However, studies on healthy individuals have demonstrated that rTMS, tDCS, and PAS protocols are able to modulate the excitability of the leg motor cortex,82,99,117 and a behavioral effect of anodal tDCS on lower-limb motor performance, as transient enhancement of maximal leg force, has been observed. 118
Because the pathophysiological reorganization of leg motor areas after stroke is still unclear, based on the model of interhemispheric competition after stroke established for the upper extremity, NIBS techniques have been mainly applied for inhibiting the unlesioned hemisphere or facilitating the lesioned hemisphere.
Inhibition of the Unlesioned Hemisphere
Jayaram and Stinear 119 evaluated the effect of different NIBS protocols applied during walking on lower-limb motor cortex excitability. They tested the inhibitory PAS and the inhibitory 1-Hz rTMS applied to preferentially stimulate the unaffected lower-limb M1 as well as the excitatory anodic tDCS applied with electrodes offset to preferentially stimulate the affected M1. They found that all NIBS protocols were effective in modulating excitability of both lower-limb motor areas. The modulator effects consisted of the increase of MEP amplitude over the paretic limb and the decrease of MEP amplitude over the nonparetic limb, but behavioral effects were tested in this study. Wang et al 120 first evaluated, in chronic stroke patients, the clinical effect of task-oriented training associated with 1-Hz rTMS performed to inhibit the unaffected lower-limb motor cortex. The authors performed a sham controlled study, using a figure-of-8 coil tangentially positioned over the leg area at the optimal site for response from the rectus femoris muscle. They showed that 10 daily sessions of rTMS enhanced the effect of task-oriented training on walking performance and motor control ability, thereby leading to a more symmetric gait pattern. Moreover, rTMS improved the symmetry of interhemispheric corticomotor excitability. Recently, the effect of 1-Hz rTMS over the leg motor area of the unaffected hemisphere has been evaluated also in the subacute phase of stroke (either infarction or hemorrhage). Here, 15 days of real or sham stimulation was followed by 45 minutes of physical therapy. Both the real and sham groups demonstrated significant improvement in all the clinical measurements tested. The application of 1-Hz rTMS yielded the beneficial effect of physical therapy alone (sham group) on balance, mobility, and ADL independence but not on the paretic lower-limb motor function specifically assessed with the Fugl-Meyer scale 121 (Table 1).
Behavioral Effects of NIBS on Lower-Limb Motor Function.
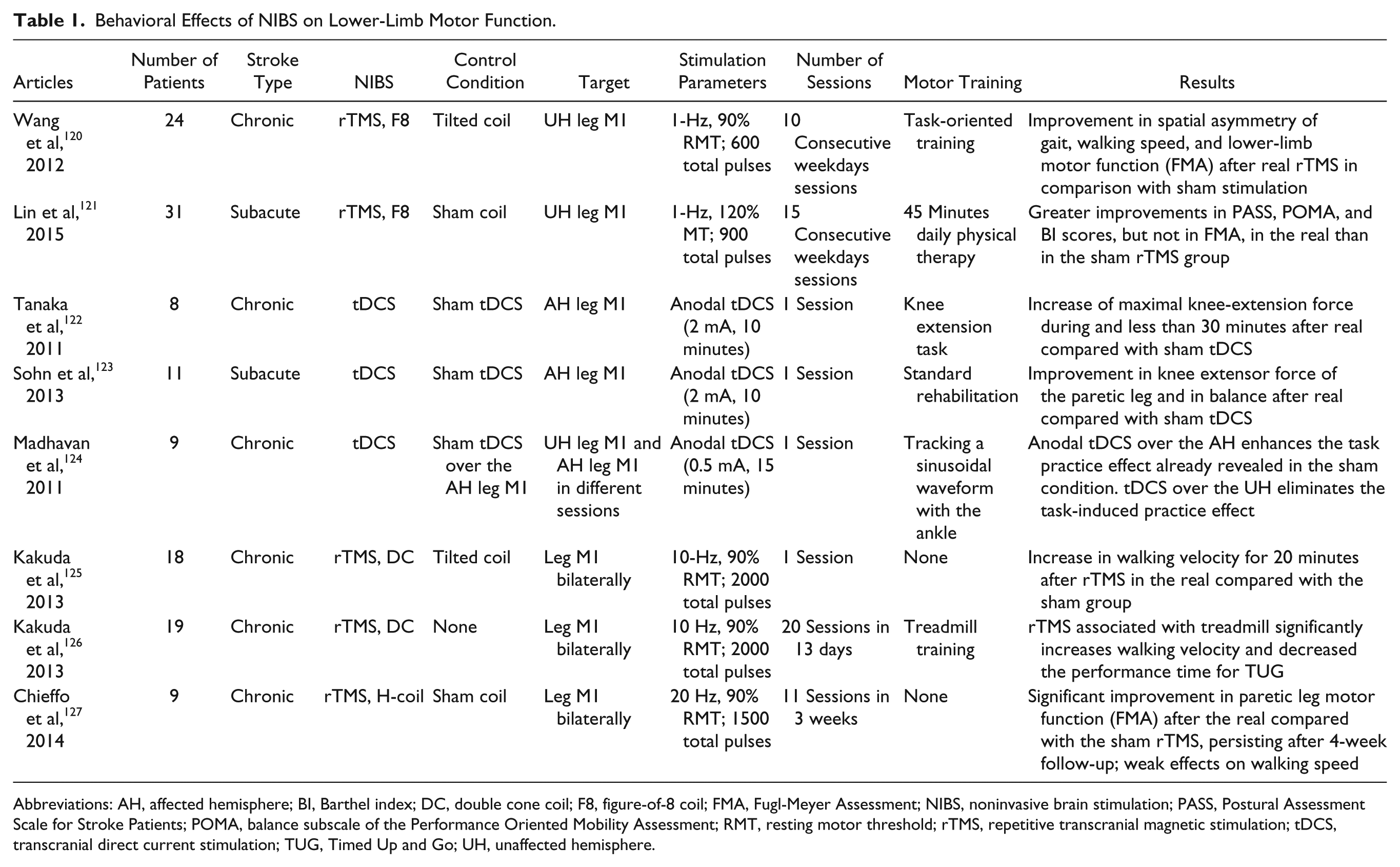
Abbreviations: AH, affected hemisphere; BI, Barthel index; DC, double cone coil; F8, figure-of-8 coil; FMA, Fugl-Meyer Assessment; NIBS, noninvasive brain stimulation; PASS, Postural Assessment Scale for Stroke Patients; POMA, balance subscale of the Performance Oriented Mobility Assessment; RMT, resting motor threshold; rTMS, repetitive transcranial magnetic stimulation; tDCS, transcranial direct current stimulation; TUG, Timed Up and Go; UH, unaffected hemisphere.
Facilitation of the Lesioned Hemisphere
A single session of anodal tDCS over the affected hemisphere transiently increases the knee extensor force in subacute and chronic stroke patients.122,123 Moreover, the effect of tDCS on fine motor control of the paretic foot has been evaluated by applying anodal or sham tDCS in random order over the lower limb M1 of the lesioned or nonlesioned hemisphere. In each session, tDCS was associated with a motor task that consisted of tracking a sinusoidal waveform for 15 minutes using dorsiflexion–plantar flexion movements of their paretic ankle. Anodal tDCS over the affected hemisphere enhanced the task practice effect already revealed in the sham stimulation condition. In contrast, tDCS applied over the unlesioned hemisphere eliminated the task-induced practice effect 124 (Table 1).
Bihemispheric facilitation
Hypothesizing a positive rather than detrimental role of the unaffected lower-extremity motor cortex in recovery mechanisms occurring after stroke, more recent studies have, instead, investigated the safety and efficacy of a high-frequency bilateral stimulation protocol targeting both the lower-limb motor cortices. Kakuda et al, 125 in a placebo-controlled cross-over study, found that a single session of high-frequency rTMS, over the leg motor area, bilaterally, using a double-cone coil, significantly improves walking performance for 20 minutes after stimulation in comparison with sham stimulation in a group of chronic poststroke patients (Table 1). The same authors in an open study (without placebo control) tested the effect of 20 sessions of high-frequency rTMS associated with treadmill training in poststroke hemiparetic patients with gait disturbance. They found that the combination of the 2 treatments significantly increased walking velocity and decreased the performance time for the Timed Up and Go Test 126 (Table 1). The effect of high-frequency (20 Hz) rTMS delivered with the H-coil over the leg motor cortices has been recently evaluated in a group of chronic subcortical stroke patients. They received both real and sham rTMS in a random sequence separated by a 4-week wash-out period; each treatment (real or sham) was composed of 11 sessions (administered over 3 weeks). In this study, rTMS treatment was not associated with specific motor training involving the lower limbs. The authors found that real rTMS treatment was associated with a significant improvement in leg motor function evaluated with the Fugl-Meyer scale. Such an effect persisted after 4 weeks of follow-up and was significantly greater than that observed with sham stimulation. A significant increase in walking speed (10 m walking test) was also found after real rTMS, but this effect did not reach statistical significance in comparison with sham stimulation 127 (Table 1).
Conclusions
Despite the deep localization in the central solcus, there is evidence that the cortical representation of leg muscles can be reached by NIBS techniques. In particular, rTMS coupled with double-cone or H-coils is able to produce a deep and wide electric field inducing an extensive and bilateral stimulation of the lower-limb motor cortices. In contrast, rTMS with figure-of-8 or tDCS delivered with small size electrodes seems to be able to focally stimulate one motor cortex, but the electric field produced is more superficial. The application of these techniques in clinical practice is still limited because of lack of exhaustive data providing definitive conclusions about the effectiveness and the best NIBS intervention to improve poststroke gait impairment. Indeed, contrasting NIBS approaches have been tested, with all producing positive results. So far, the mechanisms of motor recovery after stroke have been synthesized in the interhemispheric competition and vicariation models. The main difference between the 2 models regards the role of the unaffected hemisphere in recovery. The vicariation model refers to the positive contribution of the unaffected hemisphere to functional recovery after contralateral stroke lesions. Converging functional studies in chronic stroke patients suggest that the reorganization of the contralesional motor system is potentially adaptive for lower-limb recovery. High-frequency rTMS with double-cone or H-coils have been applied in chronic stroke patients to facilitate both the leg motor cortices producing positive effects on paretic lower-limb motor function or gait performance. The bilateral stimulation protocol is of interest considering that a bihemispheric control of leg movements has been hypothesized in healthy individuals and that the relative contribution of the 2 hemispheres in motor recovery after stroke could be modulated by several factors such as age, lesion size, lesion localization, time postonset, and the residual integrity of the ipsilesional CST. The interhemispheric competition model leads, instead, to an opposite NIBS approach because it assumes a detrimental role of the unaffected hemisphere in functional recovery because of excessive transcallosal inhibition. The application of NIBS to disrupt the activity of the unaffected hemisphere would be, therefore, beneficial for stroke recovery by reducing the abnormal transcallosal inhibition to the affected hemisphere. It is necessary to specify that the studies on which this model is based refer to the upper-limb motor system and mainly involve patients in the acute/subacute phases after stroke. There are no exhaustive functional data indicating that the interhemispheric competition model can be also applied for the lower-limb motor reorganization, especially in the chronic phases after stroke. However, 1-Hz rTMS delivered with the figure-of-8 coil to inhibit the unaffected lower-limb motor cortex enhanced the effect of task-oriented training, improving walking performance and motor control ability in a sham controlled study. Concluding, NIBS is feasible and potentially effective for poststroke gait disturbance. Given the huge variability among stroke patients, different procedures could be effective, but further studies with larger sample sizes are needed to better understand how clinical and neurological factors (ie, degree of impairment, lesion localization, structural and functional reserve) could influence the response to NIBS techniques.
Footnotes
Acknowledgements
The authors are grateful to Dr Simon Tiberi for writing advice.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Letizia Leocani reports support from the joint Italian-Israeli laboratory on brain modulation in neuroimmune, neurodegenerative, and mental disorders (Italian Ministry of Foreign Affairs).